The Chloroplast Genome Sequence of Scutellaria baicalensis Provides Insight into Intraspecific and Interspecific Chloroplast Genome Diversity in Scutellaria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling, DNA Extraction and Sequencing
2.2. Chloroplast Genome Assembly and Annotation
2.3. Codon Usage
2.4. Analysis of Tandem Repeats and Microsatellites
2.5. Interspecific Comparison
2.6. Intraspecific Comparison
2.7. Phylogenetic Reconstruction
3. Results
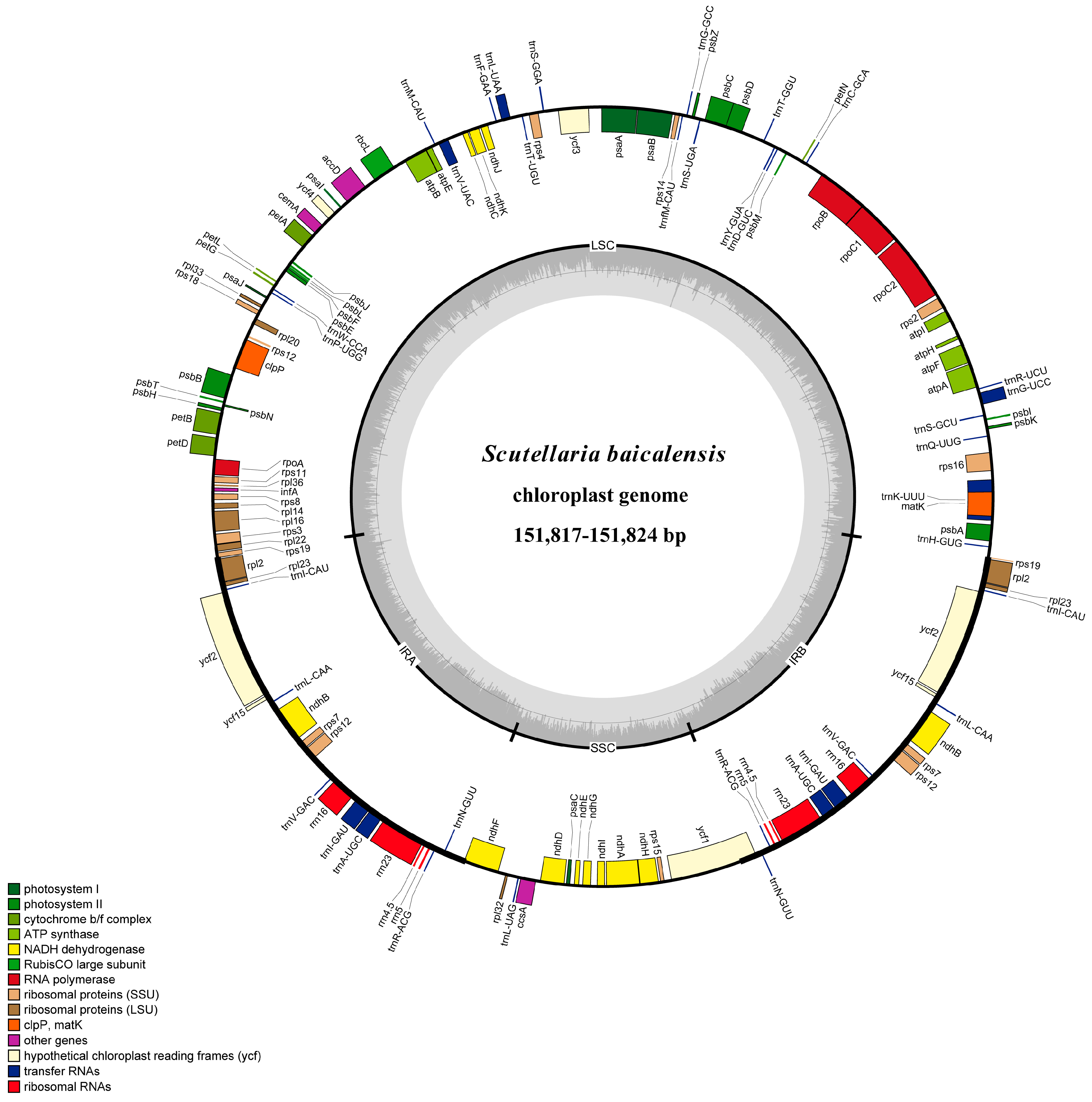
3.1. Genome Size and Features
3.2. Codon Usage
3.3. Repeat and Microsatellites Analysis
3.4. Interspecific Comparison
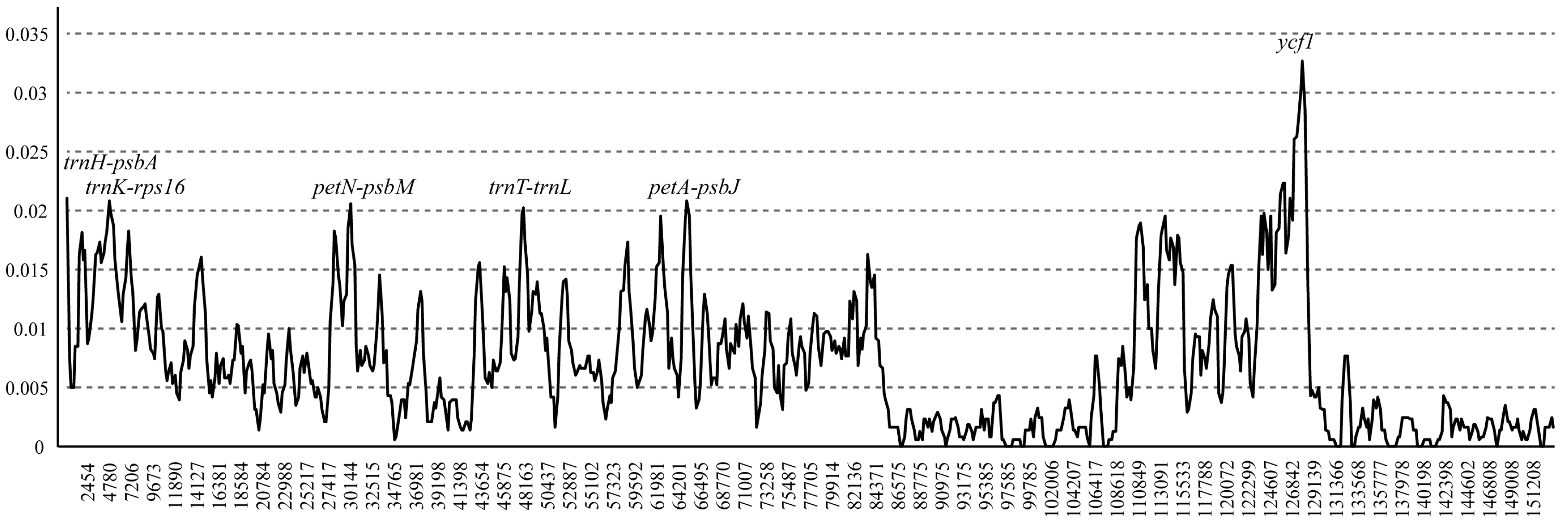
3.5. Intraspecific Comparison
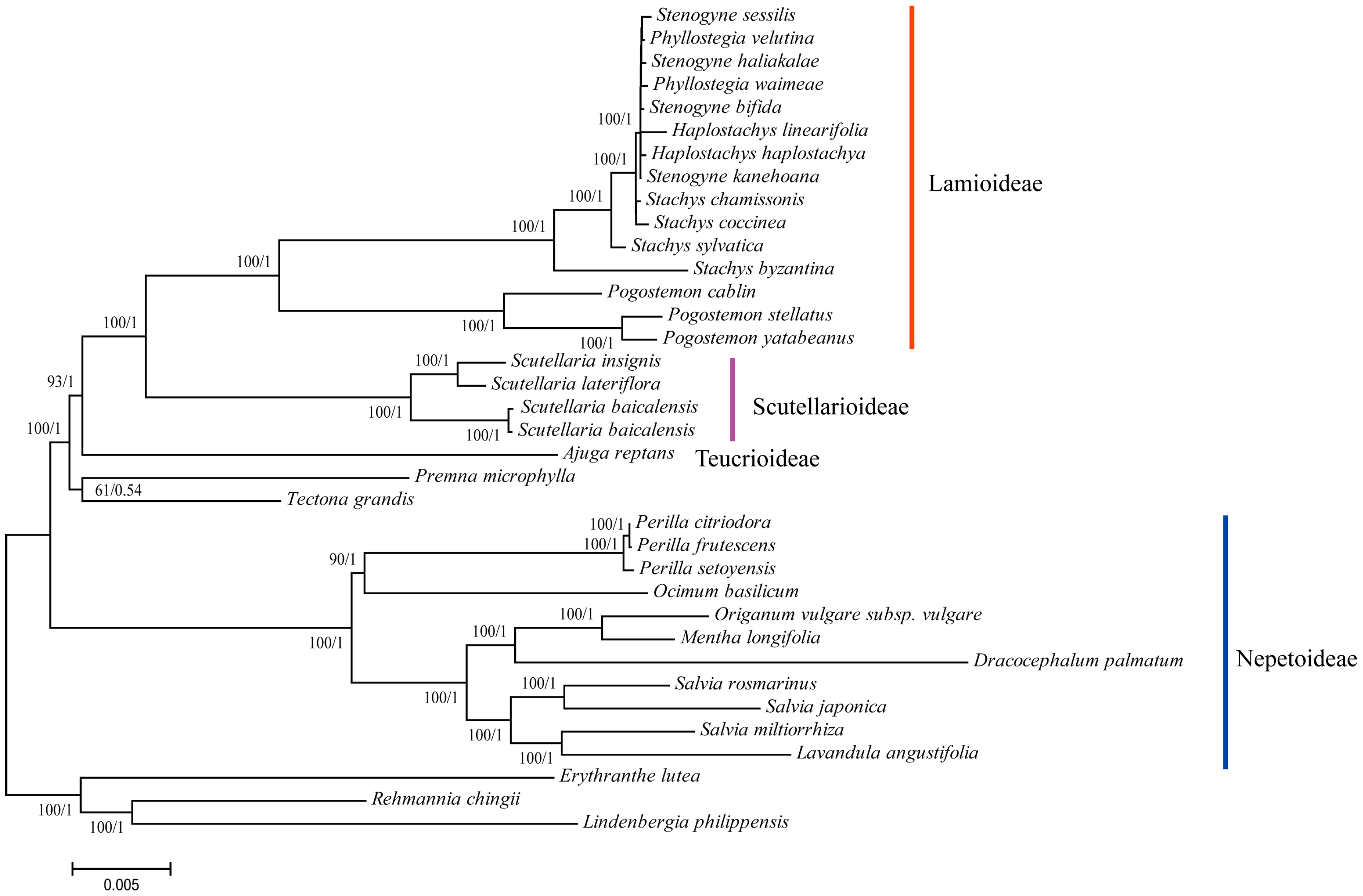
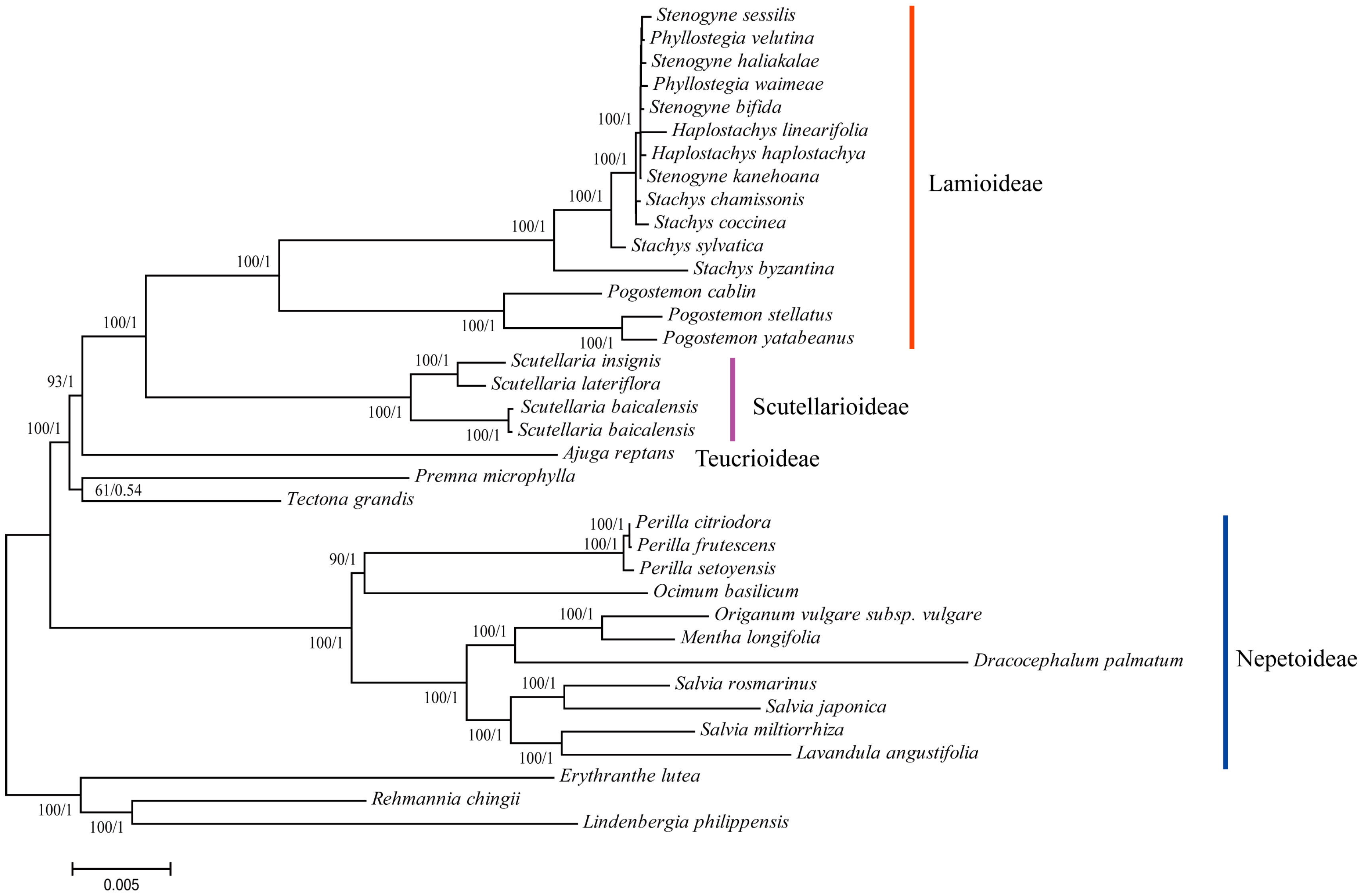
3.6. Phylogenetic Reconstruction of Lamiaceae
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Y.; Shan, L.; Hua, Y.; Wang, D.; Zeng, H.; Liu, R.; Zhang, W.; Hu, Z. Baicalein selectively induces apoptosis in activated lymphocytes and ameliorates concanavalin a-induced hepatitis in mice. PLoS ONE 2013, 8, e69592. [Google Scholar] [CrossRef] [PubMed]
- Huynh, D.L.; Sharma, N.; Kumar Singh, A.; Singh Sodhi, S.; Zhang, J.J.; Mongre, R.K.; Ghosh, M.; Kim, N.; Ho Park, Y.; Kee Jeong, D. Anti-tumor activity of wogonin, an extract from Scutellaria baicalensis, through regulating different signaling pathways. Chin. J. Nat. Med. 2017, 15, 15–40. [Google Scholar] [CrossRef]
- Lai, C.C.; Huang, P.H.; Yang, A.H.; Chiang, S.C.; Tang, C.Y.; Tseng, K.W.; Huang, C.H. Baicalein attenuates lung injury induced by myocardial ischemia and reperfusion. Am. J. Chin. Med. 2017, 45, 791–811. [Google Scholar] [CrossRef] [PubMed]
- Li, H.D.; Chen, X.; Yang, Y.; Huang, H.M.; Zhang, L.; Zhang, X.; Zhang, L.; Huang, C.; Meng, X.M.; Li, J. Wogonin attenuates inflammation by activating ppar-gamma in alcoholic liver disease. Int. Immunopharmacol. 2017, 50, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.; Zhou, H. Guidelines on Modern Cultivation, Breeding and Processing of Chinese Materia Medica; China Traditional Chinese Medicine Press: Beijing, China, 1999. [Google Scholar]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The molecular genetics of crop domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, X.; Su, W.; Zhang, G.; Zhou, R. DNA barcodes for discriminating the medicinal plant Scutellaria baicalensis (Lamiaceae) and its adulterants. Biol. Pharm. Bull. 2011, 34, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Long, P.; Jiang, C.; Li, M.; Huang, L. Development and characterization of simple sequence repeat (SSR) markers based on a full-length cdna library of scutellaria baicalensis. Genomics 2015, 105, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.J.; Zhang, Z.Y.; Hu, J.; Guo, L.P.; Shao, A.J.; Huang, L.Q. Impacts of recent cultivation on genetic diversity pattern of a medicinal plant, Scutellaria baicalensis (Lamiaceae). BMC Genet. 2010, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yuan, Q.; Jiang, D.; Zhang, Y.; Huang, L. Development and characterisation of microsatellite markers for the medicinal plant Scutellaria baicalensis (Lamiaceae). Biochem. Syst. Ecol. 2014, 54, 267–271. [Google Scholar] [CrossRef]
- Han, J.-P.; Shi, L.-C.; Chen, X.-C.; Lin, Y.-L. Comparison of four DNA barcodes in identifying certain medicinal plants of Lamiaceae. J. Syst. Evol. 2012, 50, 227–234. [Google Scholar] [CrossRef]
- Pogson, B.J.; Albrecht, V. Genetic dissection of chloroplast biogenesis and development: An overview. Plant Phys. 2011, 155, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Cheng, T.; Lin, K.; Zhou, S. Sequencing angiosperm plastid genomes made easy: A complete set of universal primers and a case study on the phylogeny of saxifragales. Genome Biol. Evol. 2013, 5, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhan, D.-F.; Jia, X.; Dai, H.-F.; Mei, W.-L.; Chen, X.-T.; Peng, S.-Q. Complete chloroplast genome sequence of Aquilaria sinensis (Lour.) gilg and the evolution analysis within the malvalesorder. Front. Plant Sci. 2016, 7, 280. [Google Scholar] [PubMed]
- Yang, Y.; Zhou, T.; Duan, D.; Yang, J.; Feng, L.; Zhao, G. Comparative analysis of the complete chloroplast genomes of five Quercus species. Front. Plant Sci. 2016, 7, 959. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. Ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Li, W.; Xie, X.; Lu, Y.; Liu, Y.; Jin, X.; Suo, Z. Phylogenetic resolution in juglans based on complete chloroplast genomes and nuclear DNA sequences. Front. Plant Sci. 2017, 8, 1148. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Cheng, T.; Li, C.; Xu, C.; Long, P.; Chen, C.; Zhou, S. Discriminating plants using the DNA barcode rbclb: An appraisal based on a large dataset. Mol. Ecol. Resour. 2014, 14, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Dong, W.; Liu, B.; Xu, C.; Yao, X.; Gao, J.; Corlett, R.T. Comparative analysis of complete chloroplast genome sequences of two tropical trees Machilus yunnanensis and Machilus balansae in the family lauraceae. Front. Plant Sci. 2015, 6, 662. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Dong, W.; Li, W.; Lu, Y.; Xie, X.; Jin, X.; Shi, J.; He, K.; Suo, Z. Comparative analysis of six Lagerstroemia complete chloroplast genomes. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Perdereau, A.; Klaas, M.; Barth, S.; Hodkinson, T.R. Plastid genome sequencing reveals biogeographical structure and extensive population genetic variation in wild populations of phalaris arundinacea l. In north-western europe. GCB Bioenergy 2017, 9, 46–56. [Google Scholar] [CrossRef]
- Curci, P.L.; De Paola, D.; Sonnante, G. Development of chloroplast genomic resources for cynara. Mol. Ecol. Resour. 2016, 16, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.-F.; Hinsinger, D.D.; Strijk, J.S. Comparison of intraspecific, interspecific and intergeneric chloroplast diversity in cycads. Sci. Rep. 2016, 6, 31473. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, S.; Li, F.; Zhang, S.; Zhang, H.; Wang, X.; Sun, R.; Bonnema, G.; Borm, T.J.A. A phylogenetic analysis of chloroplast genomes elucidates the relationships of the six economically important Brassica species comprising the triangle of u. Front. Plant Sci. 2017, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J. DNA protocols for plants-ctab total DNA isolation. In Molecular Techniques in Taxonomy; Springer: Berlin, Germnay, 1991; pp. 283–293. [Google Scholar]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with dogma. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed]
- Conant, G.C.; Wolfe, K.H. Genomevx: Simple web-based creation of editable circular chromosome maps. Bioinformatics 2008, 24, 861–862. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- MIcroSAtellite. Available online: http://pgrc.ipk-gatersleben.de/misa (accessed on 30 July 2017).
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. Reputer: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. Vista: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. Mafft multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Se-Al: Sequence Alignment Editor; Version 2.0. Available online: http://tree.bio.ed.ac.uk/software/ (accessed on 10 September 2017).
- Librado, P.; Rozas, J. Dnasp v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Ruhfel, B.R.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E.; Burleigh, J.G. From algae to angiosperms-inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes. BMC Evol. Biol. 2014, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Santorum, J.M.; Darriba, D.; Taboada, G.L.; Posada, D. Jmodeltest.Org: Selection of nucleotide substitution models on the cloud. Bioinformatics 2014, 30, 1310–1311. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Lukas, B.; Novak, J. The complete chloroplast genome of Origanum vulgare L. (Lamiaceae). Gene 2013, 528, 163. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Song, J.; Gao, H.; Zhu, Y.; Xu, J.; Pang, X.; Yao, H.; Sun, C.; Li, X.; Li, C.; et al. The complete chloroplast genome sequence of the medicinal plant Salvia miltiorrhiza. PLoS ONE 2013, 8, e57607. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Cantino, P.D.; Olmstead, R.G.; Bramley, G.L.; Xiang, C.L.; Ma, Z.H.; Tan, Y.H.; Zhang, D.X. A large-scale chloroplast phylogeny of the lamiaceae sheds new light on its subfamilial classification. Sci. Rep. 2016, 6, 34343. [Google Scholar] [CrossRef] [PubMed]
- Welch, A.J.; Collins, K.; Ratan, A.; Drautz-Moses, D.I.; Schuster, S.C.; Lindqvist, C. The quest to resolve recent radiations: Plastid phylogenomics of extinct and endangered hawaiian endemic mints (Lamiaceae). Mol. Phylogenet. Evol. 2016, 99, 16–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Vázquez, L.; Chen, X.; Li, H.; Zhang, H.; Liu, Z.; Zhao, G. Development of chloroplast and nuclear DNA markers for chinese oaks (Quercus subgenus Quercus) and assessment of their utility as DNA barcodes. Front. Plant Sci. 2017, 8, 816. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.A.; Donoghue, M.J. Rates of molecular evolution are linked to life history in flowering plants. Science 2008, 322, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Welch, J.J.; Bromham, L. Watching the clock: Studying variation in rates of molecular evolution between species. Trends Ecol. Evol. 2010, 25, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liang, D.; Gao, T.; Pang, X.; Song, J.; Yao, H.; Han, J.; Liu, Z.; Guan, X.; Jiang, K.; et al. Ptigs-idit, a system for species identification by DNA sequences of the psba-trnh intergenic spacer region. BMC Bioinform. 2011, 12 (Suppl. 13), S4. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Liu, C.; Shi, L.; Liu, R.; Liang, D.; Li, H.; Cherny, S.S.; Chen, S. Utility of the trnh-psba intergenic spacer region and its combinations as plant DNA barcodes: A meta-analysis. PLoS ONE 2012, 7, e48833. [Google Scholar] [CrossRef] [PubMed]
- Wills, D.M.; Hester, M.L.; Liu, A.Z.; Burke, J.M. Chloroplast SSR polymorphisms in the compositae and the mode of organellar inheritance in helianthus annuus. Theor. Appl. Genet. 2005, 110, 941–947. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xiao, H.; Deng, C.; Xiong, L.; Yang, J.; Peng, C. The complete chloroplast genome sequences of the medicinal plant pogostemon cablin. Int. J. Mol. Sci. 2016, 17, 820. [Google Scholar] [CrossRef] [PubMed]
- Kelchner, S.A. The evolution of non-coding chloroplast DNA and its application in plant systematics. Ann. Mo. Bot. Gard. 2000, 87, 482–498. [Google Scholar] [CrossRef]
- Borsch, T.; Quandt, D. Mutational dynamics and phylogenetic utility of noncoding chloroplast DNA. Plant Syst. Evol. 2009, 282, 169–199. [Google Scholar] [CrossRef]
- Ingvarsson, P.K.; Ribstein, S.; Taylor, D.R. Molecular evolution of insertions and deletion in the chloroplast genome of silene. Mol. Biol. Evol. 2003, 20, 1737–1740. [Google Scholar] [CrossRef] [PubMed]
- Lohne, C.; Borsch, T. Molecular evolution and phylogenetic utility of the petd group ii intron: A case study in basal angiosperms. Mol. Biol. Evol. 2005, 22, 317–332. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | BOP028265 | BOP025266 |
|---|---|---|
| Total cpDNA size (bp) | 151,824 | 151,817 |
| LSC size (bp) | 83,976 | 83,960 |
| IR size (bp) | 25,255 | 25,263 |
| SSC size (bp) | 17,338 | 17,331 |
| Number of genes | 114 | 114 |
| Number of different protein-coding genes | 80 | 80 |
| Number of different tRNA genes | 30 | 30 |
| Number of different rRNA genes | 4 | 4 |
| Number of different duplicated genes | 17 | 17 |
| GC content | 38.3% | 38.3% |
| GC content of LSC | 36.3% | 36.3% |
| GC content of SSC | 32.7% | 32.7% |
| GC content of IR | 43.6% | 43.6% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, D.; Zhao, Z.; Zhang, T.; Zhong, W.; Liu, C.; Yuan, Q.; Huang, L. The Chloroplast Genome Sequence of Scutellaria baicalensis Provides Insight into Intraspecific and Interspecific Chloroplast Genome Diversity in Scutellaria. Genes 2017, 8, 227. https://doi.org/10.3390/genes8090227
Jiang D, Zhao Z, Zhang T, Zhong W, Liu C, Yuan Q, Huang L. The Chloroplast Genome Sequence of Scutellaria baicalensis Provides Insight into Intraspecific and Interspecific Chloroplast Genome Diversity in Scutellaria. Genes. 2017; 8(9):227. https://doi.org/10.3390/genes8090227
Chicago/Turabian StyleJiang, Dan, Zhenyu Zhao, Teng Zhang, Wenhao Zhong, Chunsheng Liu, Qingjun Yuan, and Luqi Huang. 2017. "The Chloroplast Genome Sequence of Scutellaria baicalensis Provides Insight into Intraspecific and Interspecific Chloroplast Genome Diversity in Scutellaria" Genes 8, no. 9: 227. https://doi.org/10.3390/genes8090227
APA StyleJiang, D., Zhao, Z., Zhang, T., Zhong, W., Liu, C., Yuan, Q., & Huang, L. (2017). The Chloroplast Genome Sequence of Scutellaria baicalensis Provides Insight into Intraspecific and Interspecific Chloroplast Genome Diversity in Scutellaria. Genes, 8(9), 227. https://doi.org/10.3390/genes8090227