
The Effect of Short-Term Artificial Feed Domestication on the Expression of Oxidative-Stress-Related Genes and Antioxidant Capacity in the Liver and Gill Tissues of Mandarin Fish (Siniperca chuatsi)
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Holding and Experimental Design
2.3. Antioxidant Capacity Assays
2.4. Gene Expression Analysis
2.5. Statistical Analysis
3. Results
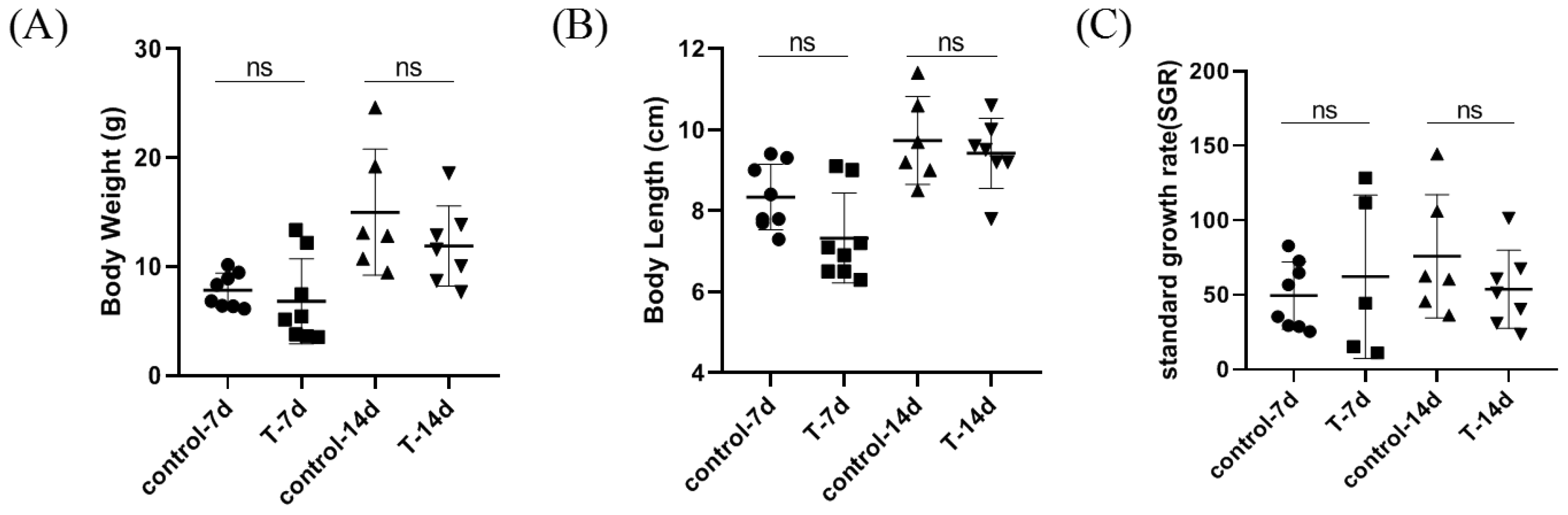
3.1. Artificial Feed Domestication Did Not Affect Weight and Length in S. chuatsi
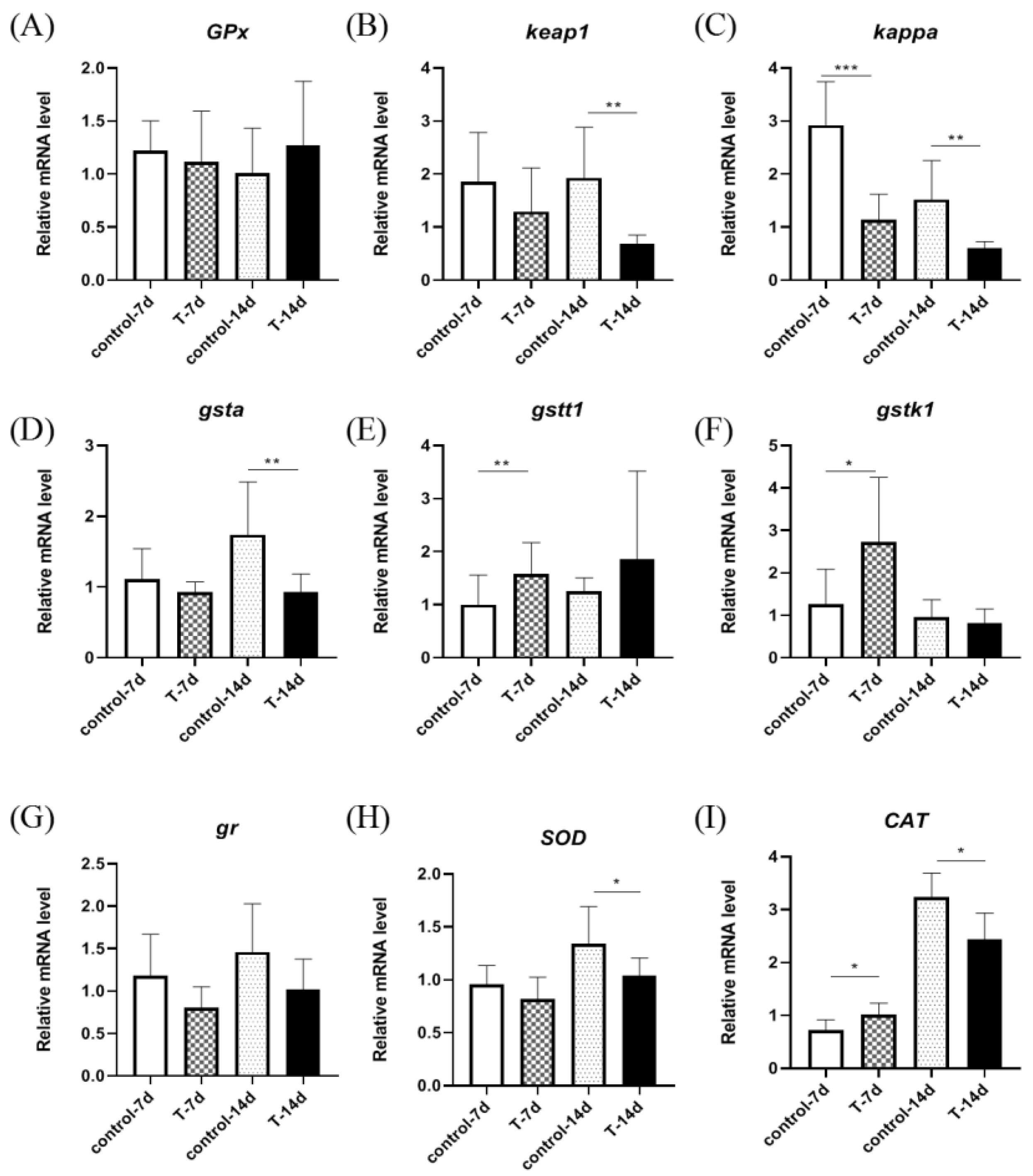
3.2. Artificial Feed Domestication Changed the mRNA Expression of Gill Antioxidant Genes in S. chuatsi
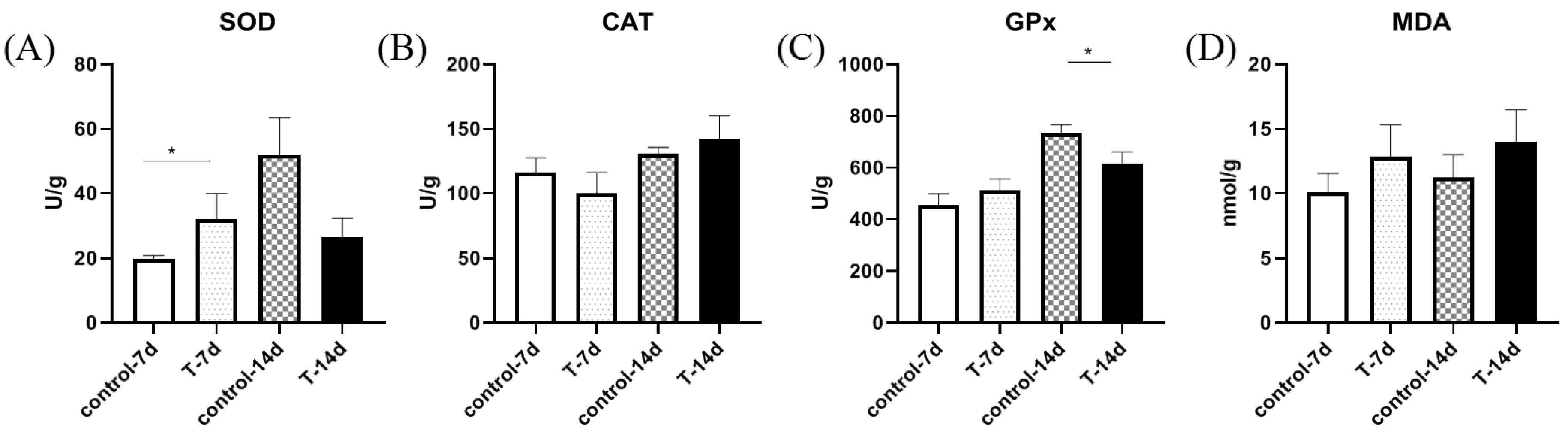
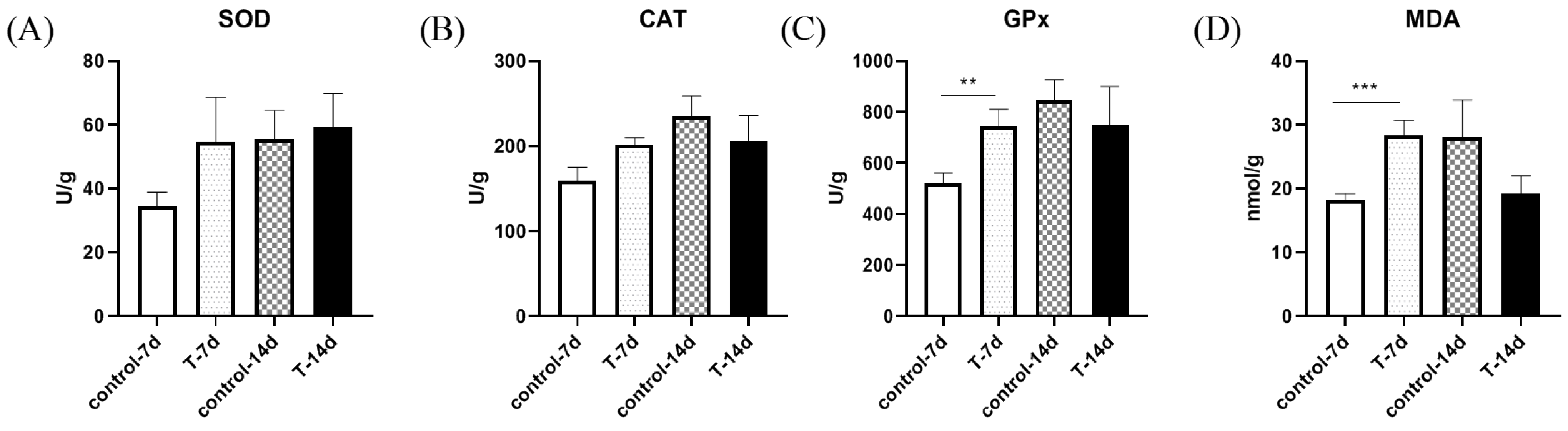
3.3. Artificial Feed Domestication Changed the Antioxidant Ability of Gills in S. chuatsi
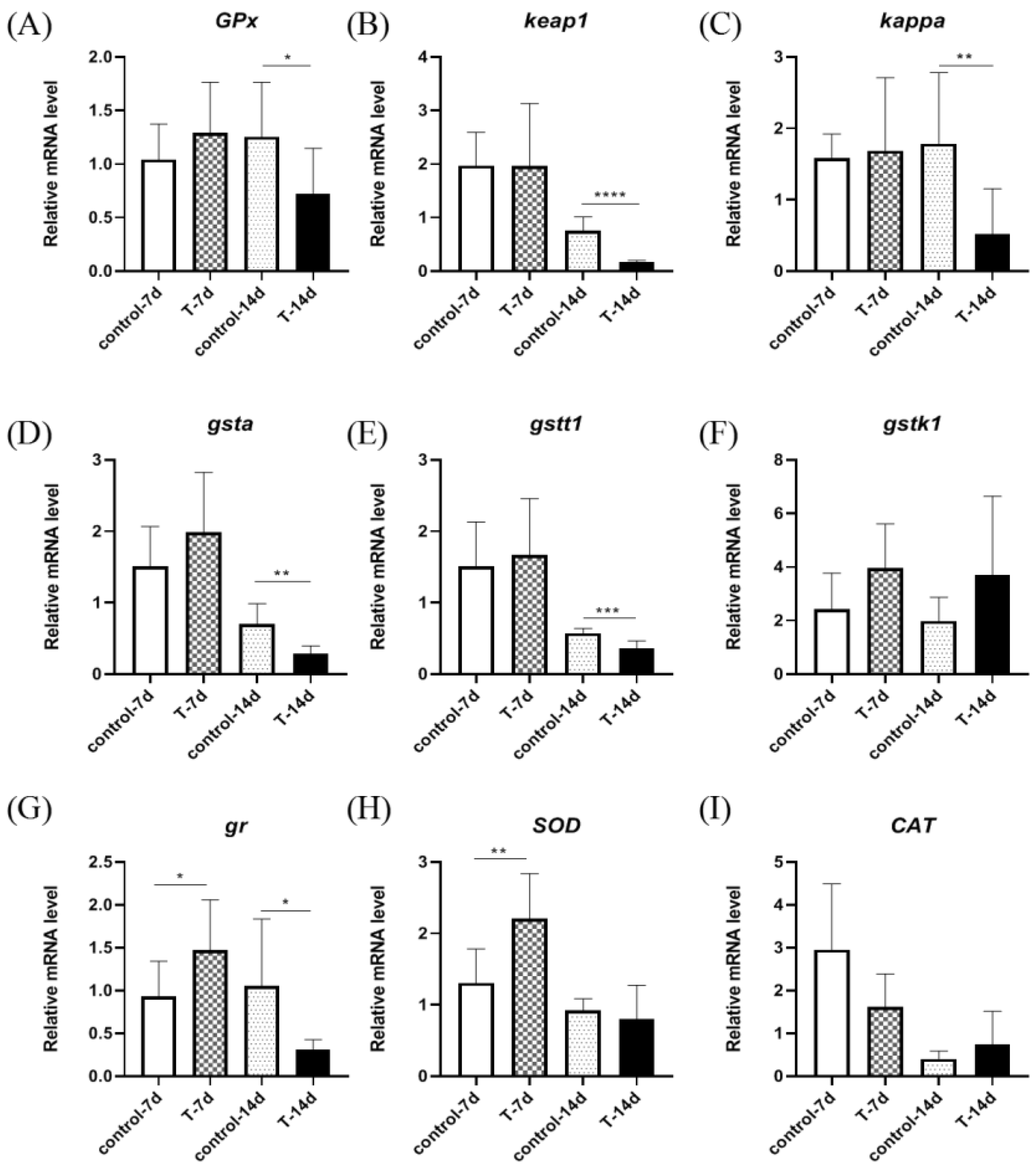
3.4. Artificial Feed Domestication Changed the mRNA Expression of Liver Antioxidant Genes in S. chuatsi
3.5. Artificial Feed Domestication Changed Liver Antioxidant Ability in S. chuatsi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gui, J.F.; Zhu, Z.Y. Molecular basis and genetic improvement of economically important traits in aquaculture animals. Sci. Bull. 2012, 57, 1751–1760. [Google Scholar] [CrossRef]
- Gui, J.F.; Zhou, L.; Li, X.Y. Rethinking fish biology and biotechnologies in the challenge era for burgeoning genome resources and strengthening food security. Water Biol. Secur. 2022, 1, 100002. [Google Scholar] [CrossRef]
- Sun, C.F.; Sun, H.L.; Dong, J.J.; Tian, Y.Y.; Hu, J.; Ye, X. Correlation analysis of mandarin fish (Siniperca chuatsi) growth hormone gene polymorphisms and growth traits. J. Genet. 2019, 98, 58. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chen, L.; Huang, R.; Gui, B.; Li, Y.; Li, Y.; Li, Y.; Liao, L.; Zhu, Z.; Wang, Y. Screening of Genes Related to Sex Determination and Differentiation in Mandarin Fish (Siniperca chuatsi). Int. J. Mol. Sci. 2022, 23, 7692. [Google Scholar] [CrossRef] [PubMed]
- LI, S.Z. A study on the geographical distribution of fish in the subfamily Sinipericinae. China J. Zool. 1991, 4, 40–44. [Google Scholar] [CrossRef]
- Wang, G.Y.; Zeng, K.W.; Gao, Y.A.; Li, Q.; Xia, R.L. The optimum dietary protein level for siniperca chuatsi. Acta Hydrobiol. Sin. 2005, 29, 189–192. [Google Scholar] [CrossRef]
- Liang, X.F. Principle and technology of Siniperca Chuatsi domesticated by artificial diet. Freshw. Fish. 1994, 24, 36–37. [Google Scholar]
- Wu, Z.L.; Li, P. Study on domestication and cultivation of Siniperca Chuatsi by artificial diet. Freshw. Fish. 1996, 26, 16–19. [Google Scholar]
- Liu, L.J.; Peng, K. Research progress on substitution of iced trash fish with aquatic compound feed. Feed. Res. 2021, 44, 141–144. [Google Scholar] [CrossRef]
- Wang, F.B.; Sun, C.F.; Dong, J.J. Effects of compound feed and live bait on nutrient composition and meat quality of Siniperca chuatsi. Freshw. Fish. 2022, 52, 102–111. [Google Scholar] [CrossRef]
- Niu, S.H.; Li, H.Y.; Pan, H.J. Effects of live prey fish and artificial diet on nutrient compositions and texture properties in the muscle of mandarin fish (Siniperca chuatsi). Acta Hydrobiol. Sin. 2023, 47, 37–44. [Google Scholar] [CrossRef]
- Chen, X.; Sun, C.F.; Dong, J.J. Comparative analysis of the gut microbiota of mandarin fish (Siniperca chuatsi) feeding on compound diets and live baits. Front. Genet. 2022, 13, 797420. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri, L.; Mackinnon, B.; St-Hilaire, S. Infectious diseases reported in warm-water marine fish cage culture in East and Southeast Asia—A systematic review. Aquac. Res. 2022, 53, 2081–2108. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H.Q. Targeting oxidative stress in disease: Promise and limitations of anti-oxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.X.; Cheng, C.; Xiao, M.; Zhu, M.J.; Li, L.J.; Mei, F.; Cha, Y.F.; Jian, S.J. Effects of stocking density on growth, antioxidant enzyme activity and related gene expression of juvenile Siberian hybrid sturgeon [Acipenserbaerii(♀) × A. schrenckii(♂)]. J. Fish. China 2022, 46, 1582–1592. [Google Scholar] [CrossRef]
- Wang, W.J.; Meng, Q.Q.; Li, M.J.; Yao, Z.B.; Wang, Y.J.; Wu, M.T.; Zhang, M.Q.; Wang, W.X. Effects of dietary levels on growth, serum biochemical indices, glycometabolism and antioxidant capacity of common carp. Jiangsu J. Agr. Sci. 2023, 39, 1908–1916. [Google Scholar]
- Fan, Y.W.; Zhang, M.Z.; Li, M.; Qian, Y.X.; Wang, R.X.; Jiang, H.B. Effects of dietary Haematococcus pluvialis on growth, antioxidant enzyme activity, immune response and ammonia tolerance in yellow catfish palteobagrus fulvidraco. J. Fish. China 2022, 46, 2168–2176. [Google Scholar] [CrossRef]
- Rudneva, I.I. Antioxidant system of Black Sea animals in early development. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1999, 122, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Knapen, M.F.C.M.; Zusterzeel, P.L.M.; Peters, W.H.M. Glutathione and glutathione-related enzymes in reproduction. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 82, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Alfafara, C.G.; Kanda, A.; Shioi, T. Effect of amino acids on glutathione production by Naccharomyces cereeiiae. Appl. Microbiol. Biotechnol. 1992, 36, 538–540. [Google Scholar] [CrossRef]
- Feng, T.; Zheng, W.Y.; Hong, W.S.; Yu, Q.; Zheng, S.L.; Zhang, Q. The effects of benzo(a)pyrene on antioxidant defenses in the liver of Boleophthalmus pectinirostris. Mar. Sci. 2000, 24, 27–30. [Google Scholar]
- Wu, P.; Chen, L.; Cheng, J.; Pan, Y.X.; Zhu, X.; Chu, W.Y.; Zhang, J.S. Effect of starvation and refeeding on reactive oxygen species, autophagy and oxidative stress in Chinese perch (Siniperca chuatsi) muscle growth. Fish Biol. 2022, 101, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Calo, J.; Blanco, A.M.; Comesaña, S. First evidence for the presence of amino acid sensing mechanisms in the fish gastrointestinal tract. Sci. Rep. 2021, 11, 4933. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.D.; Xu, J.; Song, Y.D. Effects of replacing live bait with compound feed on growth, digestion, and expression of small peptide transporter (PepT1) gene of Siniperca chuatsi. J. South. Agric. 2021, 52, 228–237. [Google Scholar] [CrossRef]
- Liu, W.; Hong, Y.C.; Sun, K.H.; Chai, Y. Research progress on feeding habits and domesticated feed combinations of Micropterus salmoides fry. Nutr. Feed 2022, 8, 68–69. [Google Scholar] [CrossRef]
- Shi, Y.; Zhong, L.; Fan, Y.; Zhang, J.; Zhong, H.; Liu, X.; Shao, C.; Hu, Y. The protective effect of mulberry leaf flavonoids on high-carbohydrate-induced liver oxidative stress, inflammatory response, and intestinal microbiota disturbance in Monopterusalbus. Antioxidants 2022, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.M.; Abdal Dayem, A.; Cho, S.G. Correlation between oxidative stress, nutrition, and cancer initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef] [PubMed]
- Pickering, A.D. Rainbow trout husbandry: Management of the stress response. Aquaculture 1992, 100, 125–139. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Yang, Y.N. Research progress of oxidative stress in diabetic keratopathy. Int. Eye Sci. 2022, 22, 399–402. [Google Scholar] [CrossRef]
- Li, H.; Xu, W.; Wu, L.; Dong, B.; Jin, J.; Han, D.; Zhu, X.; Yang, Y.; Liu, H.; Xie, S. Differential regulation of endoplasmic reticulum stress-induced autophagy and apoptosis in two strains of gibel carp (Carassius gibelio) exposed to acute waterborne cadmium. Aquat. Toxicol. 2021, 231, 105721. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Huang, Z.; Lin, H.; Ma, Z.; Wang, J.; Wang, Y.; Yu, W. Rhizoma curcumae Longae ameliorates high dietary carbohydrate-induced hepatic oxidative stress, inflammation in golden pompano Trachinotus ovatus. Fish Shellfish Immunol. 2022, 130, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Xie, S.; Zhuang, Z. Chlorogenic acid improves health in juvenile largemouth bass (Micropterus salmoides) fed high-fat diets: Involvement of lipid metabolism, antioxidant ability, inflammatory response, and intestinal integrity. Aquaculture 2021, 545, 737169. [Google Scholar] [CrossRef]
- Xu, L.; Ran, C.; He, S. Effects of dietary yeast nucleotides on growth, non-specific immunity, intestine growth and intestinal microbiota of juvenile hybrid tilapia Oreochromis niloticus♀ × Oreochromis aureus♂. Anim. Nutr. 2015, 1, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Reda, R.M.; Selim, K.M.; Mahmoud, R. Effect of dietary yeast nucleotide on antioxidant activity, non-specific immunity, intestinal cytokines, and disease resistance in Nile tilapia. Fish Shellfish Immunol. 2018, 80, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Jin, M.; Yuan, Y. Dietary nucleotide-rich yeast supplementation improves growth, innate immunity and intestinal morphology of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1425–1435. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free. Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef]
- Cong, S.M.; Wang, W.F.; Gao, C.R. Effects of compound diet and fresh frozen Ammodytes personatus on growth, antioxidant ability and lipid metabolism of hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂) juveniles. J. Fish. China 2016, 40, 1398–1407. [Google Scholar] [CrossRef]
- Zhou, C.; Liu, B.; Ge, X. Effect of dietary carbohydrate on the growth performance, immune response, hepatic antioxidant abilities and heat shock protein 70 expression of Wuchang bream, Megalobrama amblycephala. J. Appl. Ichthyol. 2013, 29, 1348–1356. [Google Scholar] [CrossRef]
- Garrett, T.; Rachel, A.; Ruoqiong, C. Glutathione as a marker for human disease. Adv. Clin. Chem. 2018, 87, 141–159. [Google Scholar] [CrossRef]
- Zhang, h.; Yan, X.; Yang, L.P.; Qin, C.B.; Pang, P.; Yang, B.W.; Nie, G.X. Effects of high-carbohydrate diet supplemented with sea buckthorn powder on the growth, antioxidant ability and non-specific immunity of Yellow River carp (Cyprinus carpio haematopterus). J. Fish. China 2023, 47, 109610. [Google Scholar] [CrossRef]
- Zhang, Y.R.; Gao, K.D.; Ren, Y.H. Broken Xinyang Maojian tea supplementation in a high -fat diet improves the growth performance, flesh quality and lipid metabolism of yellow river carp (Cyprinus carpio). Aquac. Rep. 2022, 25, 101236. [Google Scholar] [CrossRef]
- Dong, Y.Z.; Xia, T.; Lin, J.B. Quercetin attenuates high-fat diet induced excessive fat deposition of spotted seabass (Lateolabrax maculatus) through the regulatory for mitochondria and endoplasmic reticulum. Front. Mar. Sci. 2021, 8, 76811. [Google Scholar] [CrossRef]
- Wang, T.; Chen, J.L.; Jin, J.H.; Ming, J.H.; Shao, X.P.; Yang, X.; Ye, J.Y.; Wu, C.L. Effecte of Quercetin on growth performance, antioxidant capacity, lipid metabolism and intestina flora of Black Carp (Mylopharyngodon piceus) fed high-fat diets. Chin. J. Anim. Nutr. 2023, 35, 6598–6613. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, H.; Liang, G.Y. Effects of dietary baicalein on growth, serum anti-oxidation indicators and flesh quality of Ctenopharyngodon idella. J. Fish. China 2019, 43, 2383–23932. [Google Scholar] [CrossRef]
- Sun, R.Y.; Cheng, Y.F.; Zhang, B.Y. Effects of dietary quercetin on growth performance, digestive enzyme activities, immune and antioxidant functions of Cyprinus carpio. Chin. J. Anim. Nutr. 2023, 35, 2514–2523. [Google Scholar] [CrossRef]
- Zhang, C.N.; Jiang, D.X.; Wang, J.H. The effects of TPT and dietary quercetin on growth, hepatic oxidative damage and apoptosis in zebrafish. Ecotoxicol. Environ. Saf. 2021, 224, 112697. [Google Scholar] [CrossRef] [PubMed]
- Ghafarifarsani, H.; Hoseinifar, S.H.; Javahery, S. Effects of dietary vitamin C, thyme essential oil, and quercetin on the immunological and antioxidant status of common carp (Cyprinus carpio). Aquaculture 2022, 553, 738053. [Google Scholar] [CrossRef]
- Kong, Y.X.; Tian, J.X.; Niu, X.T. Effects of dietary quercetin on growth, antioxidant capacity, immune response and immune-related gene expression in snakehead fish, Channa argus. Aquac. Rep. 2022, 26, 101314. [Google Scholar] [CrossRef]
- Xu, Q.; Zhao, Q.R.; Li, R.M.; Ren, Y.B.; Wang, G.Q. Effects of Lycium barbarum Polysaccharides in Feed on Growth, Antioxidant Capacity and Immunity of Channa. J. Econ. Anim. 2024, 4, 1–7. Available online: http://link.cnki.net/urlid/22.1258.S.20240124.0921.002 (accessed on 2 April 2024).
- Zhang, Y.; Gong, Y.; Li, W.F.; Wang, Z.J.; Chen, N.S.; Li, S.L. Effects of Dietary Phospholipid Level on Growth Performance, Antioxidant Capacity and Fat Deposition of Juvenile Snakehead (Channa argus). J. Guangdong Ocean Univ. 2023, 6, 9–18. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | Function |
|---|---|---|
| CAT | F: CCCGATATGGTGTGGGACTT R: GAAGGTGTGAGAGCCGTAGC | Antioxidant-related |
| GPx | F: GCCCATCCCCTGTTTGTG R: AACTTCCTGCTGTAACGCTTG | Antioxidant-related |
| GR | F: CAGGCATCCTTTCCACCC R: TCCAGTCCTCTGTCCGTTTTA | Antioxidant-related |
| GSTA | F: TGGAGCACAAGTCACAGGAAG R: TGCTGCGTAGGATTCATTCA | Antioxidant-related |
| GSTK1 | F: AAGCCTCCTGGTCTGGTTCC R: ACCCGCTCCACCTGCTTG | Antioxidant-related |
| GSTT1 | F: CGAAGGCGAAGATGGACG R: GATTTTGTCGCCGATGATGAA | Antioxidant-related |
| KAPPA | F: GTGGCAACCCAGGAGGAG R: GGGAATGGCAACGGACA | Antioxidant-related |
| KEAP1 | F: TTCCACGCCCTCCTCAA R: TGTACCCTCCCGTATG | Antioxidant-related |
| SOD | F: ACAATCCCCACGGCAAGAAT R: TTGAGTAGGGGCCAGTGAGG | Antioxidant-related |
| Rpl13 | F: CACAAGAAGGAGAAGGCTCGGGT R: TTTGGCTCTCTTGGCACGGAT | Housekeeping gene |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Yuan, X.; Wu, H.; Gao, J.; Wu, J.; Xiong, Z.; Feng, Z.; Xie, M.; Li, S.; Xie, Z.; et al. The Effect of Short-Term Artificial Feed Domestication on the Expression of Oxidative-Stress-Related Genes and Antioxidant Capacity in the Liver and Gill Tissues of Mandarin Fish (Siniperca chuatsi). Genes 2024, 15, 487. https://doi.org/10.3390/genes15040487
Zhang Z, Yuan X, Wu H, Gao J, Wu J, Xiong Z, Feng Z, Xie M, Li S, Xie Z, et al. The Effect of Short-Term Artificial Feed Domestication on the Expression of Oxidative-Stress-Related Genes and Antioxidant Capacity in the Liver and Gill Tissues of Mandarin Fish (Siniperca chuatsi). Genes. 2024; 15(4):487. https://doi.org/10.3390/genes15040487
Chicago/Turabian StyleZhang, Zhou, Xiping Yuan, Hao Wu, Jinwei Gao, Jiayu Wu, Zhenzhen Xiong, Zhifeng Feng, Min Xie, Shaoming Li, Zhonggui Xie, and et al. 2024. "The Effect of Short-Term Artificial Feed Domestication on the Expression of Oxidative-Stress-Related Genes and Antioxidant Capacity in the Liver and Gill Tissues of Mandarin Fish (Siniperca chuatsi)" Genes 15, no. 4: 487. https://doi.org/10.3390/genes15040487