
Impact of Vanadium–Titanium–Magnetite Mining Activities on Endophytic Bacterial Communities and Functions in the Root Systems of Local Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Plant Roots and Extraction of Endophytic Bacterial DNA from the Roots
2.2. PCR Amplification and Detection
2.3. Library Preparation, Sequencing, and Raw Data Processing
2.4. OTU Cluster and Species Annotation
2.5. α and β Diversity Analyses
2.6. Functional Prediction of Endophytic Bacteria
2.7. Statistical Analysis
3. Results
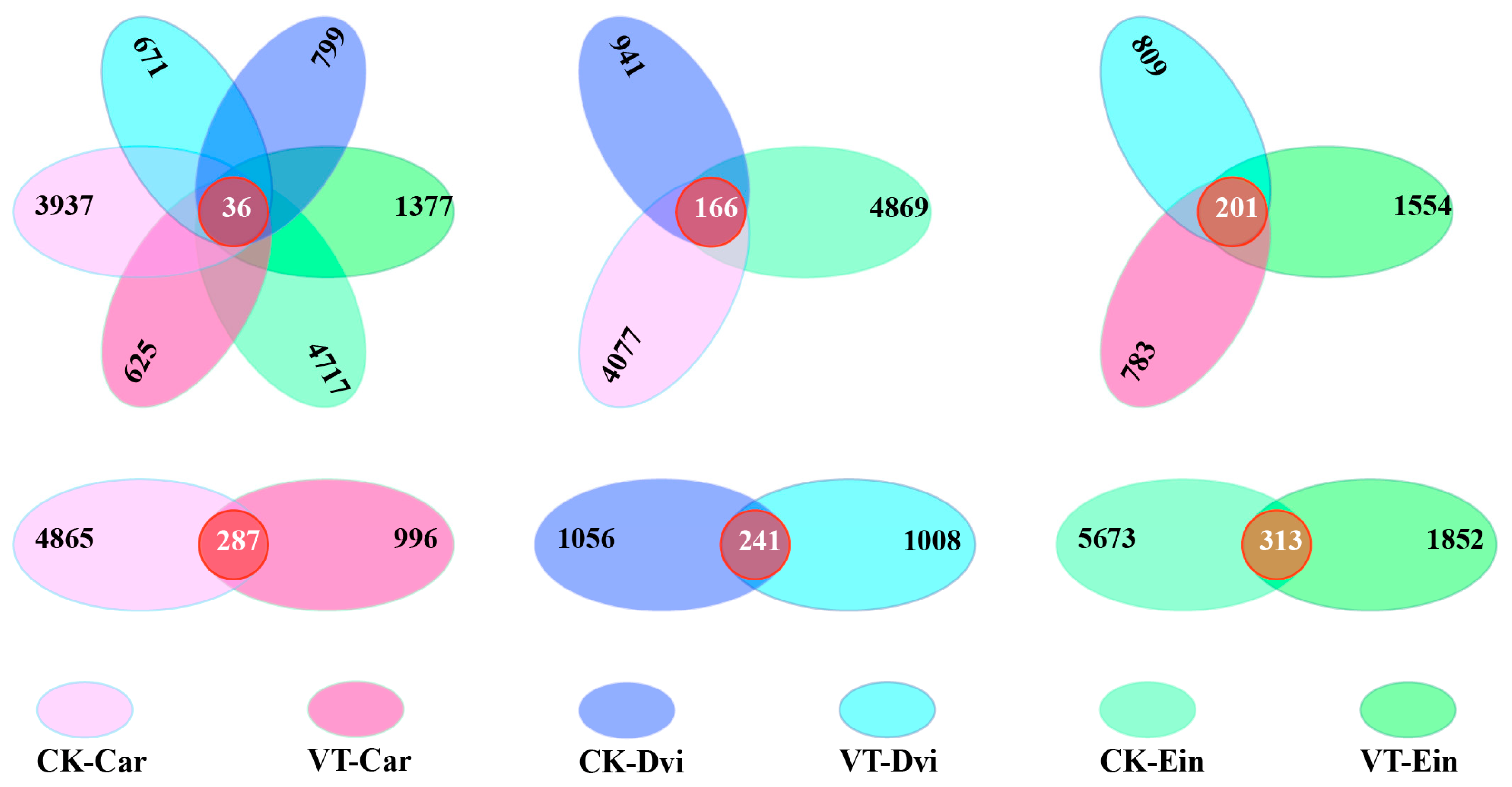
3.1. High-Throughput Sequencing Statistics of Endophytic Bacterial Communities
3.2. α Diversity Indices
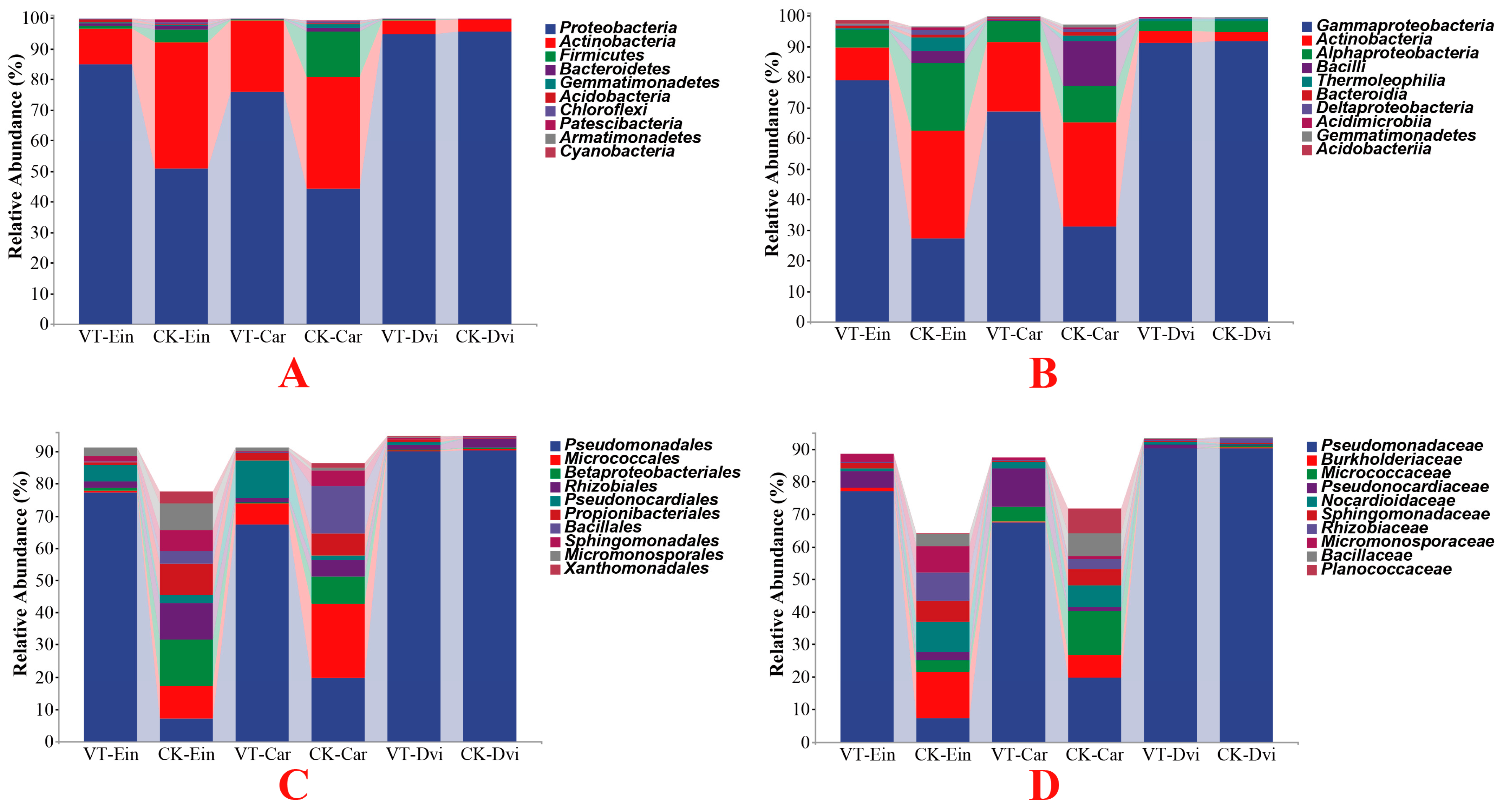
3.3. Taxonomic Analyses of the Endophytic Bacterial Communities
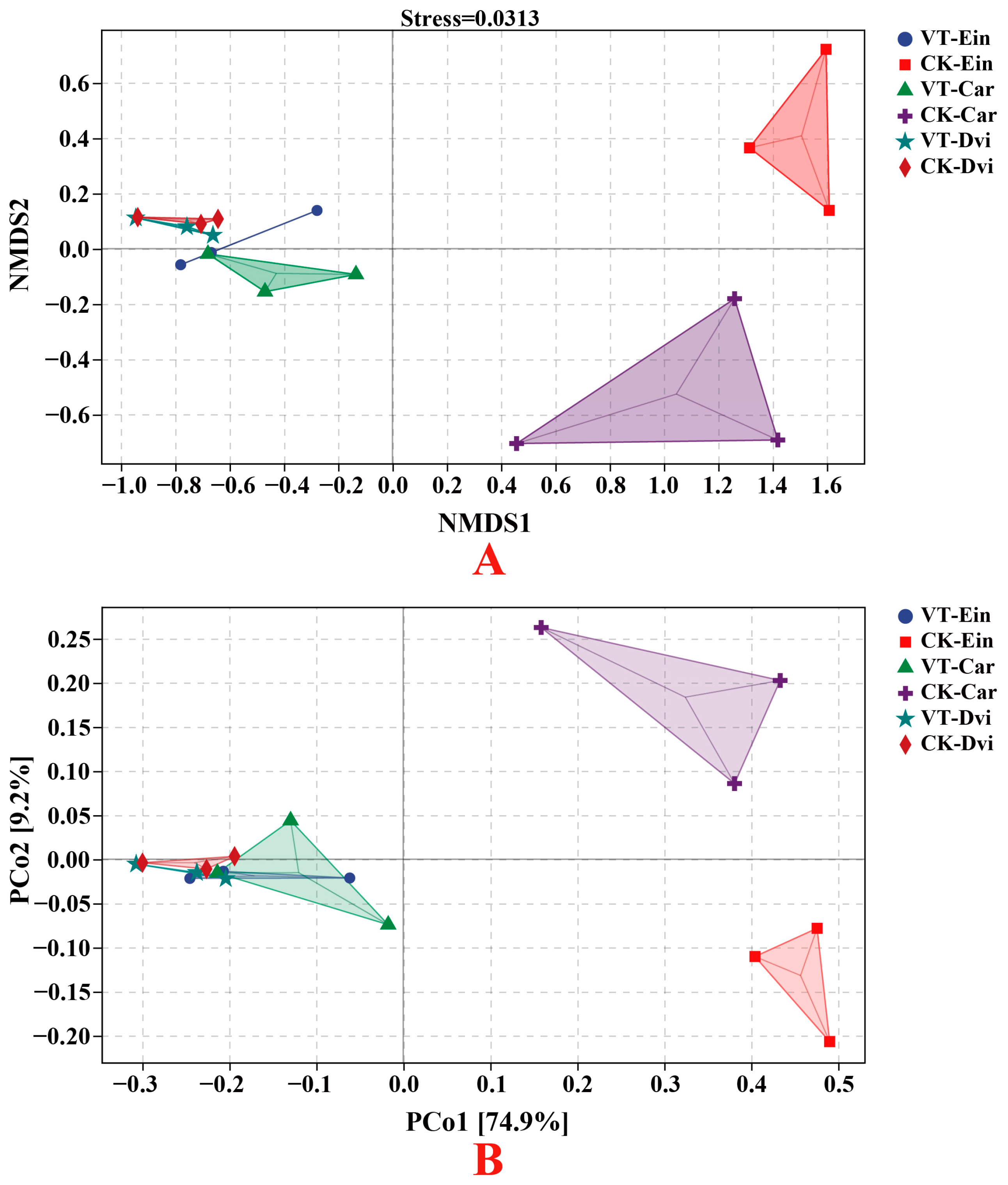
3.4. Structural Differentiation of Endophytic Bacterial Communities
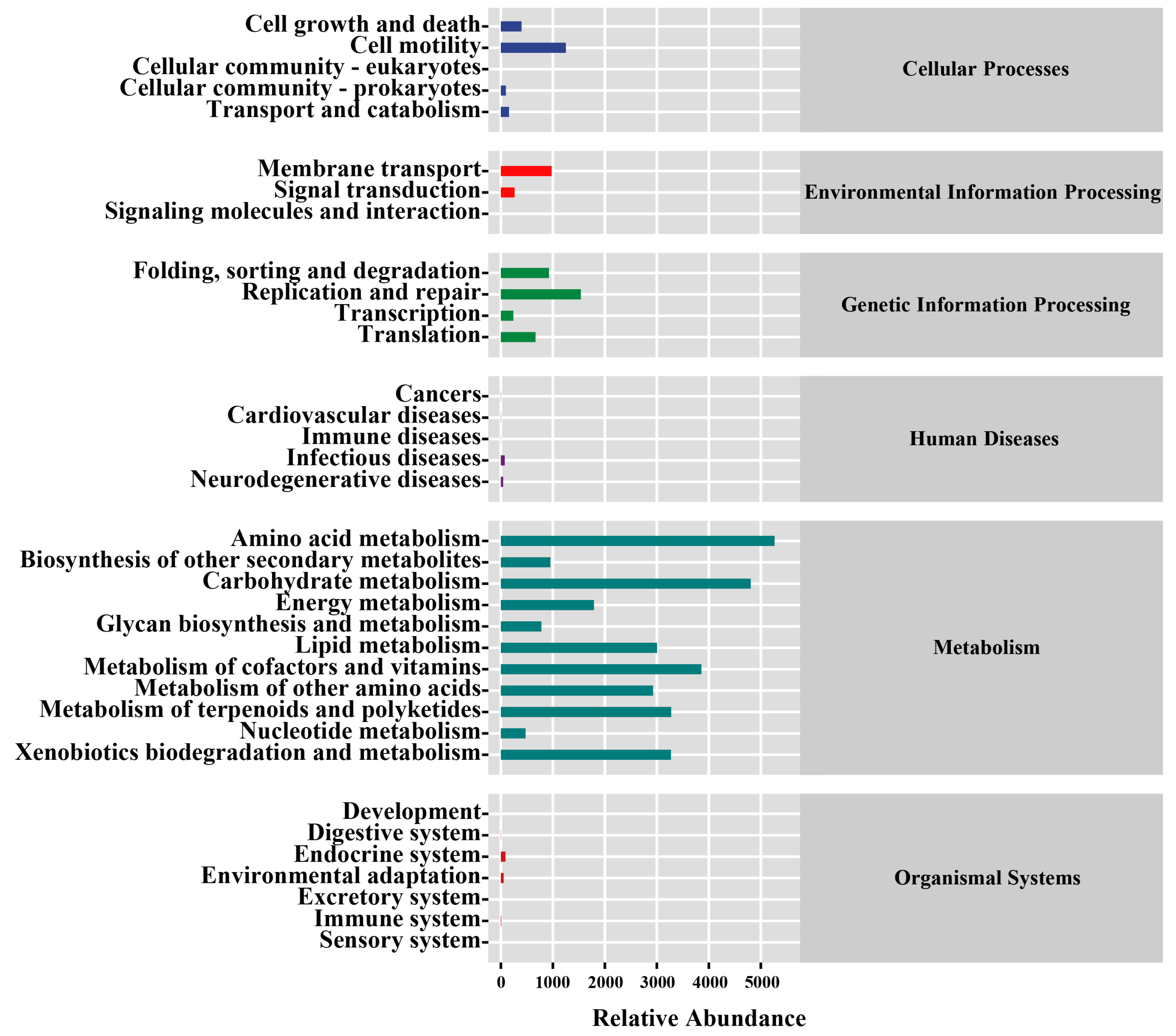
3.5. Function Prediction of the Endophytic Bacterial Communities
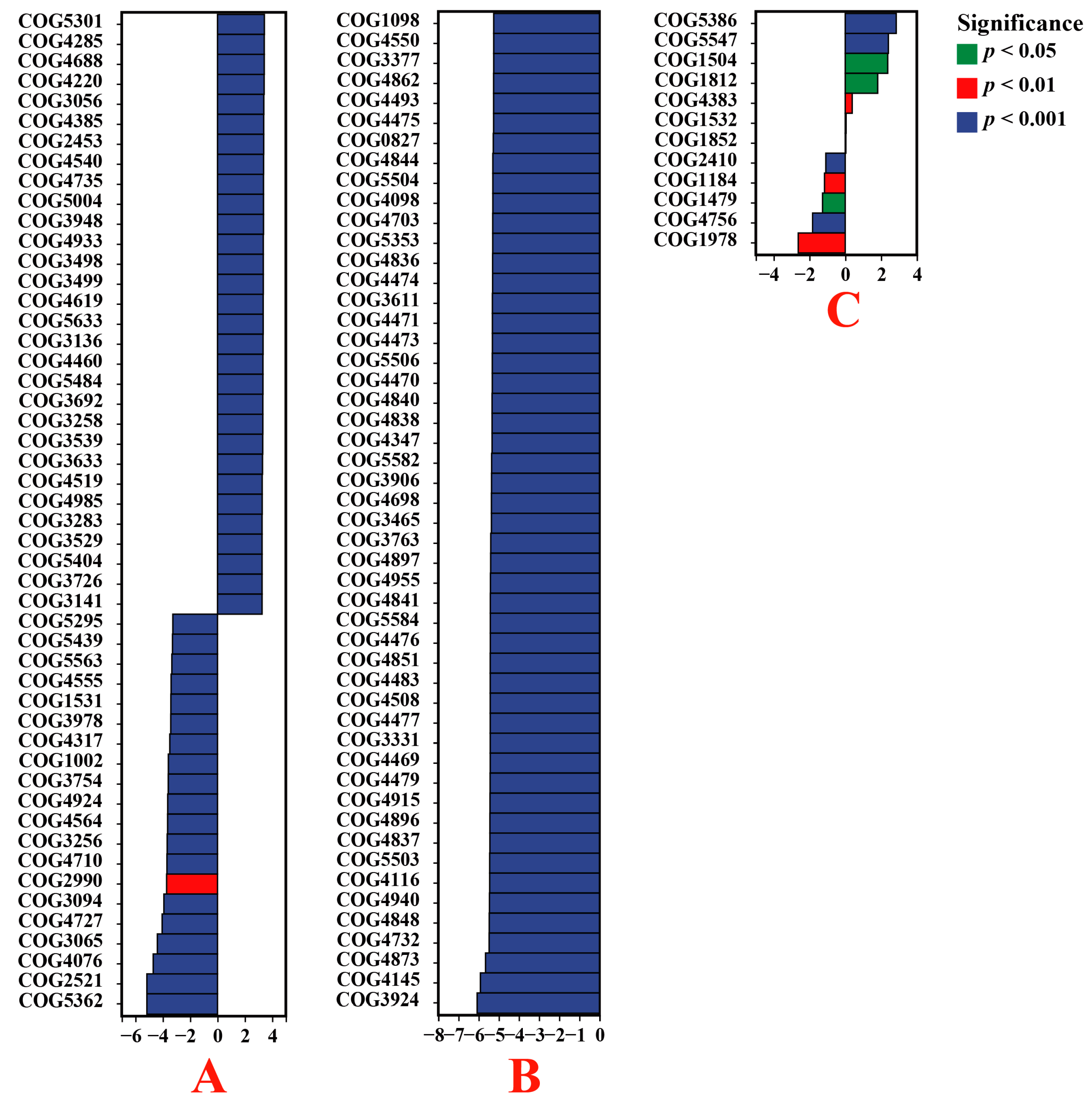
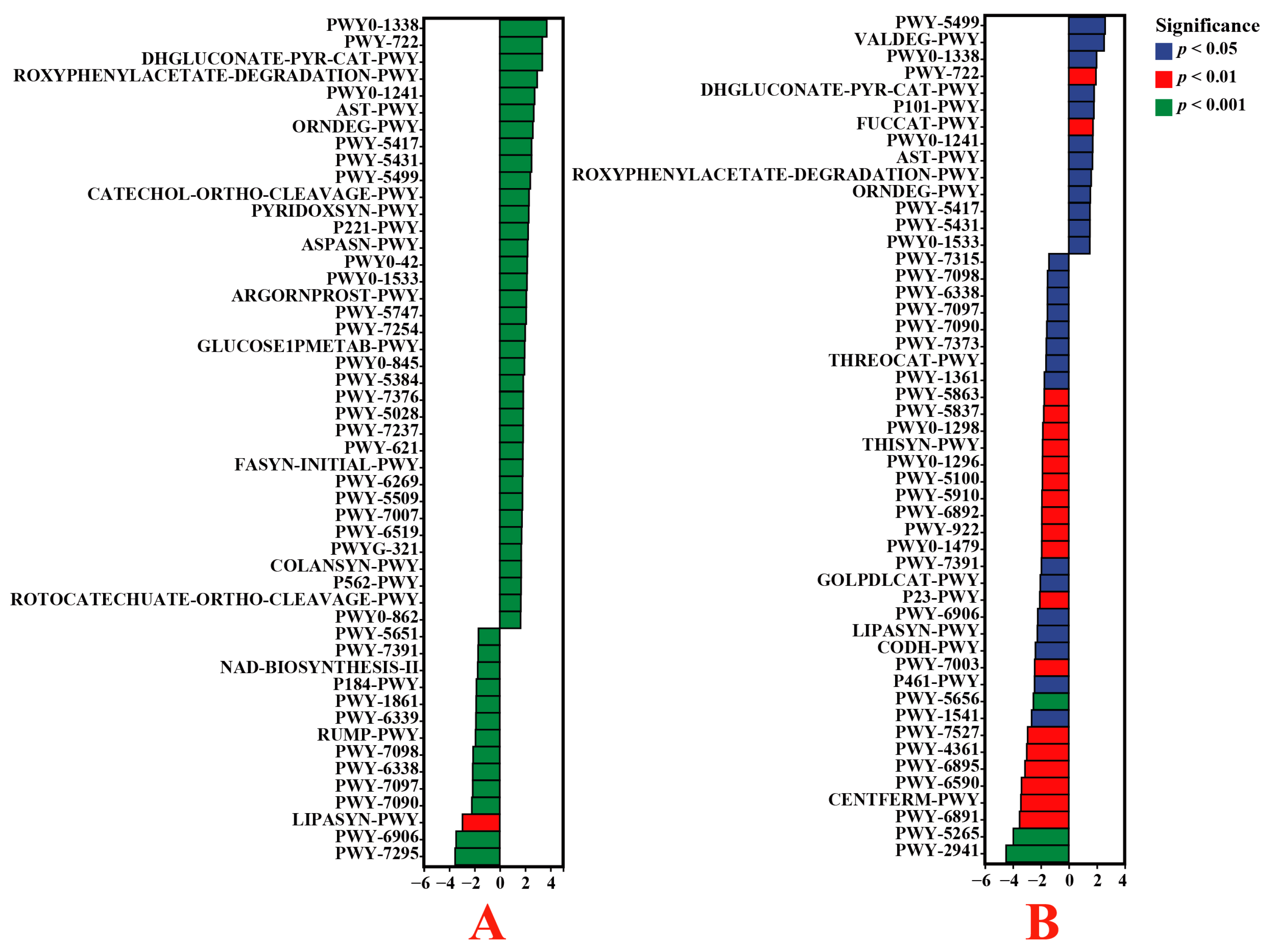
3.6. Functional Enrichment of Endophytic Bacteria
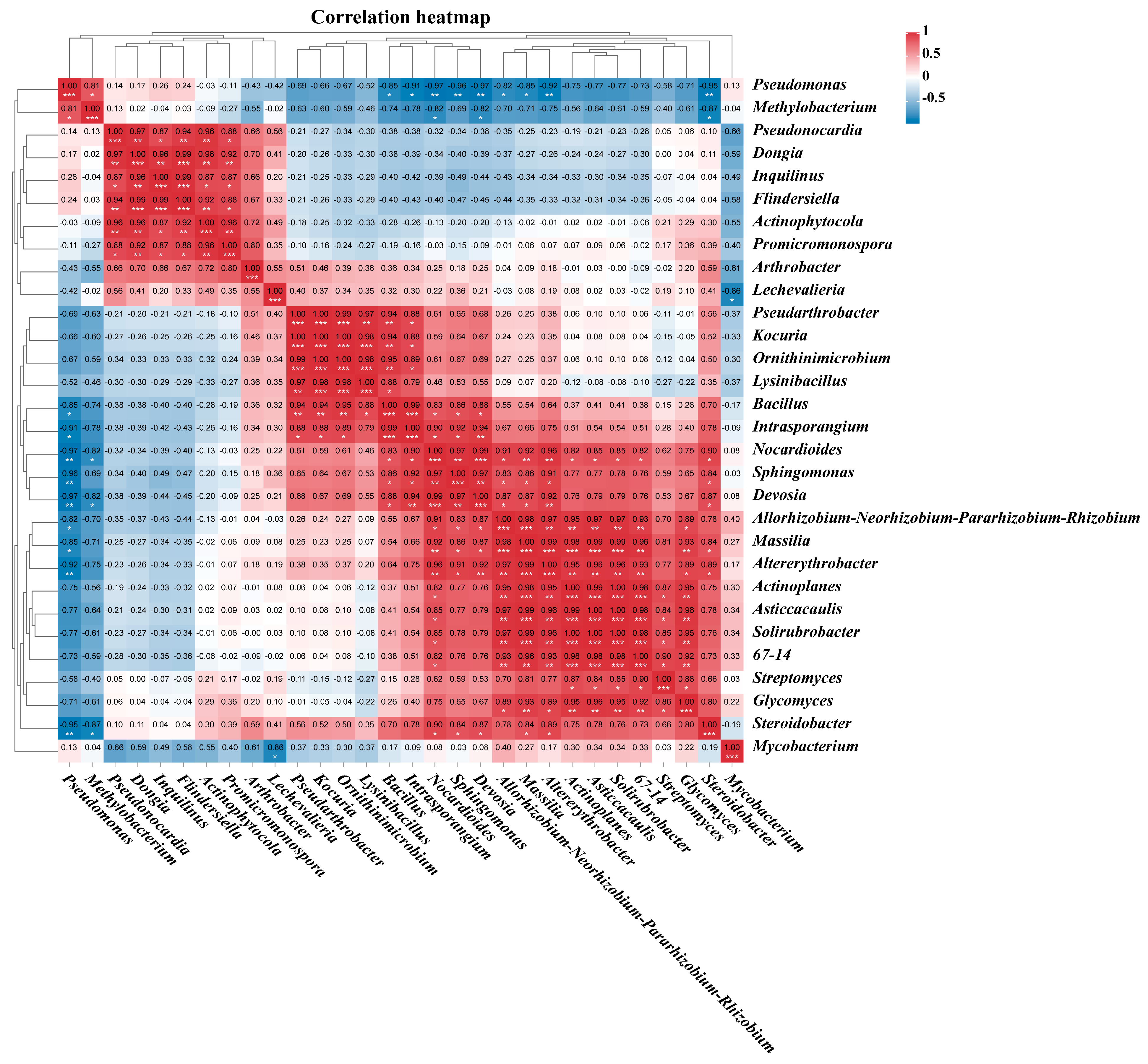
3.7. Correlation Analysis of the Endophytic Bacteria Communities
4. Discussion
4.1. Microbial Community Responses
4.2. Functional Enrichment Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Lai, L.; Zhou, H.; Lai, B. Heterogeneous degradation of bisphenol A by peroxymonosulfate activated with vanadium-titanium magnetite: Performance, transformation pathways and mechanism. Chem. Eng. J. 2018, 349, 633–645. [Google Scholar] [CrossRef]
- Chen, S.Y.; Fu, X.J.; Chu, M.S.; Li, X.Z.; Liu, Z.G.; Tang, J. Carbothermic Reduction Mechanism of Vanadium-titanium Magnetite. J. Iron Steel Res. Int. 2016, 23, 409–414. [Google Scholar] [CrossRef]
- Li, W.; Fu, G.Q.; Chu, M.S.; Zhu, M.Y. Non-isothermal Reduction Behavior and Mechanism of Hongge Vanadium Titanomagnetite Pellet with Simulated Shaft Furnace Gases. Isij Int. 2018, 58, 415–421. [Google Scholar] [CrossRef]
- Feng, C.; Chu, M.-S.; Tang, J.; Liu, Z.-G. Effects of smelting parameters on the slag/metal separation behaviors of Hongge vanadium-bearing titanomagnetite metallized pellets obtained from the gas-based direct reduction process. Int. J. Miner. Met. Mater. 2018, 25, 609–622. [Google Scholar] [CrossRef]
- Li, W.; Wang, N.; Fu, G.Q.; Chu, M.S.; Zhu, M.Y. Influence of roasting characteristics on gas-based direct reduction behavior of Hongge vanadium titanomagnetite pellet with simulated shaft furnace gases. Powder Technol. 2017, 310, 343–350. [Google Scholar] [CrossRef]
- Zhou, M.; Jiang, T.; Yang, S.T.; Xue, X.X. Vanadium-titanium magnetite ore blend optimization for sinter strength based on iron ore basic sintering characteristics. Int. J. Miner. Process. 2015, 142, 125–133. [Google Scholar] [CrossRef]
- Wang, S.; Chen, M.; Guo, Y.; Jiang, T.; Zhao, B. Reduction and Smelting of Vanadium Titanomagnetite Metallized Pellets. JOM 2019, 71, 1144–1149. [Google Scholar] [CrossRef]
- Li, W.; Wang, N.; Fu, G.; Chu, M.; Zhu, M. Effects of Preheating Temperature and Time of Hongge Vanadium Titanomagnetite Pellet on Its Gas-Based Direct Reduction Behavior with Simulated Shaft Furnace Gases. ISIJ Int. 2018, 58, 594–603. [Google Scholar] [CrossRef]
- Yang, J.; Jiang, T.; Ma, S.-H.; Yang, S.-T.; Zhou, M. Kinetics and mechanism of coal-based direct reduction of high-chromium vanadium–titanium magnetite. J. Iron Steel Res. Int. 2022, 29, 1723–1733. [Google Scholar] [CrossRef]
- Li, R.M.; Liu, T.; Zhang, Y.M.; Huang, J.; Xu, C.B. Efficient Extraction of Vanadium from Vanadium-Titanium Magnetite Concentrate by Potassium Salt Roasting Additives. Minerals 2018, 8, 25. [Google Scholar] [CrossRef]
- Moskalyk, R.R.; Alfantazi, A.M. Processing of vanadium: A review. Miner. Eng. 2003, 16, 793–805. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, J.P.; Sun, D.Q.; Li, H.M. Two pass laser welding of TC4 Titanium alloy to 301L stainless steel via pure V interlayer. J. Mater. Res. Technol. 2020, 9, 1400–1404. [Google Scholar] [CrossRef]
- Yu, J.; Hu, N.; Xiao, H.; Gao, P.; Sun, Y. Reduction behaviors of vanadium-titanium magnetite with H2 via a fluidized bed. Powder Technol. 2021, 385, 83–91. [Google Scholar] [CrossRef]
- Lee, J.C.; Kurniawan; Kim, E.Y.; Chung, K.W.; Kim, R.; Jeon, H.S. A review on the metallurgical recycling of vanadium from slags: Towards a sustainable vanadium production. J. Mater. Res. Technol. 2021, 12, 343–364. [Google Scholar] [CrossRef]
- Rack, H.J.; Qazi, J.I. Titanium alloys for biomedical applications. Mater. Sci. Eng. C 2006, 26, 1269–1277. [Google Scholar] [CrossRef]
- Geetha, M.; Singh, A.K.; Asokamani, R.; Gogia, A.K. Ti based biomaterials, the ultimate choice for orthopaedic implants—A review. Prog. Mater. Sci. 2009, 54, 397–425. [Google Scholar] [CrossRef]
- Tang, X.; Huang, Y.; Li, Y.; Yang, Y.; Cheng, X.; Jiao, G.; Dai, H. The response of bacterial communities to V and Cr and novel reducing bacteria near a vanadium-titanium magnetite refinery. Sci. Total Environ. 2022, 806, 151214. [Google Scholar] [CrossRef] [PubMed]
- Gan, C.-D.; Tang, Q.-X.; Wang, H.; Yang, J.-Y.; Nikitin, A. Shewanella oneidensis MR-1 and oxalic acid mediated vanadium reduction and redistribution in vanadium-containing tailings. J. Hazard. Mater. 2023, 451, 131077. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, T.; Lu, L.; Tang, Y. In Vitro Health Risk Assessment of Ingesting Metal-Enriched Soils and Dusts in a Chinese Mining City. Hum. Ecol. Risk Assess. Int. J. 2015, 21, 2005–2021. [Google Scholar] [CrossRef]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef]
- Wang, S.-S.; Liu, J.-M.; Sun, J.; Sun, Y.-F.; Liu, J.-N.; Jia, N.; Fan, B.; Dai, X.-F. Diversity of culture-independent bacteria and antimicrobial activity of culturable endophytic bacteria isolated from different Dendrobium stems. Sci. Rep. 2019, 9, 10389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, Y.; Li, N.; Liu, H.; Zheng, H.; Wang, W.; Liu, Y. High-throughput sequencing-based analysis of the composition and diversity of endophytic bacterial community in seeds of saline-alkali tolerant rice. Microbiol. Res. 2021, 250, 126794. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Su, Q.; Cui, Y.; He, H.; Wang, J.; Zhang, Y.; Zhao, Y.; Abul, H.; Yang, Y.; Long, Y. Temporal and spatial pattern of endophytic fungi diversity of Camellia sinensis (cv. Shu Cha Zao). BMC Microbiol. 2020, 20, 270. [Google Scholar] [CrossRef] [PubMed]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial interactions within the plant holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Aswani, R.; Jishma, P.; Radhakrishnan, E.K. 2—Endophytic bacteria from the medicinal plants and their potential applications. In Microbial Endophytes; Kumar, A., Singh, V.K., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 15–36. [Google Scholar]
- Li, H.; Yan, C.; Tang, Y.; Ma, X.; Chen, Y.; Chen, S.; Lin, M.; Liu, Z. Endophytic bacterial and fungal microbiota in different cultivars of cassava (Manihot esculenta Crantz). J. Microbiol. 2020, 58, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Khan, Q.M.; Sessitsch, A. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants. Chemosphere 2014, 117, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Tardif, S.; Yergeau, É.; Tremblay, J.; Legendre, P.; Whyte, L.G.; Greer, C.W. The Willow Microbiome Is Influenced by Soil Petroleum-Hydrocarbon Concentration with Plant Compartment-Specific Effects. Front. Microbiol. 2016, 7, 186190. [Google Scholar] [CrossRef] [PubMed]
- Liotti, R.G.; da Silva Figueiredo, M.I.; da Silva, G.F.; de Mendonça, E.A.F.; Soares, M.A. Diversity of cultivable bacterial endophytes in Paullinia cupana and their potential for plant growth promotion and phytopathogen control. Microbiol. Res. 2018, 207, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.I.A.; Monteiro, C.; Vega, A.L.; Castro, P.M.L. Endophytic culturable bacteria colonizing Lavandula dentata L. plants: Isolation, characterization and evaluation of their plant growth-promoting activities. Ecol. Eng. 2016, 87, 91–97. [Google Scholar] [CrossRef]
- Knoth, J.L.; Kim, S.H.; Ettl, G.J.; Doty, S.L. Biological nitrogen fixation and biomass accumulation within poplar clones as a result of inoculations with diazotrophic endophyte consortia. New Phytol. 2013, 201, 599–609. [Google Scholar] [CrossRef]
- Shi, Y.W.; Lou, K.; Li, C. Growth promotion effects of the endophyte Acinetobacter johnsonii strain 3-1 on sugar beet. Symbiosis 2011, 54, 159–166. [Google Scholar] [CrossRef]
- Rungin, S.; Indananda, C.; Suttiviriya, P.; Kruasuwan, W.; Jaemsaeng, R.; Thamchaipenet, A. Plant growth enhancing effects by a siderophore-producing endophytic streptomycete isolated from a Thai jasmine rice plant (Oryza sativa L. cv. KDML105). Antonie Leeuwenhoek 2012, 102, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Barnawal, D.; Bharti, N.; Tripathi, A.; Pandey, S.S.; Chanotiya, C.S.; Kalra, A. ACC-Deaminase-Producing Endophyte Brachybacterium paraconglomeratum Strain SMR20 Ameliorates Chlorophytum Salinity Stress via Altering Phytohormone Generation. J. Plant Growth Regul. 2016, 35, 553–564. [Google Scholar] [CrossRef]
- Zouari, I.; Jlaiel, L.; Tounsi, S.; Trigui, M. Biocontrol activity of the endophytic Bacillus amyloliquefaciens strain CEIZ-11 against Pythium aphanidermatum and purification of its bioactive compounds. Biol. Control 2016, 100, 54–62. [Google Scholar] [CrossRef]
- Wicaksono, W.A.; Jones, E.E.; Casonato, S.; Monk, J.; Ridgway, H.J. Biological control of Pseudomonas syringae pv. actinidiae (Psa), the causal agent of bacterial canker of kiwifruit, using endophytic bacteria recovered from a medicinal plant. Biol. Control 2018, 116, 103–112. [Google Scholar] [CrossRef]
- Xie, J.; Shu, P.; Strobel, G.; Chen, J.; Wei, J.; Xiang, Z.; Zhou, Z.; SWg, P.A. promotes disease protection and seedling growth. Biol. Control 2017, 113, 9–17. [Google Scholar] [CrossRef]
- Xu, W.F.; Wang, F.; Zhang, M.; Ou, T.; Wang, R.L.; Strobel, G.; Xiang, Z.H.; Zhou, Z.Y.; Xie, J. Diversity of cultivable endophytic bacteria in mulberry and their potential for antimicrobial and plant growth-promoting activities. Microbiol. Res. 2019, 229, 126328. [Google Scholar] [CrossRef]
- Strobel, G.A. Endophytes as sources of bioactive products. Microbes Infect. 2003, 5, 535–544. [Google Scholar] [CrossRef]
- Rangjaroen, C.; Lumyong, S.; Sloan, W.T.; Sungthong, R. Herbicide-tolerant endophytic bacteria of rice plants as the biopriming agents for fertility recovery and disease suppression of unhealthy rice seeds. BMC Plant Biol. 2019, 19, 580. [Google Scholar] [CrossRef]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.; Dunfield, K.E.J.A.J.O.B. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef]
- Xia, F.; Chen, X.; Guo, M.-Y.; Bai, X.-H.; Liu, Y.; Shen, G.-R.; Li, Y.-L.; Lin, J.; Zhou, X.-W. High-throughput sequencing-based analysis of endogenetic fungal communities inhabiting the Chinese Cordyceps reveals unexpectedly high fungal diversity. Sci. Rep. 2016, 6, 33437. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Beiko, R.G.; Langille, M.G.I. Predicting the Functional Potential of the Microbiome from Marker Genes Using PICRUSt. Methods Mol. Biol. 2018, 1849, 169–177. [Google Scholar]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- He, Z.; Yue, H.-R.; Xue, X. Study of the High Temperature Metallurgical Properties of On-Site Samples with Vanadium–Titanium Magnetite. Trans. Indian Inst. Met. 2018, 71, 2001–2013. [Google Scholar] [CrossRef]
- Gan, C.D.; Cui, S.F.; Wu, Z.Z.; Yang, J.Y. Multiple heavy metal distribution and microbial community characteristics of vanadium-titanium magnetite tailing profiles under different management modes. J. Hazard. Mater. 2022, 429, 128032. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef] [PubMed]
- White, J.F.; Kingsley, K.L.; Zhang, Q.; Verma, R.; Obi, N.; Dvinskikh, S.; Elmore, M.T.; Verma, S.K.; Gond, S.K.; Kowalski, K.P. Review: Endophytic microbes and their potential applications in crop management. Pest Manag. Sci. 2019, 75, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, S. Bioremediation of heavy metals from industrial effluents by endophytes and their metabolic activity: Recent advances. Bioresour. Technol. 2021, 339, 125589. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xiang, P.; Li, L.; Zhang, T.; Wu, Q.; Bao, Z.; Tu, W.; Zhao, C. Phosphorus mining activities alter endophytic bacterial communities and metabolic functions of surrounding vegetables and crops. Plant Soil 2023, 497, 155–174. [Google Scholar] [CrossRef]
- Jose, J.; Giridhar, R.; Anas, A.; Bharathi, P.A.L.; Nair, S. Heavy metal pollution exerts reduction/adaptation in the diversity and enzyme expression profile of heterotrophic bacteria in Cochin estuary, India. Environ. Pollut. 2011, 159, 2775–2780. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, S.; Diao, M.; Fu, J.; Xie, M.; Shi, J.; Liu, Z.; Jiang, Y.; Cao, X.; Borthwick, A.G.L. Microbial Community Responses to Vanadium Distributions in Mining Geological Environments and Bioremediation Assessment. J. Geophys. Res. Biogeosci. 2019, 124, 601–615. [Google Scholar] [CrossRef]
- Tang, Q.-X.; Gan, C.-D.; Yang, J.-Y.; Huang, Y. Dynamics of vanadium and response of inherent bacterial communities in vanadium-titanium magnetite tailings to beneficiation agents, temperature, and illumination. Environ. Pollut. 2023, 330, 121743. [Google Scholar] [CrossRef]
- Li, Q.; Xiong, Z.; Xiang, P.; Zhou, L.; Zhang, T.; Wu, Q.; Zhao, C. Effects of uranium mining on soil bacterial communities and functions in the Qinghai-Tibet plateau. Chemosphere 2024, 347, 140715. [Google Scholar] [CrossRef]
- Guan, N.; Li, J.; Shin, H.-D.; Du, G.; Chen, J.; Liu, L. Microbial response to environmental stresses: From fundamental mechanisms to practical applications. Appl. Microbiol. Biotechnol. 2017, 101, 3991–4008. [Google Scholar] [CrossRef] [PubMed]
- Xiang, P.; Liao, W.; Xiong, Z.; Xiao, W.; Luo, Y.; Peng, L.; Zou, L.; Zhao, C.; Li, Q. Effects of polystyrene microplastics on the agronomic traits and rhizosphere soil microbial community of highland barley. Sci. Total Environ. 2023, 907, 167986. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Zhang, Q.; Chen, X.; Dong, F.; Chen, H.; Liu, M.; Ali, I. Speciation Distribution of Heavy Metals in Uranium Mining Impacted Soils and Impact on Bacterial Community Revealed by High-Throughput Sequencing. Front. Microbiol. 2019, 10, 1867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, S.; Liu, Z.; Li, Y.; Wang, Q.; Zhang, X.; Li, M.; Zhang, B. Community assembly and microbial interactions in an alkaline vanadium tailing pond. Environ. Res. 2024, 246, 118104. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Barker-Finkel, J.; Galloway, T.S.; Handy, R.D.; Ford, T.E. Temporal bacterial diversity associated with metal-contaminated river sediments. Ecotoxicology 2010, 19, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Liang, X.; Guo, T.; Wu, T.; Chai, B. Bacterial community succession and influencing factors for Imperata cylindrica litter decomposition in a copper tailings area of China. Sci. Total Environ. 2022, 815, 152908. [Google Scholar] [CrossRef] [PubMed]
- Balíková, K.; Vojtková, H.; Duborská, E.; Kim, H.; Matús, P.; Urík, M. Role of Exopolysaccharides of Pseudomonas in Heavy Metal Removal and Other Remediation Strategies. Polymers 2022, 14, 4253. [Google Scholar] [CrossRef] [PubMed]
- Fakhar, A.; Gul, B.; Gurmani, A.R.; Khan, S.M.; Ali, S.; Sultan, T.; Chaudhary, H.J.; Rafique, M.; Rizwan, M. Heavy metal remediation and resistance mechanism of Aeromonas, Bacillus, and Pseudomonas: A review. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1868–1914. [Google Scholar] [CrossRef]
- Saha, J.; Chaki, M.G.; Karmakar, S.; Chatterjee, A.; Pal, A. Effect of different heavy metals on lipase production by a multiple heavy metal-resistant Pseudomonas aeruginosa strain isolated from arable land. Biologia 2023, 78, 2975–2985. [Google Scholar] [CrossRef]
- Tripathi, M.; Kumar, S.; Makarana, G.; Goel, R. Metal-Tolerant Bioinoculant Pseudomonas putida KNP9 Mediated Enhancement of Soybean Growth under Heavy Metal Stress Suitable for Biofuel Production at the Metal-Contaminated Site. Energies 2023, 16, 4508. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, S.; Zhong, Q.; Gong, G.; Wang, G.; Guo, X.; Xu, X. Effects of soil chemical properties and fractions of Pb, Cd, and Zn on bacterial and fungal communities. Sci. Total Environ. 2020, 715, 136904. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, B.; He, C.; Shi, J.; Borthwick, A.G.L.; Huang, X. Synchronous microbial vanadium (V) reduction and denitrification in groundwater using hydrogen as the sole electron donor. Water Res. 2018, 141, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, C.; Xie, Y.; Luo, Y.; Sheng, M.; Xu, F.; Xu, H. Ecological responses of soil microbial abundance and diversity to cadmium and soil properties in farmland around an enterprise-intensive region. J. Hazard. Mater. 2020, 392, 122478. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, T.; Selvarajan, R.; Msagati, T.; Venkatachalam, S.; Meddows-Taylor, S. Defunct gold mine tailings are natural reservoir for unique bacterial communities revealed by high-throughput sequencing analysis. Sci. Total Environ. 2019, 650, 2199–2209. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Luo, H.; Wang, X.; Liu, H.; Peng, H.; Sheng, M.; Xu, F.; Xu, H. Effects of environmental factors on soil bacterial community structure and diversity in different contaminated districts of Southwest China mine tailings. Sci. Total Environ. 2021, 802, 149899. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.-H.; Lu, C.-W.; Liu, F.-G.; Kao, C.-M.; Chen, S.C. Draft genome sequence of Pseudomonas sp. A46 isolated from mercury-contaminated wastewater. J. Basic Microbiol. 2022, 62, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zeng, Z.; Wang, Y.; He, G.; Hu, Y.; Gao, D.; Dai, Y.; Li, Q.; Zhang, H. Ecological risk assessment and identification of the distinct microbial groups in heavy metal-polluted river sediments. Environ. Geochem. Health 2023, 45, 1311–1329. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-K.; Park, Y.-J.; Kim, M.; Kim, M.-C.; Ibal, J.C.; Ung, K.G.; Lee, G.-D.; Tagele, S.B.; Kwon, H.-J.; Kang, M.-S.; et al. Complete genome sequence of a plant growth-promoting bacterium Pseudarthrobacter sp. NIBRBAC000502772, isolated from shooting range soil in the Republic of Korea. Korean J. Microbiol. 2020, 56, 390–393. [Google Scholar]
- Wang, T.-J.; Su, N.-N.; Lei, P.; Qiu, M.-Y.; Chen, Z.-J.; Yao, L.-G.; Han, H. Community Structure of Heavy Metal Immobilized Bacteria in the Lettuce (Lactuca sativa L.) Rhizosphere in Soil Polluted by Heavy Metals and Its Effects on Reducing Heavy Metal Accumulation in Lettuce. Huan Jing Ke Xue = Huanjing Kexue 2019, 40, 5133–5141. [Google Scholar]
- Mano, H.; Morisaki, H. Endophytic bacteria in the rice plant. Microbes Environ. 2008, 23, 109–117. [Google Scholar] [CrossRef]
- Jacquiod, S.; Nunes, I.; Brejnrod, A.; Hansen, M.A.; Holm, P.E.; Johansen, A.; Brandt, K.K.; Priemé, A.; Sorensen, S.J. Long-term soil metal exposure impaired temporal variation in microbial metatranscriptomes and enriched active phages. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Longhi, C.; Maurizi, L.; Conte, A.L.; Marazzato, M.; Comanducci, A.; Nicoletti, M.; Zagaglia, C. Extraintestinal Pathogenic Escherichia coli: Beta-Lactam Antibiotic and Heavy Metal Resistance. Antibiotics 2022, 11, 328. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.L.; Tian, X.; Nie, G.X.; Wang, J.L.; Liu, M.; Jiang, K.Y.; Wang, B.J.; Guo, Q.Q.; Huang, J.R.; Wang, L. The transcriptomic response to copper exposure in the digestive gland of Japanese scallops (Mizuhopecten yessoensis). Fish Shellfish. Immunol. 2015, 46, 161–167. [Google Scholar] [CrossRef]
- Yao, Q.; Yang, A.J.; Hu, X.; Zou, H.T.; Chen, J.F.; Li, Q.; Lv, S.H.; Yu, X.G.; Li, C.X. Effects of antimony exposure on DNA damage and genome-wide variation in zebrafish (Danio rerio) liver. Aquat. Toxicol. 2023, 259, 106524. [Google Scholar] [CrossRef]
- Kaur, J.; Reinhardt, D.P.; Chapter; Vishwakarma, A.; Sharpe, P.; Shi, S.; Ramalingam, M. (Eds.) Stem Cell Biology and Tissue Engineering in Dental Sciences; Academic Press: Boston, MA, USA, 2015; pp. 25–45. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Z.; Zhang, Y.; Chen, X.; Sha, A.; Xiao, W.; Luo, Y.; Peng, L.; Zou, L.; Li, Q. Impact of Vanadium–Titanium–Magnetite Mining Activities on Endophytic Bacterial Communities and Functions in the Root Systems of Local Plants. Genes 2024, 15, 526. https://doi.org/10.3390/genes15050526
Xiong Z, Zhang Y, Chen X, Sha A, Xiao W, Luo Y, Peng L, Zou L, Li Q. Impact of Vanadium–Titanium–Magnetite Mining Activities on Endophytic Bacterial Communities and Functions in the Root Systems of Local Plants. Genes. 2024; 15(5):526. https://doi.org/10.3390/genes15050526
Chicago/Turabian StyleXiong, Zhuang, Yunfeng Zhang, Xiaodie Chen, Ajia Sha, Wenqi Xiao, Yingyong Luo, Lianxin Peng, Liang Zou, and Qiang Li. 2024. "Impact of Vanadium–Titanium–Magnetite Mining Activities on Endophytic Bacterial Communities and Functions in the Root Systems of Local Plants" Genes 15, no. 5: 526. https://doi.org/10.3390/genes15050526