Over a Thousand Years of Evolutionary History of Domestic Geese from Russian Archaeological Sites, Analysed Using Ancient DNA

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
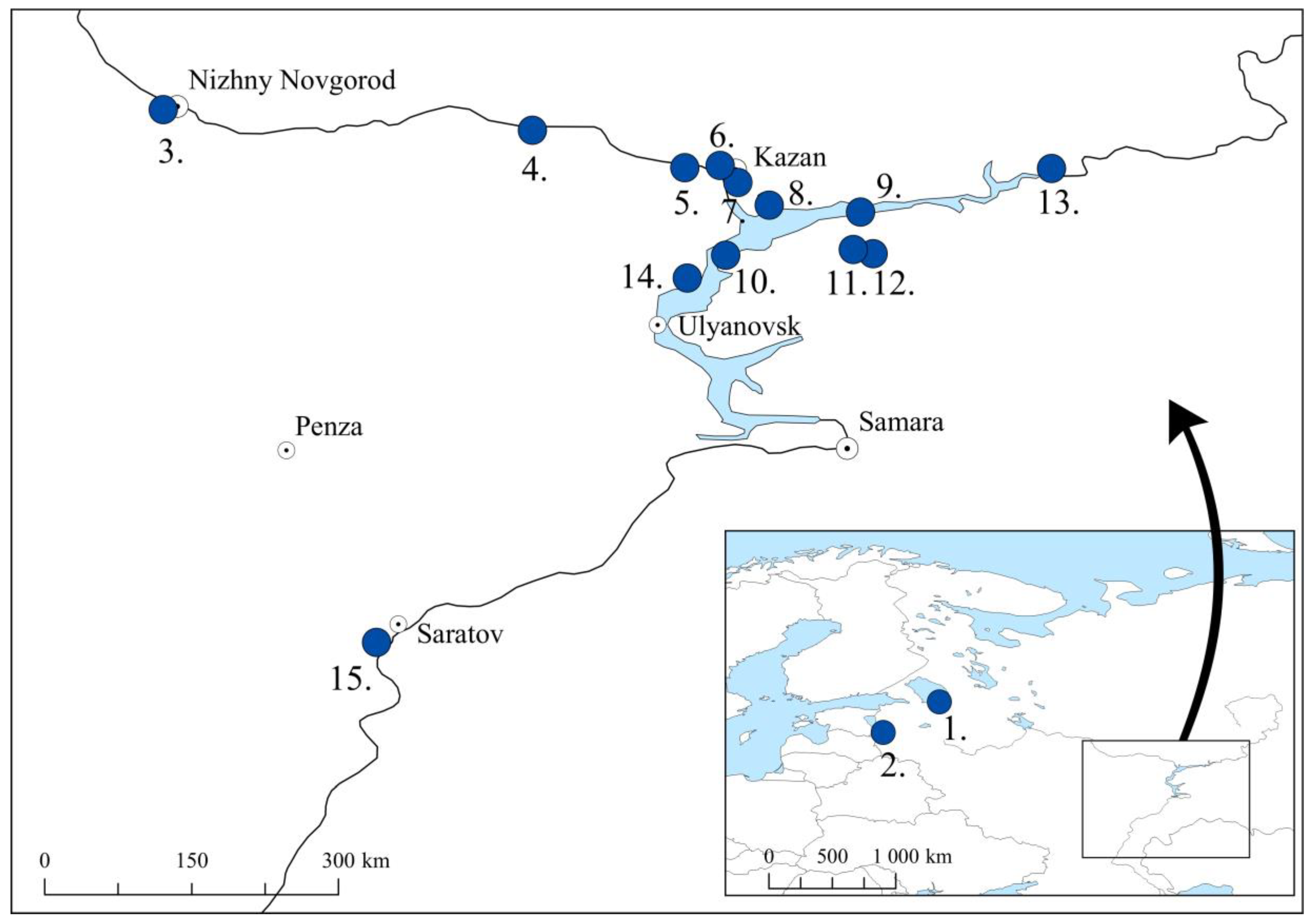
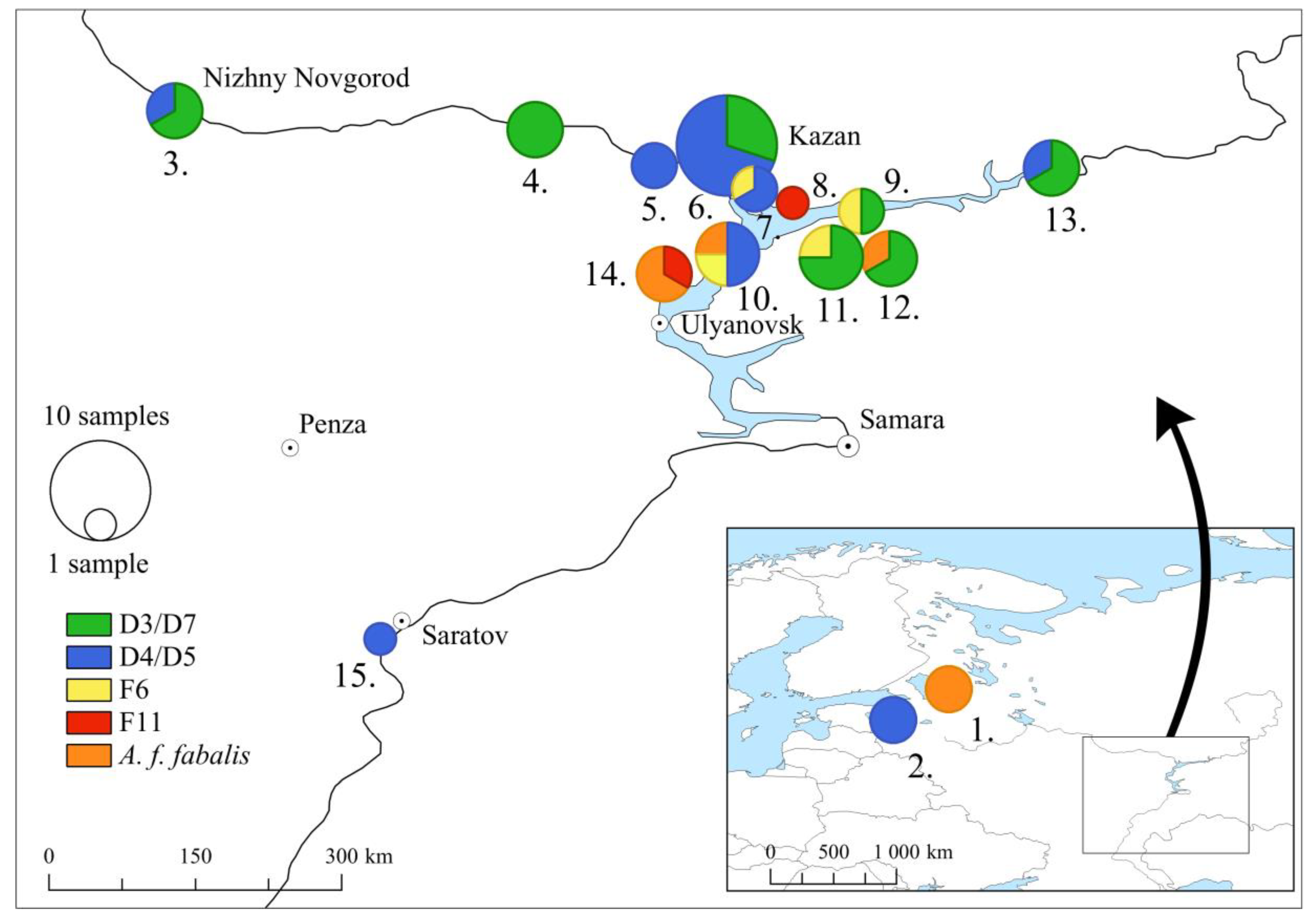
2.1. Sample Material
2.2. DNA Extraction and Amplification
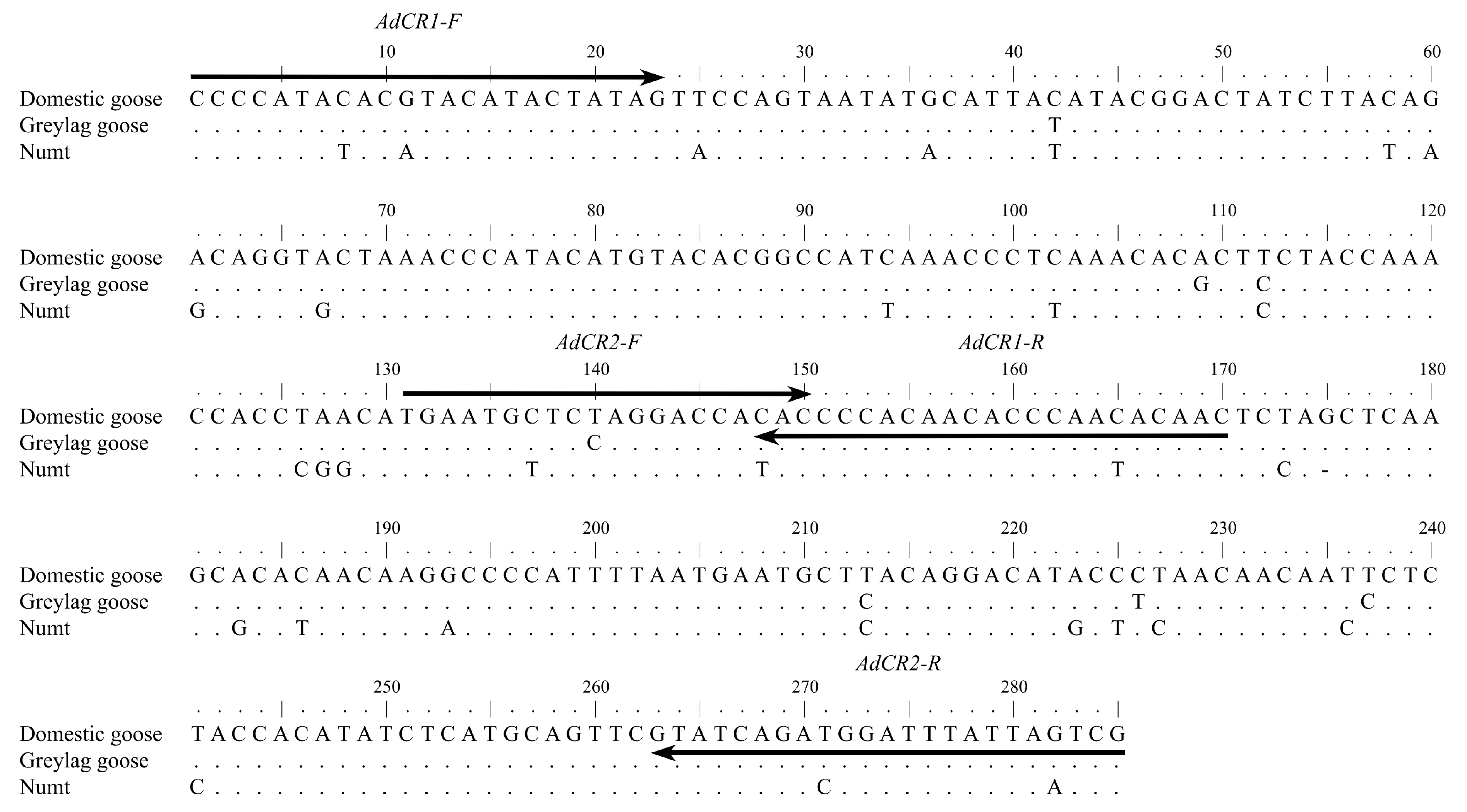
2.3. Authentication
2.4. Sequence Analyses
3. Results
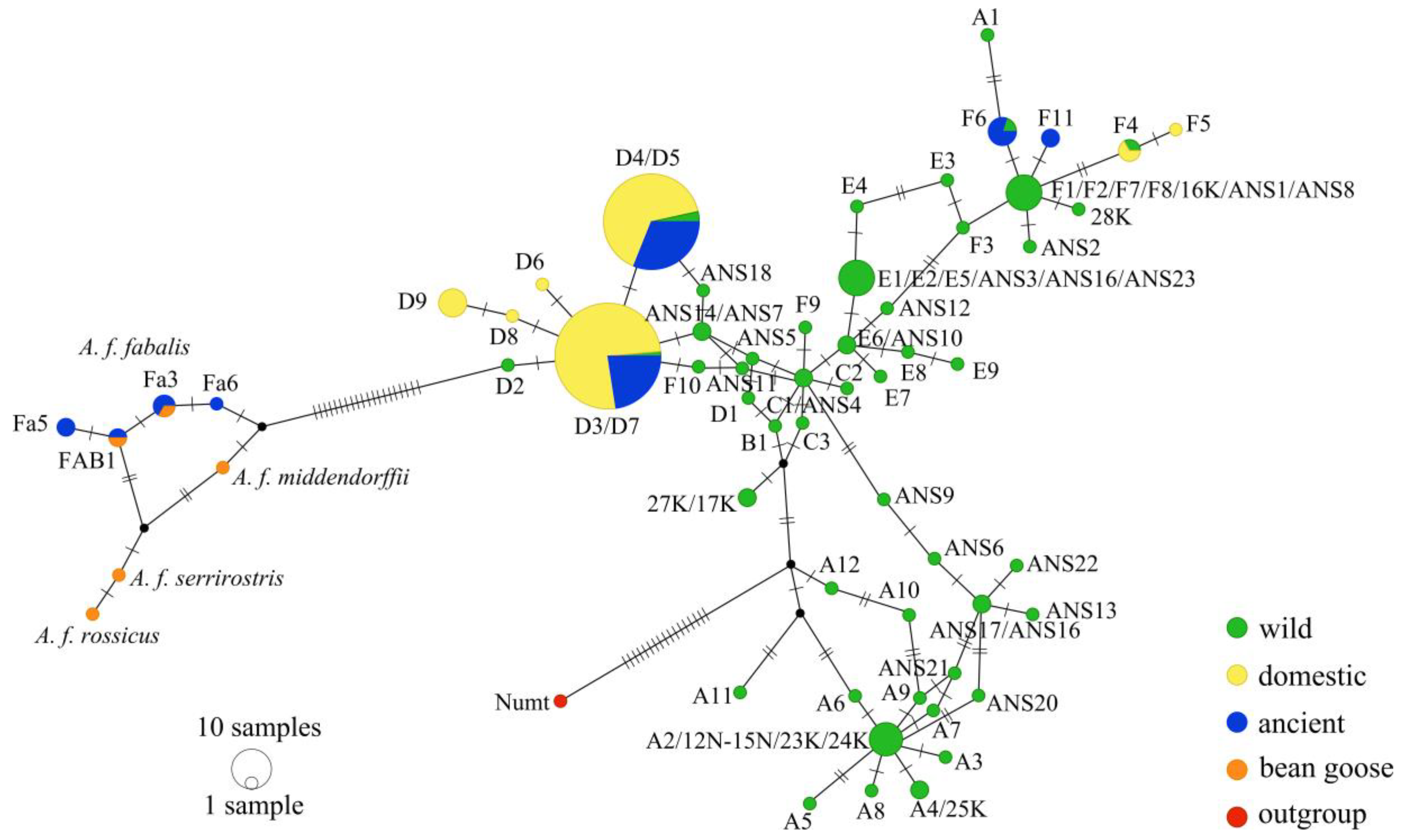
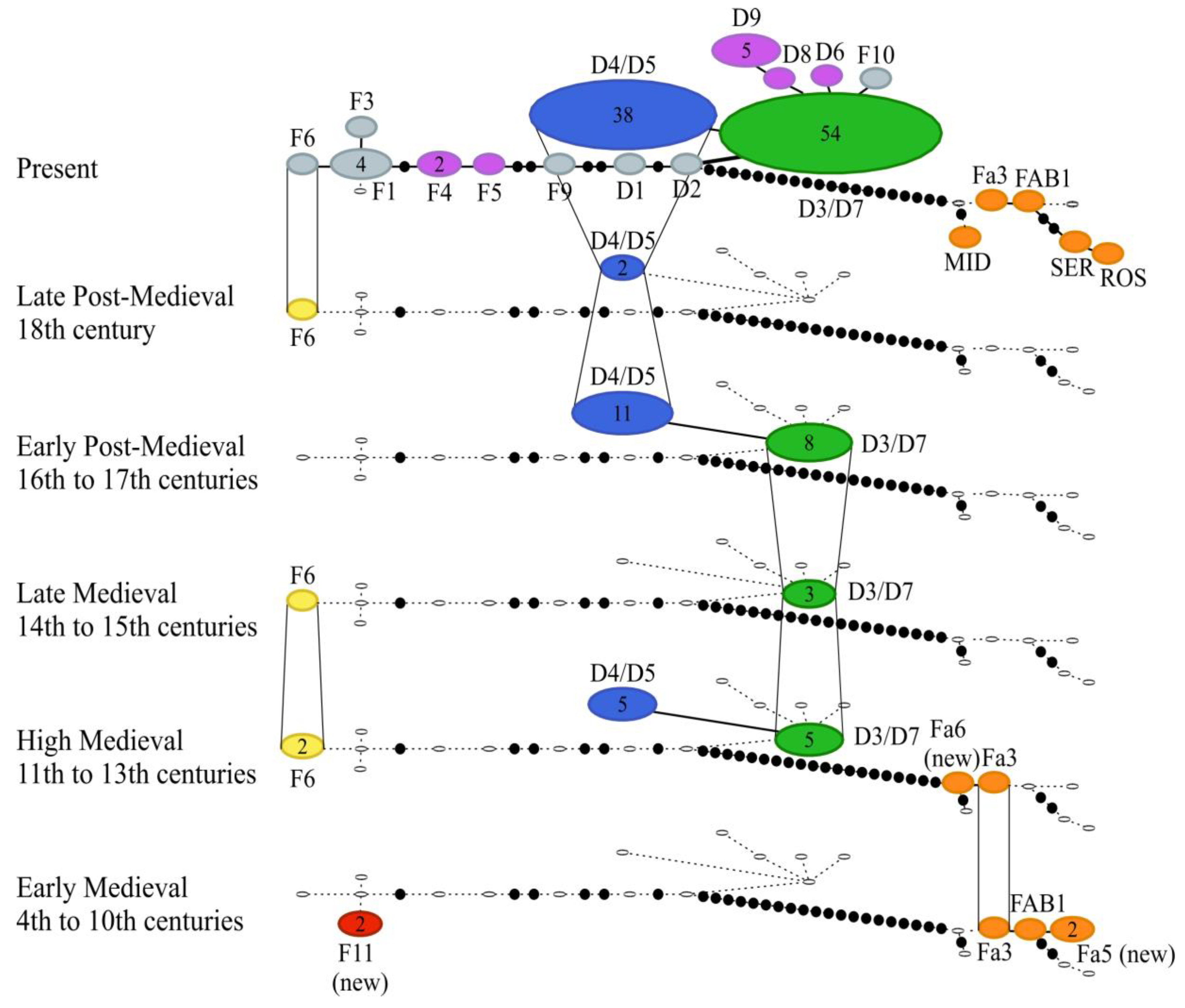
3.1. Mitochondrial DNA Haplotypes
3.2. Genetic Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Albarella, U. Alternate fortunes? The role of domestic ducks and geese from Roman to Medieval times in Britain. In Documenta Archaeobiologiae 3. Feathers, Grit and Symbolism; Grupe, G., Peters, J., Eds.; Verlag Marie Leidorf: Rahden/Westphalia, Germany, 2005; pp. 249–258. [Google Scholar]
- Zeuner, F.E. A History of Domesticated Animals; Hutchinson: London, UK, 1963. [Google Scholar]
- Shi, X.W.; Wang, J.W.; Zeng, F.T.; Qiu, X.P. Mitochondrial DNA cleavage patterns distinguish independent origin of Chinese domestic geese and Western domestic geese. Biochem. Genet. 2006, 44, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.M.; Way, T.D.; Chang, Y.C.; Yen, N.T.; Hu, C.L.; Nien, P.C.; Jea, Y.S.; Chen, L.R.; Kao, J.Y. The origin of the white Roman goose. Biochem. Genet. 2010, 48, 938–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kear, J. Man and Wildfowl; Poyser: London, UK, 1990. [Google Scholar]
- Larson, G.; Fuller, D.Q. The evolution of animal domestication. Ann. Rev. Ecol. Evol. Syst. 2014, 45, 115–136. [Google Scholar] [CrossRef]
- Crawford, R.D. Goose. In Evolution of Domesticated Animals; Mason, I.L., Ed.; Longman: London, UK, 1984; pp. 345–349. [Google Scholar]
- Li, H.F.; Zhu, W.Q.; Chen, K.W.; Xu, W.J.; Song, W. Two maternal origins of Chinese domestic goose. Poult. Sci. 2011, 90, 2705–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhang, S.; He, D.Q.; Chen, S.Y.; Duan, Z.Y.; Yao, Y.G.; Liu, Y.P. Matrilineal genetic structure of domestic geese. J. Poult. Sci. 2014, 51, 130–137. [Google Scholar] [CrossRef]
- Ren, T.; Liang, S.; Zhao, A.; He, K. Analysis of the complete mitochondrial genome of the Zhedong White goose and characterization of NUMTs: Reveal domestication history of goose in China and Euro. Gene 2016, 577, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, M.E.; Ruokonen, M.; Alexander, M.; Aspi, J.; Pyhäjärvi, T.; Searle, J.B. Relationship between wild greylag and European domestic geese based on mitochondrial DNA. Anim. Genet. 2015, 46, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyrberg, T. The archaeological record of domesticated and tamed birds in Sweden. Acta Zool. Cracov. 2002, 45, 215–231. [Google Scholar]
- MacHugh, D.E.; Larson, G.; Orlando, L. Taming the past: Ancient DNA and the study of animal domestication. Annu. Rev. Anim. Biosci. 2017, 5, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Barnes, I.; Young, J.P.W.; Dobney, K.M. DNA-based identification of goose species from two archaeological sites in Lincolnshire. J. Archaeol. Sci. 2000, 27, 91–100. [Google Scholar] [CrossRef]
- Barnes, I.; Dobney, K.M.; Young, J.P.W. The molecular palaeoecology of geese: Identification of archaeological goose remains using ancient DNA analysis. Int. J. Osteoarchaeol. 1998, 8, 280–287. [Google Scholar] [CrossRef]
- Paxinos, E.E.; James, H.F.; Olson, S.L.; Sorenson, M.D.; Jackson, J.; Fleischer, R.C. mtDNA from fossils reveals a radiation of Hawaiian geese recently derived from the Canada goose (Branta canadensis). Proc. Natl. Acad. Sci. USA 2002, 99, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.J.; Crockford, S.J.; Johnson, J.W.; Malhi, R.S.; Kemp, B.M. Genetic and archaeological evidence for a former breeding population of Aleutian cackling goose (Branta hutchinsii leucopareia) on Adak Island, central Aleutians, Alaska. Can. J. Zool. 2011, 89, 732–743. [Google Scholar] [CrossRef]
- Paxinos, E.E.; James, H.F.; Olson, S.L.; Ballou, J.D.; Leonard, J.A.; Fleischer, R.C. Prehistoric decline of genetic diversity in the nene. Science 2002, 296, 1827. [Google Scholar] [CrossRef] [PubMed]
- Galimova, D.N.; Askeyev, I.V.; Askeyev, O.V. Bird remains from 5th–17th century AD archaeological sites in the Middle Volga Region of Russia. Int. J. Osteoarchaeol. 2014, 24, 347–357. [Google Scholar] [CrossRef]
- Bacher, A. Vergleichend morphologische Untersuchungen an Einzelknochen des postkranialen Skeletts in Mitteleuropa vorkommender Schwäne und Gänse. Doctoral Dissertation, Institut für Palaeoanatomie, Domestikationsforschung und Geschichte der Tiermedizin der Universität München, München, Germany, 1967. (In German). [Google Scholar]
- Umanskaya, A.S. Domestic Birds from Archaeological Sites of Ukraine; Natural Environment and Fauna of the Past: Kiev, Ukraine, 1972. (In Russian) [Google Scholar]
- Serjeantson, D. Birds; Cambridge Manuals in Archaeology; Cambridge University Press: New York, NY, USA, 2009. [Google Scholar]
- Ruokonen, M.; Kvist, L.; Lumme, J. Close relatedness between mitochondrial DNA from seven Anser goose species. J. Evol. Biol. 2000, 13, 532–540. [Google Scholar] [CrossRef]
- Askeyev, I.V.; Galimova, D.N.; Askeyev, O.V. Birds of the Middle Volga Region during the V–XVIII centuries AD (according to archaeological excavations). Volga River Reg. Archaeol. (Zhurnal Povolz. Arkheologiya) 2013, 3, 116–144. (In Russian) [Google Scholar] [CrossRef]
- Shaymuratova (Galimova), D.N.; Askeyev, I.V.; Askeyev, O.V. The Studies of Archaeological Bird Remains of Medieval Staraya Ladoga: New Results and Interpretation. In Proceedings of the Monographs of the Archaeological Society of Finland 7, Twin Conference X Nordic Stratigrafimötet and XI Applications of the Scientific Methods in Archaeology (Accepted), Helsinki, Finland, 20–23 October 2015. [Google Scholar]
- Yang, D.Y.; Eng, B.; Waye, J.S.; Dudar, J.C.; Saunders, S.R. Technical note: Improved DNA extraction from ancient bones using silica-based spin columns. Am. J. Phys. Anthropol. 1998, 105, 539–543. [Google Scholar] [CrossRef]
- Gamba, C.; Jones, E.R.; Teasdale, M.D.; McLaughlin, R.L.; Gonzalez-Fortes, G.; Mattiangeli, V.; Domboróczki, L.; Kovári, I.; Pap, I.; Anders, A.; et al. Genome flux and stasis in a five millennium transect of European prehistory. Nat. Commun. 2014, 5, 5257. [Google Scholar] [CrossRef] [PubMed]
- Gamba, C.; Hanghøj, K.; Gaunitz, C.; Alfarhan, A.H.; Alquraishi, S.A.; Al-Rasheid, K.A.S.; Bradley, D.G.; Orlando, L. Comparing the performance of three ancient DNA extraction methods for high-throughput sequencing. Mol. Ecol. Resour. 2016, 16, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Orlando, L.; Ginolhac, A.; Raghavan, M.; Vilstrup, J.; Rasmussen, M.; Magnussen, K.; Steinmann, K.E.; Kapranov, P.; Thompson, J.F.; Zazula, G.; et al. True single-molecule DNA sequencing of a Pleistocene horse bone. Genome Res. 2011, 21, 1705–1719. [Google Scholar] [CrossRef] [PubMed]
- Ginolhac, A.; Vilstrup, J.; Stenderup, J.; Rasmussen, M.; Stiller, M.; Shapiro, B.; Zazula, G.; Froese, D.; Steinmann, K.E.; Thompson, J.F.; et al. Improving the performance of true single molecule sequencing for ancient DNA. BMC Genom. 2012, 13, 177. [Google Scholar] [CrossRef] [PubMed]
- Der Sarkissian, C.; Ermini, L.; Jónsson, H.; Alekseev, A.N.; Crubezy, E.; Shapiro, B.; Orlando, L. Shotgun microbial profiling of fossil remains. Mol. Ecol. 2014, 23, 1780–1798. [Google Scholar] [CrossRef] [PubMed]
- Damgaard, P.B.; Margaryan, A.; Schroeder, H.; Orlando, L.; Willerslev, E.; Allentoft, M.E. Improving access to endogenous DNA in ancient bones and teeth. Sci. Rep. 2015, 5, 11184. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.V.; Yuhki, N.; Masuda, R.; Modi, W.; O’Brien, S.J. Numt, a recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J. Mol. Evol. 1994, 39, 174–190. [Google Scholar] [PubMed]
- Sorenson, M.D.; Quinn, T.W. Numts: A challenge for avian systematics and population biology. Auk 1998, 115, 214–221. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A. Ancient DNA: Do it right or not at all. Science 2000, 289, 1139. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Pellegrino, I.; Cucco, M.; Follestad, A.; Boos, M. Lack of genetic structure in greylag goose (Anser anser) populations along the European Atlantic flyway. PeerJ 2015, 3, e1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomakina, N.F.; Rozhkov, Y.I.; Linkov, A.B. Direct Submission. GenBank: Accession No. EU601724–EU601734. National Center for Biotechnology Information (NCBI).
- Ruokonen, M.; Litvin, K.; Aarvak, T. Taxonomy of the bean goose-pink-footed goose. Mol. Phylogenet. Evol. 2008, 48, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Honka, J.; Kvist, L.; Heikkinen, M.E.; Helle, P.; Searle, J.B.; Aspi, J. Determining the subspecies composition of bean goose harvests in Finland using genetic methods. Eur. J. Wildl. Res. 2017, 63, 19. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bensasson, D.; Zhang, D.X.; Hartl, D.L.; Hewitt, G.M. Mitochondrial pseudogenes: Evolution’s misplaced witnesses. Trends Ecol. Evol. 2001, 16, 314–321. [Google Scholar] [CrossRef]
- Prost, S.; Anderson, C.N.K. TempNet: A method to display statistical parsimony networks for heterochronous DNA sequence data. Methods Ecol. Evol. 2011, 2, 663–667. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2016. Available online: https://www.r-project.org/ (accessed on 15 January 2018).
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Fisinin, V.I.; Zlochevskaya, K.V. Geese. In Animal Genetic Resources of the USSR; Dmitriev, N.G., Ernst, L.K., Eds.; FAO: Rome, Italy, 1989; pp. 469–506. [Google Scholar]
- Askeyev, I.V. Birds of the Late Holocene of the Middle and Lower Volga Region. Unpublished work.
- Zeder, M.A.; Emshwiller, E.; Smith, B.D.; Bradley, D.G. Documenting domestication: The intersection of genetics and archaeology. Trends Genet. 2006, 22, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.D.; Xie, H.B.; Peng, M.S.; Irwin, D.; Zhang, Y.P. Domestication genomics: Evidence from animals. Annu. Rev. Anim. Biosci. 2014, 2, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Tegetmeier, W.B. The Poultry Book: Comprising the Breeding and Management of Profitable and Ornamental Poultry; to Which is Added “the Standard of Excellence in Exhibition Birds”; Routledge: London, UK, 1867. [Google Scholar]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, G.; Karlsson, E.K.; Perri, A.; Webster, M.T.; Ho, S.Y.W.; Peters, J.; Stahl, P.W.; Piper, P.J.; Lingaas, F.; Fredholm, M.; et al. Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc. Natl. Acad. Sci. USA 2012, 109, 8878–8883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, L.; Persson, H. Natal and breeding dispersal in the Baltic greylag goose Anser anser. Wildfowl 2001, 52, 21–30. [Google Scholar]
- Rychkov, P.I. Topography Orenburg, That is a Detailed Description of the Orenburg Province; Part 1; The Imperial Academy of Sciences: St. Petersburg, Russia, 1762. (In Russian) [Google Scholar]
- Middendorf, A. Travel to the North and East of Siberia 2, 5 Siberian Fauna; The Imperial Academy of Sciences: St. Petersburg, Russia, 1869. (In Russian) [Google Scholar]
- Gray, R. The Birds of the West of Scotland, Including the Outer Hebrides; T. Murray: Glasgow, UK, 1871. [Google Scholar]
- Antonovich, V. (Ed.) Memoirs Related to the History of Southern Russia, Issue 1 (16th Century); Korchak-Novitsky: Kiev, Ukraine, 1890. (In Russian) [Google Scholar]
- Sirelius, U.T. Suomen Kansanomaista Kulttuuria: Esineellisen Kansatieteen Tuloksia; Otava: Helsinki, Finland, 1919. (In Finnish) [Google Scholar]
- Spangenberg, E.P.; Feigin, G.A. Hunting birds of Kyzylorda District of Syrdarya Region. Proc. Forest Exp. Bus. 1930, VII, 157–192. (In Russian) [Google Scholar]
- Zverev, M.D. Predatory Ways of Catching Commercial Birds in the Barabinsk Steppe; News of the Siberian Regional Scientific Hunting and Animal Production Station no. 1: Novosibirsk, Russia, 1930. (In Russian) [Google Scholar]
- Shulpin, L.M. Commercial, Hunting and Predatory Birds of Primorye; Far Eastern Branch of the Academy of Sciences of the USSR: Vladivostok, Russia, 1936. (In Russian) [Google Scholar]
- Tugarinov, A.J. Anseriformes; Fauna of USSR Birds, Edition De L’Academie Des Sciences De l’URSS: Moscow-Leningrad, Russia, 1941. (In Russian) [Google Scholar]
- Rudenko, S.I. The Bashkirs: Historical-Ethnographic Essays; Academy of Sciences of the USSR: Moscow, Russia, 1955. (In Russian) [Google Scholar]
- Spangenberg, E.P. Goose Country. Birds, Hares, Foxes and Others ...: Stories of the Naturalist; Children’s Literature Publishing House: Moscow, Russia, 1973. (In Russian) [Google Scholar]
- Krivushev, A.V.; Vlasov, M.A.; Matev, V.E. Bean goose in the Udorsk Region of the Komi Republic. Casarca 2000, 6, 87. (In Russian) [Google Scholar]
- Ottoni, C.; Girdland Flink, L.; Evin, A.; Geörg, C.; De Cupere, B.; Van Neer, W.; Bartosiewicz, L.; Linderholm, A.; Barnett, R.; Peters, J.; et al. Pig domestication and human-mediated dispersal in western Eurasia revealed through ancient DNA and geometric morphometrics. Mol. Biol. Evol. 2013, 30, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Frantz, L.A.F.; Schraiber, J.G.; Madsen, O.; Megens, H.J.; Cagan, A.; Bosse, M.; Paudel, Y.; Crooijmans, R.P.M.A.; Larson, G.; Groenen, M.A.M. Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes. Nat. Genet. 2015, 47, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Park, S.D.E.; Magee, D.A.; McGettigan, P.A.; Teasdale, M.D.; Edwards, C.J.; Lohan, A.J.; Murphy, A.; Braud, M.; Donoghue, M.T.; Liu, Y.; et al. Genome sequencing of the extinct Eurasian wild aurochs, Bos primigenius, illuminates the phylogeography and evolution of cattle. Genome Biol. 2015, 16, 234. [Google Scholar] [CrossRef] [PubMed]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Kashani, B.H.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warmuth, V.; Eriksson, A.; Bower, M.; Barker, G.; Barrett, E.; Hanks, B.; Li, S.; Lomitashvili, D.; Ochir-Goryaeva, M.; Sizonovg, G.V.; et al. Reconstructing the origin and spread of horse domestication in the Eurasian steppe. Proc. Natl. Acad. Sci. USA 2012, 109, 8202–8206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, B.; Marshall, F.B.; Chen, S.; Rosenbom, S.; Moehlman, P.D.; Tuross, N.; Sabin, R.C.; Peters, J.; Barich, B.; Yohannes, H.; et al. Ancient DNA from Nubian and Somali wild ass provides insights into donkey ancestry and domestication. Proc. R. Soc. B Biol. Sci. 2011, 278, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almathen, F.; Charruau, P.; Mohandesan, E.; Mwacharo, J.M.; Orozco-TerWengel, P.; Pitt, D.; Abdussamad, A.M.; Uerpmann, M.; Uerpmann, H.P.; De Cupere, B.; et al. Ancient and modern DNA reveal dynamics of domestication and cross-continental dispersal of the dromedary. Proc. Natl. Acad. Sci. USA 2016, 113, 6707–6712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyazov, L.A. The Socio-Economic Development of the Population of the Middle Volga Region in the Middle of the First Millennium AD (Based on the Materials of Imenkov Culture). Thesis for a Candidate of Historical Science, Tatarstan Academy of Sciences, Sh. Marjani Institute of History, Kazan, Russia, 2011. (In Russian). [Google Scholar]
- Voinstvensky, M.A. Ornithofauna of Olbia. Archaeological Sites of Ukraine. No 7; Institute of Archeology: Kiev, Ukraine, 1958. (In Ukrainian) [Google Scholar]
- Voinstvensky, M.A. Fossil Avifauna of Ukraine; The Natural Environment and the Fauna of the Past: Kiev, Ukraine, 1967. (In Russian) [Google Scholar]
- Burchak-Abramovich, N.I.; Tsalkin, V.I. Birds from archaeological excavations in the Moscow Kremlin. Bull. Mosc. Soc. Nat. (Biol. Ser.) 1969, 74, 49–53. (In Russian) [Google Scholar]
- Burchak-Abramovich, N.I.; Tsalkin, V.I. To the knowledge of the avifauna of South Ukraine, Crimea and the Don Region (according to archaeological materials). Bull. Mosc. Soc. Nat. (Biol. Ser.) 1971, 76, 54–63. (In Russian) [Google Scholar]
- Burchak-Abramovich, N.I.; Tsalkin, V.I. Materials for the study of European birds of RSFSR (according to archaeological sites). Bull. Mosc. Soc. Nat. (Biol. Ser.) 1972, 77, 51–59. (In Russian) [Google Scholar]
- Bryuzgina (Umanskaya), A.S. Late Anthropogenic Birds from the Ukraine and Contiguous Territories (Primarily Based on Materials from Archeological Sites). Thesis for a Candidate of Biological Science, Institute of Zoology, Kiev, Ukraine, 1975. (In Russian). [Google Scholar]
- Koshelenko, G.A.; Kruglikova, I.T.; Dolgorukov, V.S. (Eds.) Ancient States of the Northern Black Sea Littoral; Archeology of the USSR, Nauka: Moscow, Russia, 1984. (In Russian) [Google Scholar]
- Gorobets, L.; Kovalchuk, O. Birds in the medieval culture and economy of the East Slavs in the 10–13th centuries AD. Environ. Archeol. 2017, 22, 147–165. [Google Scholar] [CrossRef]
- Nekrasov, A.E. Bone Remains of Birds from the Holocene Locations of the Urals and Western Siberia; Quaternary paleozoology in the Urals: Yekaterinburg, Russia, 2003. (In Russian) [Google Scholar]
- Martynovich, N.V. Birds of “Gold-Fired” Mangazeya. Zool. J. 2013, 92, 1129–1135. (In Russian) [Google Scholar] [CrossRef]
- Askeyev, I.V.; Askeyev, O.V. History of Domestic Birds of the Finno-Ugrish people in Volga-Ural Region. Report 2018 of Biomonitoring Laboratory of The Institute of Problems in Ecology and Mineral Wealth, Tatarstan Academy of Sciences. Unpublished work.
- Askeyev, I.V.; Askeyev, O.V.; Askeyev, A.O. Anseriformes Spring and Autumn phenology. Report 2018 of Biomonitoring Laboratory of The Institute of Problems in Ecology and Mineral Wealth, Tatarstan Academy of Sciences. Unpublished work.
- Eversmann, E. Natural History of Birds of Orenburg Territory; Kazan University Press: Kazan, Russia, 1866. (In Russian) [Google Scholar]
- Bogdanov, M.N. Birds and mammals in the blacksoil zone of the Volga Region and in the valleys of the Middle and Lower Volga river. Proc. Kazan Nat. Soc. 1871, 1, 3–226. (In Russian) [Google Scholar]
- Ruzsky, M.D. The results of investigations of birds in the Kazan province. Proc. Kazan Nat. Soc. 1893, 25, 1–394. (In Russian) [Google Scholar]
- Bashkirov, I.S.; Grigoryev, N.D. Essay on the Hunting of Tataria; Works of the Volga-Kama Regional Commercial-Biological Station v.1: Kazan, Russia, 1931. (In Russian) [Google Scholar]
- Artemiev, J.T.; Popov, V.A. Order of Anseriformes. The Birds of the Volga-Kama Region: Non-Passeriformes; Popov, V.A., Ed.; Nauka: Moscow, Russia, 1977. (In Russian) [Google Scholar]
- Askeyev, I.V.; Askeyev, O.V. Birdfauna of Tatarstan Republic; Akademia Nauk Tatarstana: Kazan, Russia, 1999. (In Russian) [Google Scholar]
- McCarthy, E.M. Handbook of Avian Hybrids of the World; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Hofreiter, M.; Paijmans, J.L.A.; Goodchild, H.; Speller, C.F.; Barlow, A.; Fortes, G.G.; Thomas, J.A.; Ludwig, A.; Collins, M.J. The future of ancient DNA: Technical advances and conceptual shifts. Bioessays 2015, 37, 284–293. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | n | H | h (SD) | π (SD) | D | Fs |
|---|---|---|---|---|---|---|
| Present | 102 | 7 | 0.584 (0.030) | 0.0056 (0.0012) | −1.207 | −0.753 |
| Post-Medieval | 22 | 3 | 0.541 * (0.068) | 0.0056 (0.0028) | −1.573 | 1.652 |
| High and Late Medieval | 16 | 3 | 0.658 ** (0.075) | 0.0134 † (0.0042) | 0.482 | 3.924 |
| Time Period | Present (n = 102) | Post-Medieval (n = 22) |
|---|---|---|
| Post-Medieval (n = 22) | 0.026 | |
| High and Late Medieval (n = 16) | 0.105 * | 0.064 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honka, J.; Heino, M.T.; Kvist, L.; Askeyev, I.V.; Shaymuratova, D.N.; Askeyev, O.V.; Askeyev, A.O.; Heikkinen, M.E.; Searle, J.B.; Aspi, J. Over a Thousand Years of Evolutionary History of Domestic Geese from Russian Archaeological Sites, Analysed Using Ancient DNA. Genes 2018, 9, 367. https://doi.org/10.3390/genes9070367
Honka J, Heino MT, Kvist L, Askeyev IV, Shaymuratova DN, Askeyev OV, Askeyev AO, Heikkinen ME, Searle JB, Aspi J. Over a Thousand Years of Evolutionary History of Domestic Geese from Russian Archaeological Sites, Analysed Using Ancient DNA. Genes. 2018; 9(7):367. https://doi.org/10.3390/genes9070367
Chicago/Turabian StyleHonka, Johanna, Matti T. Heino, Laura Kvist, Igor V. Askeyev, Dilyara N. Shaymuratova, Oleg V. Askeyev, Arthur O. Askeyev, Marja E. Heikkinen, Jeremy B. Searle, and Jouni Aspi. 2018. "Over a Thousand Years of Evolutionary History of Domestic Geese from Russian Archaeological Sites, Analysed Using Ancient DNA" Genes 9, no. 7: 367. https://doi.org/10.3390/genes9070367