Genome-Wide Identification and Analysis of Arabidopsis Sodium Proton Antiporter (NHX) and Human Sodium Proton Exchanger (NHE) Homologs in Sorghum bicolor

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Abiotic Stress Treatments
2.2. Identification and Characterization of Sodium Transporters
2.3. Phylogeny, Divergence, Promoter Analysis, Physical Gene Mapping, and Co-Expression Analysis
2.4. RNA Isolation and Quantitative Real-Time PCR Analysis
3. Results
3.1. Identification and Characterization of NHX/NHE Transporters in Sorghum
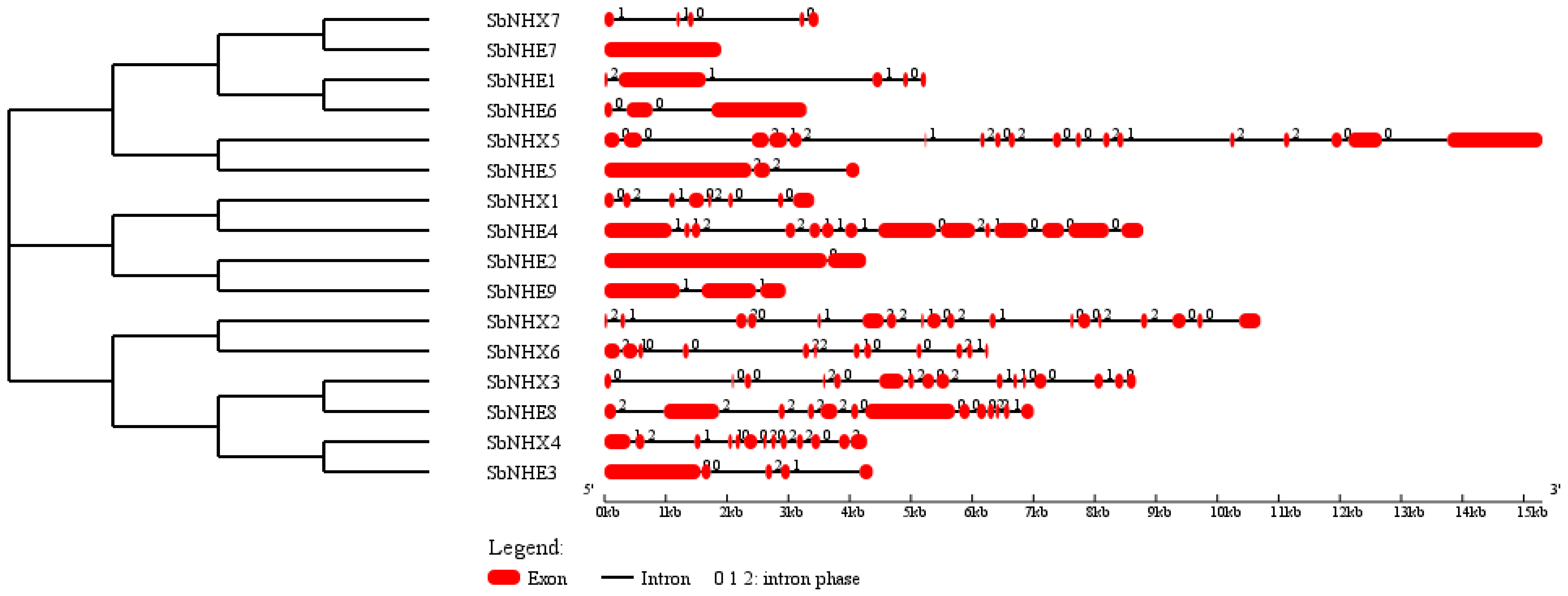
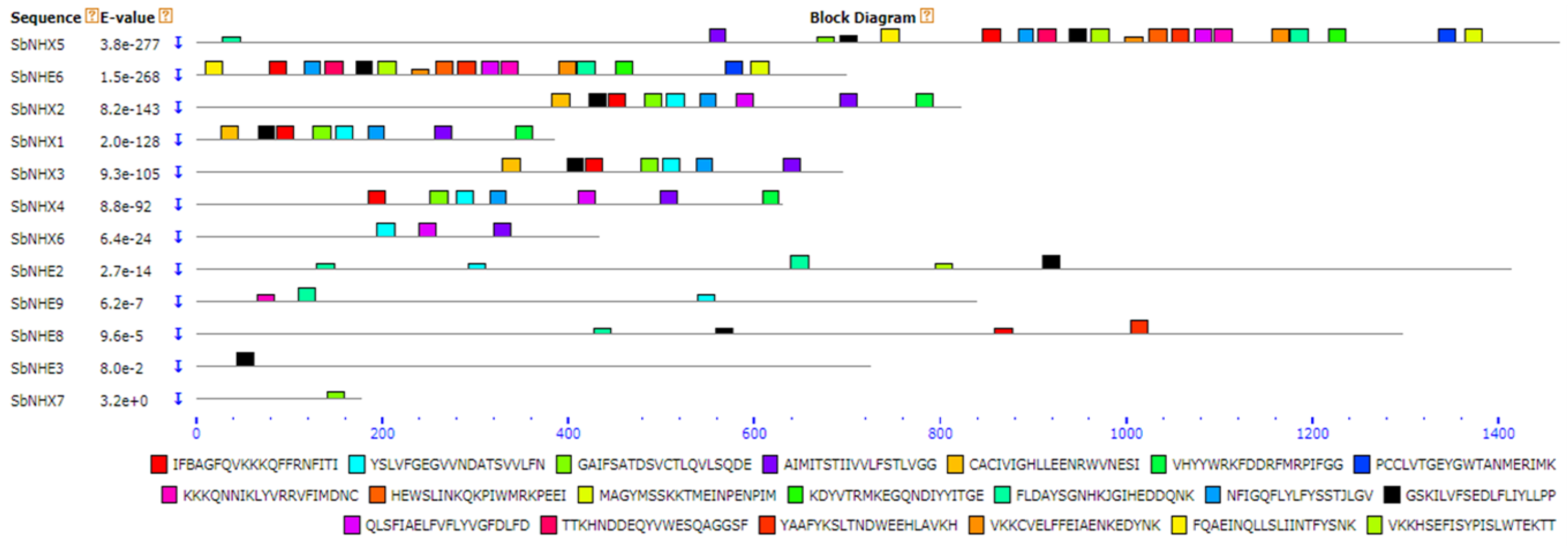
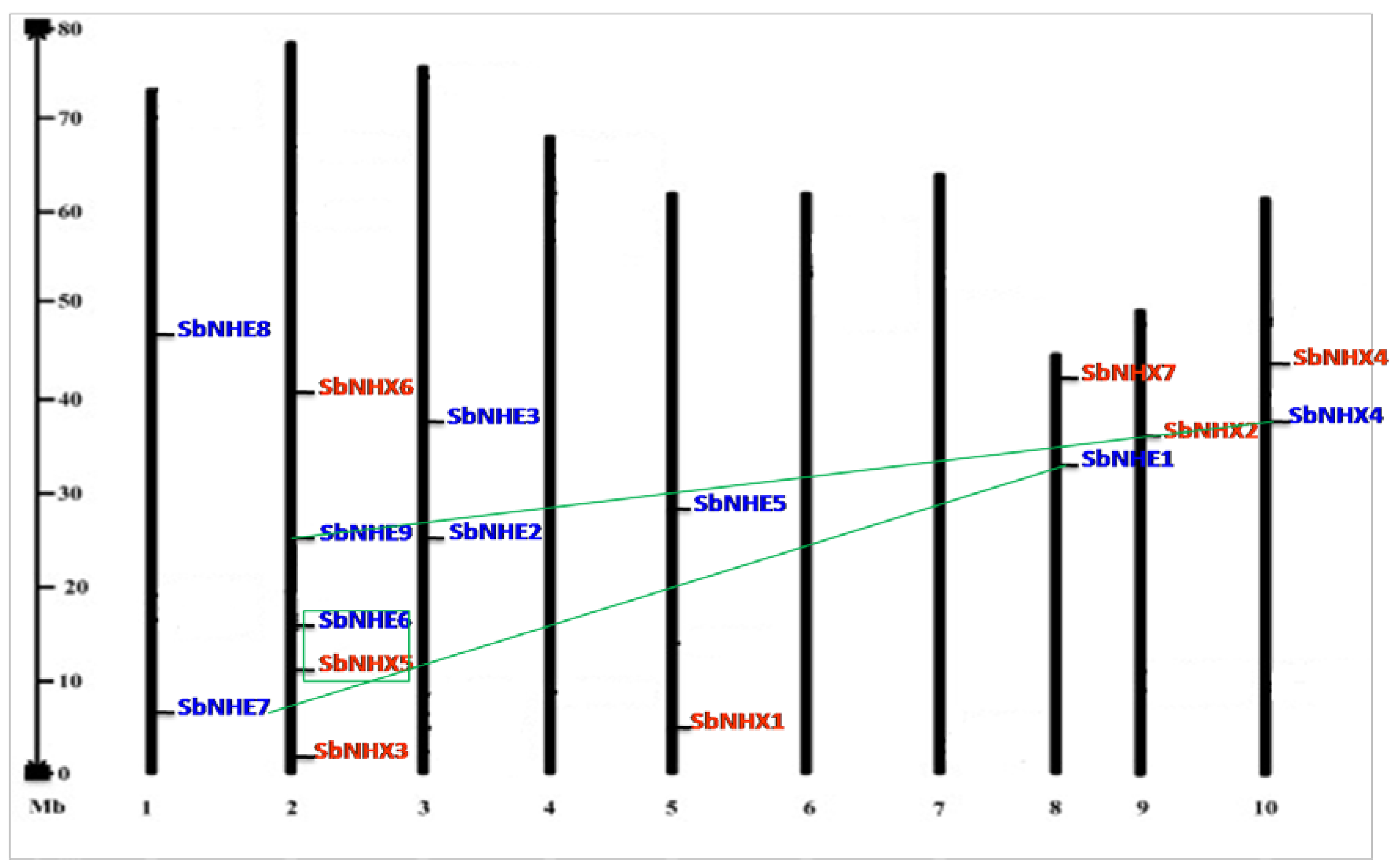
3.2. Phylogeny, Divergence, Promoter Analysis, and Physical Genome Mapping
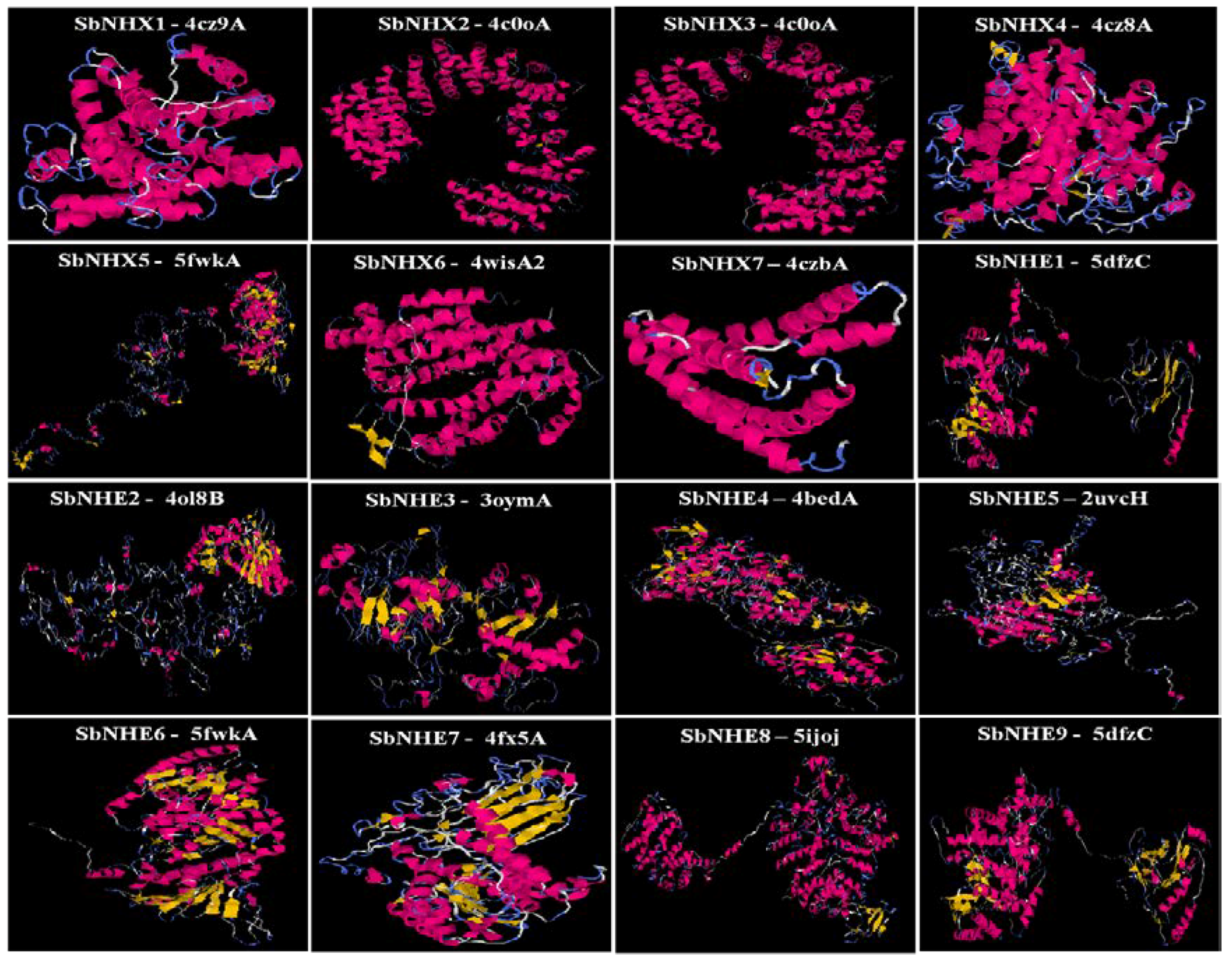
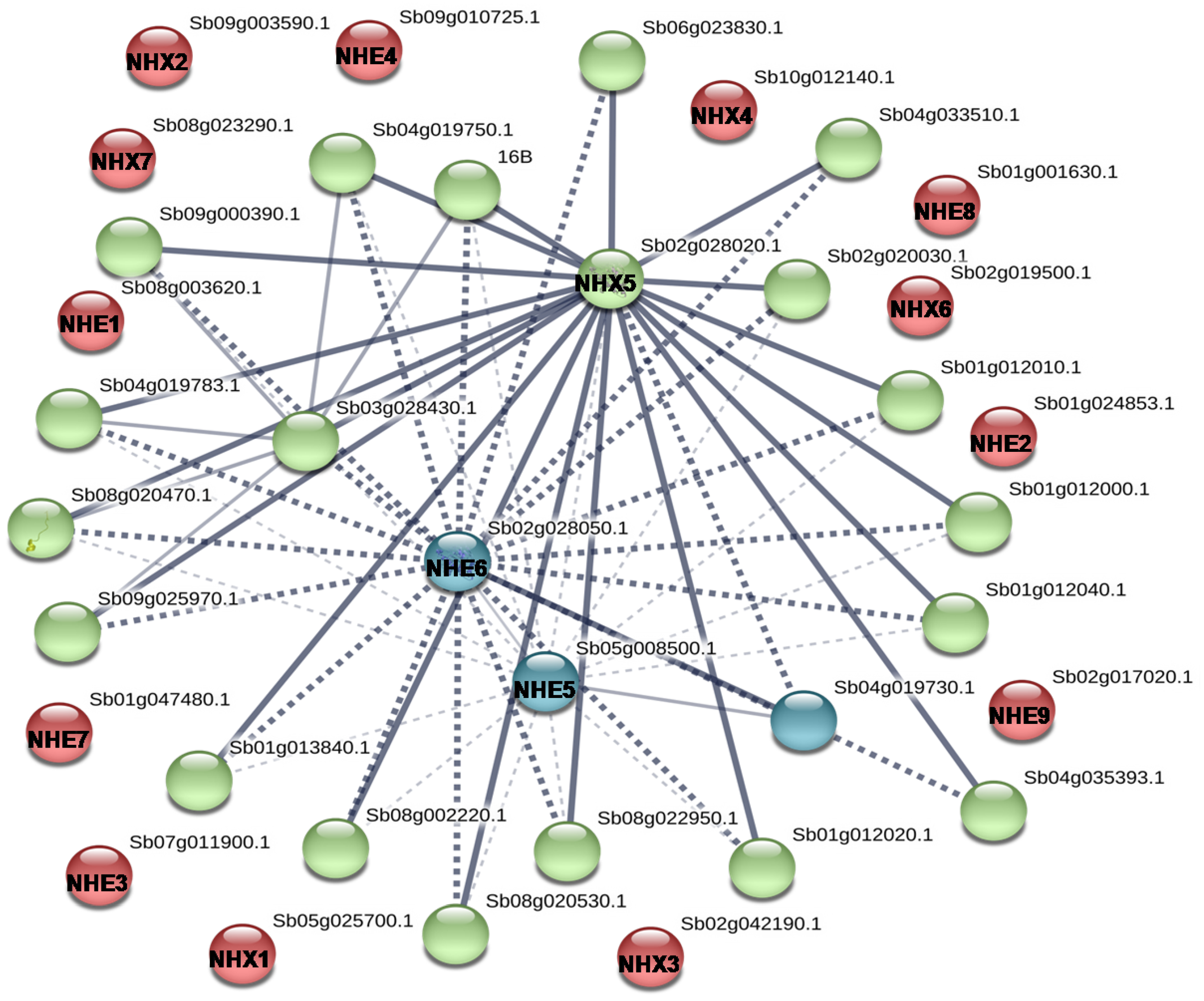
3.3. Protein–Protein Interaction Analysis Prediction
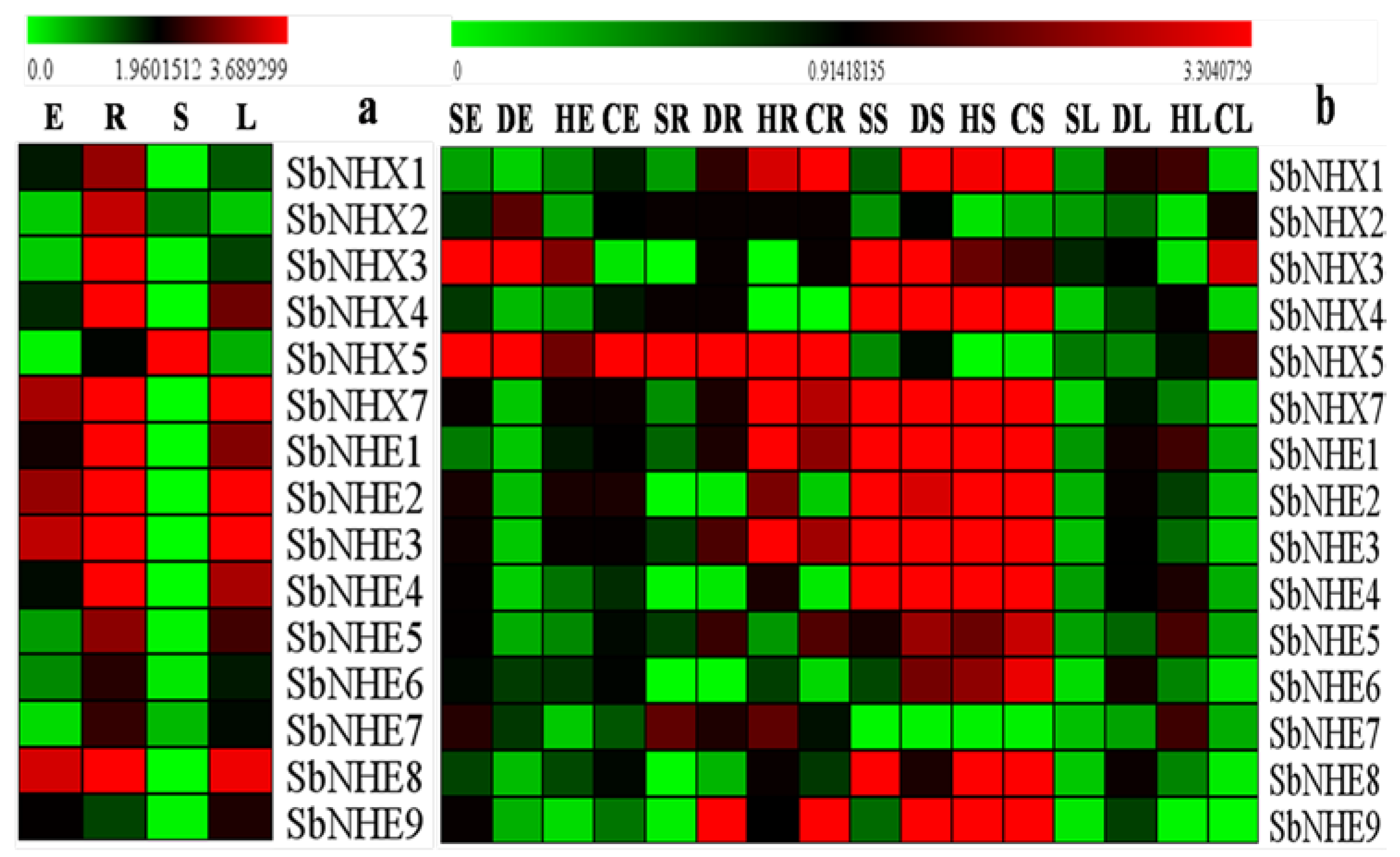
3.4. Expression Analysis of SbNHX/NHE Transporters in Different Tissues Treated with Diverse Abiotic Stresses
4. Discussion
4.1. Identification and Structural Analysis of SbNHX/NHE Genes
4.2. Motif Identification, Promoter Analysis, Phylogenetic Tree, and Divergence
4.3. Gene Expressions in Different Tissues under Abiotic Stresses
4.4. Protein–Protein Interaction Predictions
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [PubMed]
- Bohnert, H.J.; Nelsen, D.E.; Jensen, R.G. Adaptation to environmental stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Chen, Q.J.; Chai, M.F.; Lu, P.L.; Su, Z.; Qin, Z.X.; Chen, J.; Wang, X.C. AtNHX8, a member of the monovalent cation: Proton antiporter-1 family in Arabidopsis thaliana, encodes a putative Li+/H+ antiporter. Plant J. 2007, 49, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Mojca, B.; Tanja, B.; Ljerka, L.; Nada, K. A comparative genomic analysis of calcium and proton signaling/homeostasis in Aspergillus species. Fungal Genet. Biol. 2009, 46, S93–S104. [Google Scholar]
- Amtmann, A.; Sanders, D. Mechanisms of Na+ uptake by plant cells. Adv. Bot. Res. 1999, 29, 76–112. [Google Scholar]
- Schachtman, D.; Liu, W. Molecular pieces to the puzzle of the interaction between potassium and sodium uptake in plants. Trends Plant. Sci. 1999, 4, 281–287. [Google Scholar] [CrossRef]
- Maser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; et al. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef] [PubMed]
- Brett, C.L.; Donowitz, M.; Rao, R.; Christopher, L. Evolutionary origins of eukaryotic Na+/H+ exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223–C239. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Feng, M.; Muend, S.; Rao, R. A human Na+/H+ antiporter sharing evolutionary origins with bacterial NhaA may be a candidate gene for essential hypertension. Proc. Nat. Acad. Sci. USA 2007, 104, 18677–18681. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rosales, P.M.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/protonantiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Waditee, R.; Hibino, T.; Nakamura, T.; Incharoensakdi, A.; Takabe, T. Overexpression of a Na+/H+ antiporter confers salt tolerance on a freshwater cyanobacterium, making it capable of growth in sea water. Proc. Nat. Acad. Sci. USA 2002, 99, 4109–4114. [Google Scholar] [CrossRef] [PubMed]
- Putney, L.K.; Denker, S.P.; Barber, D.L. The changing face of the Na+/H+ exchanger, NHE1: Structure, regulation, and cellular actions. Ann. Rev. Pharmacol. Toxicol. 2002, 42, 527–552. [Google Scholar] [CrossRef] [PubMed]
- Leidi, E.O.; Barragan, V.; Rubio, L.; El-Hamdaoui, A.; Ruiz, M.T.; Cubero, B.; Fernández, J.A.; Bressan, R.A.; Hasegawa, P.M.; Quintero, F.J.; et al. The AtNHX1 exchanger mediates potassium compartmentation in vacuoles of transgenic tomato. Plant J. 2010, 61, 495–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef] [PubMed]
- Gaxiola, R.A.; Rao, R.; Sherman, A.; Grisafi, P.; Alper, S.L.; Fink, G.R. The Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can function in cation detoxification in yeast. Proc. Natl. Acad. Sci. USA 1999, 96, 1480–1485. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, S.; Quintero, F.J.; Cubero, B.; Ruiz, M.T.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Differential expression and function of Arabidopsis thaliana NHX Na+/H+ antiporters in the salt stress response. Plant J. 2002, 30, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Ohto, M.A.; Esumi, T.; Tajima, H.; Zhu, Z.; Cagnac, O.; Belmonte, M.; Peleg, Z.; Yamaguchi, T.; Blumwald, E. The Arabidopsis intracellular Na+/H+ antiporters NHX5 and NHX6 are endosome associated and necessary for plant growth and development. Plant Cell 2011, 23, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Tajima, H.; Liang, Y.C.; Ohto, M.A.; Ushijima, K.; Nakano, R.; Esumi, T.; Coku, A.; Belmonte, M.; Blumwald, E. The Arabidopsis Na+/H+antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction. Plant Cell 2011, 223, 3482–3492. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.X.; Blumwald, E. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol. 2001, 19, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Barragan, V.; Leidi, E.O.; Andres, Z.; Rubio, L.; Luca, A.D.; Fernandez, J.A.; Cubero, B.; Pardo, J.M. Ion exchangers NHX1 and NHX2 mediate active potassium uptake into vacuoles to regulate cell turgor and stomatal function in Arabidopsis. Plant Cell 2012, 24, 1127–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.X.; Li, J.; Wu, X.D.; Liu, Q.; Wang, Z.K.; Liu, S.S.; Li, S.N.; Ma, Y.L.; Sun, J.; Zhao, L.; et al. Ectopic expression of Arabidopsis thaliana Na+(K+)/H+ antiporter gene, AtNHX5, enhances soybean salt tolerance. Genet. Mol. Res. 2016, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Counillon, L.; Pouyssegur, J. The expanding family of eukaryotic Na+/H+ exchangers. J. Biol. Chem. 2000, 275, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J.; Grinstein, S. Na+/H+ exchangers in mammalian cells. J. Biol. Chem. 1997, 272, 22373–22376. [Google Scholar] [CrossRef] [PubMed]
- Bobulescua, I.A.; Soleb, F.D.; Moe, O.W. Na+/H+ exchangers: Physiology and link to hypertension and organ ischemia. Curr. Opin. Nephrol. Hypertense 2005, 14, 485–494. [Google Scholar] [CrossRef]
- Wakabayashi, S.; Pang, T.; Su, X.; Shigekawa, M. A novel topology model of the human Na+/H+ exchanger isoform. J. Biol. Chem. 2000, 275, 7942–7949. [Google Scholar] [CrossRef] [PubMed]
- Biemesderfer, D.; Reilly, R.F.; Exner, M.; Igarashi, P.; Aronson, P.S. Immunocytochemical characterization of Na+-H+ exchanger isoform NHE-1 in rabbit kidney. Am. J. Physiol. Ren. Fluid Electrolyte Physiol. 1992, 263, F833–F840. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, P.H.; Kumar, A.S.; Sivan, P.; Katam, R.; Suravajhala, P.; Rao, K.S.; Varshney, R.K.; Kishor, P.B.K. Overexpression of a plasma membrane Na+/H+-antiporter-like protein (SbNHXLP) confers salt tolerance and improves fruit yield in tomato by maintaining ion homeostasis. Front. Plant Sci. 2017, 7, 2027. [Google Scholar] [CrossRef] [PubMed]
- Anshita, G.; Gohar, T.; Dinesh, P.; Sanjay, G.; Anil, K. Genome-wide comparative in silico analysis of calcium transporters of rice and sorghum. Genom. Proteom. Bioinform. 2011, 9, 138–150. [Google Scholar]
- Amrutha, R.N.; Sekhar, P.N.; Varshney, R.K.; Kishor, P.B.K. Genome-wide analysis and identification of genes related to potassium transporter families in rice (Oryza sativa L.). Plant Sci. 2007, 172, 708–721. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Chen, Y.; Li, R.; Wang, H.; Wei, J. Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.). Mol. Biol. Rep. 2012, 39, 8465–8473. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Chang, E.; Li, Y.; Sun, P.; Hu, J.; Zhang, J. Expression and integrated network analyses revealed functional divergence of NHX-type Na+/H+ exchanger genes in poplar. Sci. Rep. 2017, 7, 2607. [Google Scholar] [CrossRef] [PubMed]
- Burge, C.B.; Karlin, S. Finding the genes in genomic DNA. Curr. Opin. Struct. Biol. 1998, 8, 346–354. [Google Scholar] [CrossRef]
- Guo, A.Y.; Zhu, Q.H.; Chen, X.; Luo, J.C.; Chuan, Y.I. GSDS a gene structure display server. Yi Chuan 2007, 29, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Moller, S.; Croning, M.D.R.; Apweller, R. Evaluation of methods for the prediction of membrane spanning regions. Bioinformatics 2001, 17, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.C.J.; Nekai, A.C.K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence-and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Gribskov, M. Combining evidence using p-values: Application to sequence homology searches. Bioinformatics 1998, 14, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Copley, R.R.; Schmidt, S.; Ciccarelli, F.D.; Doerks, T.; Schultz, J.; Ponting, C.P.; Bork, P. SMART 4.0: Towards genomic data integration. Nucleic Acids Res. 2004, 32D, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Cavalcanti, A.; Chen, F.C.; Bouman, P.; Li, W.H. Extent of gene duplication in the genomes of Drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhang, X.; Yue, J.X.; Tian, D.; Chen, J.Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol. Genet. Genom. 2008, 280, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef] [PubMed]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database. NucleicAcids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Lescot, M.P.; Dehais, G.; Thijs, K.; Marchal, Y.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.S.; Reddy, D.S.; Sivasakthi, K.; Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Evaluation of sorghum (Sorghum bicolor (L.)) reference genes in various tissues and under abiotic stress conditions for quantitative real-time PCR data normalization. Front. Plant Sci. 2016, 7, 529. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Munns, M.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Ann. Rev. Plant. Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, J.; Grinstein, S. Diversity of the mammalian Na+/H+exchanger SLC9 gene family. Pflugers Arch. 2004, 447, 549–565. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Blumwald, E. The ins and outs of intracellular ion homeostasis: NHX-type cation/H+ transporters. Curr. Opin. Plant Biol. 2014, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Zhang, C.; Hicks, G.R.; Raikhel, N.V. Plant vacuole morphology and vacuolar trafficking. Front. Plant Sci. 2013, 5, 476. [Google Scholar] [CrossRef] [PubMed]
- Moffat, A.S. Plant genetics. Finding new ways to protect drought-stricken plants. Science 2002, 296, 1226–1229. [Google Scholar] [CrossRef] [PubMed]
- Brett, C.L.; Tukaye, D.N.; Mukherjee, S.; Rao, R. The yeast endosomal Na+K+/H+ exchanger NHX1 regulates cellular pH to control vesicle trafficking. Mol. Biol. Cell 2005, 16, 1396–1405. [Google Scholar] [CrossRef] [PubMed]
- Chanroj, S.; Wang, G.; Venema, K.; Zhang, M.W.; Delwiche, C.F.; Sze, H. Conserved and diversified gene families of monovalent Na+/H+ antiporters from algae to flowering plants. Front. Plant. Sci. 2012, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, J.; Aronson, P. Amiloride inhibition of the Na+/H+ exchanger in renal microvillus membrane vesicles. Am. J. Physiol. Ren. Physiol. 1981, 241, F374–F379. [Google Scholar] [CrossRef] [PubMed]
- Blumwald, E.; Poole, R.J. Salt tolerance in suspension cultures of sugar beet induction of Na+/H+ antiport activity at the tonoplast by growth in salt. Plant Physiol. 1987, 83, 884–887. [Google Scholar] [CrossRef] [PubMed]
- Plesch, G.; Ehrhardt, T.; Mueller-Roeber, B. Involvement of TAAAG elements suggests a role for Dof transcription factors in guard cell-specific gene expression. Plant J. 2001, 28, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Deniaud, E.; Baguet, J.; Chalard, R.; Blanquier, B.; Brinza, L.; Meunier, J.; Michallet, M.C.; Laugraud, A.; Ah-Soon, C.; Wierinckx, A.; et al. Overexpression of transcription factor Sp1 leads to gene expression perturbations and cell cycle inhibition. PLoS ONE. 2009, 4, e7035. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, W.; Hu, X.; Liu, H.; Lin, Y. W-box and G-box elements play important roles in early senescence of rice flag leaf. Sci. Rep. 2016, 6, 20881. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Nagaraju, M.; Reddy, P.S.; Kumar, A.S.; Srivastava, R.K.; Kishor, P.B.K.; Rao, D.M. Genome-wide scanning and characterization of Sorghum bicolor L. heat shock transcription factors. Curr. Genom. 2015, 16, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Ashnest, J.R.; Huynh, D.L.; Dragwidge, J.M.; Ford, B.A.; Gendall, A.R. Arabidopsis intracellular NHX-type sodium-proton antiporters are required for seed storage protein processing. Plant. Cell. Physiol. 2015, 56, 2220–2233. [Google Scholar] [CrossRef] [PubMed]
- Katiyar-Agarwal, S.; Zhu, J.; Kim, K.; Agarwal, M.; Fu, X.; Huang, A.; Zhu, J. The plasma membrane Na+/H+ antiporter SOS1 interacts with RCD1 and functions in oxidative stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 18816–18821. [Google Scholar] [CrossRef] [PubMed]
- Quintero, F.J.; Martinez-Atienza, J.; Villalta, I.; Jiang, X.; Kim, W.Y.; Ali, Z.; Fujii, H.; Mendoza, I.; Yun, D.-J.; Zhu, K.-K.; et al. Activation of the plasma membrane Na/H antiporter salt-overly-sensitive 1 (SOS1) by phosphorylation of an auto-inhibitory C-terminal domain. Proc. Natl. Acad. Sci. USA 2011, 108, 2611–2616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Wang, G.; Ji, J.; Tian, X.; Gao, H.; Zhao, Q.; Li, J.; Wang, Y. Hydrophilic C terminus of Salicornia europaea vacuolar Na+/H+ antiporter is necessary for its function. J. Genet. 2014, 93, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Weinl, S.; Kudla, J. The CBL-CIPK Ca2+-decoding signaling network: Function and perspectives. New Phytol. 2009, 184, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, J.K. A calcium sensor homolog required for plant salt tolerance. Science 1998, 280, 1943–1945. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, M.; Liu, J.; Halfter, U.; Kim, C.S.; Shi, W.; Zhu, J.K. SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding. Plant Cell 2000, 12, 1667–1678. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.C.; Auge, R.M.; Dong, C.; Cheng, Z.M.M. Increased salt tolerance with overexpression of Na+/H+antiporter 1 genes: A meta-analysis. Plant. Biotechnol. J. 2017, 15, 162–173. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ACC Number | Gene Name | No. ofAa | Chr | Domain | DBD | pI/MW | GRAVY | TMHMM | No. of Exons | Localization | Instability INDEX |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sb05g025700.1 | SbNHX1 | 386 | 5 | NHX | 23–283 | 9.28/42,070.0 | 0.445 | 7 | 8 | PM | 40.26 |
| Sb09g003590.1 | SbNHX2 | 823 | 9 | NHX | 377–719 | 9.56/89,834.3 | 0.144 | 8 | 18 | Chlo | 48.13 |
| Sb02g042190.1 | SbNHX3 | 696 | 2 | NHX | 405–560 | 9.42/76,720.9 | 0.151 | 4 | 16 | PM | 44.18 |
| Sb10g012140.1 | SbNHX4 | 631 | 10 | NHX | 192–516 | 10.19/70,077.0 | −0.009 | 8 | 13 | PM | 42.92 |
| Sb02g028020.1 | SbNHX5 | 1466 | 2 | NHX | 459–577 | 6.74/164,308.4 | −0.483 | 2 | 18 | Chlo | 45.69 |
| Sb02g019500.1 | SbNHX6 | 434 | 2 | NHX | 191–346 | 6.41/48,771.7 | −0.400 | 0 | 12 | ER | 47.89 |
| Sb08g023290.1 | SbNHX7 | 178 | 8 | NHX | 14–178 | 6.27/18,648.0 | 0.688 | 5 | 5 | Cyto | 24.22 |
| Sb08g003620.1 | SbNHE1 | 597 | 8 | Ovate | 319–375 | 5.25/62,189.4 | −0.357 | 0 | 5 | Chlo | 46.11 |
| Unknown | SbNHE2 | 1414 | 3 | RVT | 595–752 | 6.19/163,691.7 | −0.361 | 0 | 2 | Cyto | 48.12 |
| Unknown | SbNHE3 | 726 | 3 | RVE | 258–367 | 9.21/84,058.5 | −0.587 | 0 | 5 | Nucl | 57.40 |
| Unknown | SbNHE4 | 1826 | 10 | Transposase | 686–867 | 6.75/203,438.2 | −0.738 | 0 | 14 | Nucl | 58.73 |
| Sb05g008500.1 | SbNHE5 | 955 | 5 | PKinase | 496–709 | 8.71/101,934.8 | −0.006 | 2 | 3 | PM | 49.28 |
| Sb02g028020.1 | SbNHE6 | 699 | 2 | HSP90 | 185–695 | 4.97/80,238.0 | −0.609 | 0 | 3 | Nucl | 36.89 |
| Sb01g047480.1 | SbNHE7 | 637 | 1 | VWA | 169–355 | 6.07/67,727.9 | −0.285 | 0 | 1 | Chlo | 50.99 |
| Sb01g001630.1 | SbNHE8 | 1298 | 1 | DUF | 48–362 | 6.94/145,110.4 | −0.335 | 0 | 13 | Cyto | 39.59 |
| Sb02g017020.1 | SbNHE9 | 840 | 2 | RVT1 | 65–214 | 9.10/95,836.4 | −0.320 | 0 | 3 | Mito | 36.69 |
| Gene 1 | Gene 2 | dN | dS | dN/dS |
|---|---|---|---|---|
| SbNHX5 | SbNHE6 | 0.0047 | 0.3627 | 0.0130 |
| SbNHE1 | SbNHE7 | 1.7237 | 60.2714 | 0.0286 |
| SbNHE4 | SbNHE9 | 1.3993 | 50.2271 | 0.0279 |
| Gene | Hormone Responsive | Stress Responsive | Others | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ABRE | TGAAACGAC | GARETCTGTTG | EREATTTCAAA | Me-Ja | TCACAGAAAAGGA | DRE | HSE | LTR | GT1 | MYB | Box | TC rich | Sp1 motif | GARETCTGTTG | KST1 | SKN1 | Box4 | Box I | G Box | II Box | O2 site | Circadian | |
| SbNHX1 | 10 | 1 | 0 | 0 | 4 | 1 | 2 | 3 | 0 | 2 | 2 | 1 | 0 | 18 | 0 | 4 | 2 | 1 | 1 | 12 | 0 | 1 | 2 |
| SbNHX2 | 2 | 3 | 1 | 1 | 7 | 2 | 0 | 3 | 0 | 5 | 2 | 2 | 2 | 3 | 1 | 8 | 6 | 3 | 4 | 7 | 0 | 1 | 1 |
| SbNHX3 | 7 | 0 | 1 | 1 | 3 | 0 | 0 | 1 | 0 | 7 | 3 | 2 | 1 | 1 | 1 | 13 | 7 | 3 | 3 | 18 | 0 | 2 | 1 |
| SbNHX4 | 3 | 1 | 0 | 0 | 4 | 0 | 1 | 2 | 0 | 4 | 4 | 1 | 4 | 1 | 0 | 8 | 6 | 3 | 1 | 8 | 2 | 0 | 2 |
| SbNHX5 | 0 | 0 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 2 | 1 | 1 | 3 | 0 | 1 | 4 | 6 | 0 | 2 | 5 | 0 | 1 | 0 |
| SbNHX6 | 0 | 1 | 5 | 2 | 2 | 1 | 0 | 1 | 0 | 4 | 5 | 2 | 5 | 2 | 5 | 13 | 3 | 2 | 4 | 3 | 1 | 2 | 3 |
| SbNHX7 | 0 | 0 | 0 | 0 | 4 | 3 | 1 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 6 | 2 | 5 | 2 | 8 | 1 | 1 | 3 |
| SbNHE1 | 4 | 0 | 1 | 1 | 4 | 1 | 3 | 1 | 2 | 8 | 0 | 0 | 2 | 2 | 1 | 4 | 5 | 1 | 1 | 9 | 0 | 1 | 1 |
| SbNHE2 | 0 | 0 | 0 | 0 | 1 | 0 | 4 | 0 | 0 | 2 | 1 | 0 | 1 | 9 | 0 | 6 | 0 | 2 | 1 | 6 | 4 | 1 | 3 |
| SbNHE3 | 1 | 1 | 0 | 0 | 1 | 3 | 0 | 0 | 1 | 4 | 6 | 0 | 0 | 2 | 0 | 5 | 7 | 0 | 0 | 4 | 3 | 0 | 3 |
| SbNHE4 | 1 | 0 | 0 | 0 | 2 | 1 | 2 | 0 | 0 | 1 | 1 | 0 | 2 | 3 | 0 | 7 | 7 | 2 | 0 | 2 | 1 | 2 | 4 |
| SbNHE5 | 1 | 0 | 0 | 0 | 5 | 0 | 3 | 5 | 0 | 4 | 3 | 3 | 1 | 4 | 0 | 8 | 2 | 2 | 0 | 3 | 1 | 1 | 2 |
| SbNHE6 | 1 | 2 | 1 | 0 | 0 | 2 | 0 | 3 | 0 | 4 | 3 | 1 | 2 | 1 | 1 | 7 | 10 | 3 | 1 | 0 | 4 | 3 | 4 |
| SbNHE7 | 0 | 0 | 2 | 0 | 2 | 3 | 0 | 2 | 1 | 2 | 2 | 1 | 2 | 3 | 2 | 6 | 3 | 1 | 3 | 5 | 0 | 1 | 4 |
| SbNHE8 | 2 | 2 | 0 | 0 | 2 | 1 | 1 | 1 | 1 | 4 | 2 | 0 | 0 | 6 | 0 | 3 | 4 | 1 | 0 | 3 | 4 | 1 | 1 |
| SbNHE9 | 5 | 1 | 1 | 0 | 12 | 0 | 1 | 1 | 0 | 2 | 4 | 2 | 0 | 5 | 1 | 4 | 5 | 0 | 0 | 10 | 2 | 1 | 8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hima Kumari, P.; Anil Kumar, S.; Ramesh, K.; Sudhakar Reddy, P.; Nagaraju, M.; Bhanu Prakash, A.; Shah, T.; Henderson, A.; Srivastava, R.K.; Rajasheker, G.; et al. Genome-Wide Identification and Analysis of Arabidopsis Sodium Proton Antiporter (NHX) and Human Sodium Proton Exchanger (NHE) Homologs in Sorghum bicolor. Genes 2018, 9, 236. https://doi.org/10.3390/genes9050236
Hima Kumari P, Anil Kumar S, Ramesh K, Sudhakar Reddy P, Nagaraju M, Bhanu Prakash A, Shah T, Henderson A, Srivastava RK, Rajasheker G, et al. Genome-Wide Identification and Analysis of Arabidopsis Sodium Proton Antiporter (NHX) and Human Sodium Proton Exchanger (NHE) Homologs in Sorghum bicolor. Genes. 2018; 9(5):236. https://doi.org/10.3390/genes9050236
Chicago/Turabian StyleHima Kumari, P., S. Anil Kumar, Katam Ramesh, Palakolanu Sudhakar Reddy, M. Nagaraju, A. Bhanu Prakash, Trushar Shah, Ashley Henderson, Rakesh K. Srivastava, G. Rajasheker, and et al. 2018. "Genome-Wide Identification and Analysis of Arabidopsis Sodium Proton Antiporter (NHX) and Human Sodium Proton Exchanger (NHE) Homologs in Sorghum bicolor" Genes 9, no. 5: 236. https://doi.org/10.3390/genes9050236