1. Introduction
Marfan syndrome (MFS; OMIM #154700) is an autosomal dominant connective tissue disorder with an estimated prevalence of 1/5000 individuals. It is a multisystemic disease that affects the ocular, skeletal, and cardiovascular system as well as lung, skin, and dura. Thoracic aortic aneurysm (TAA) is the main cardiovascular feature that can lead to dissection or rupture of the aortic wall, the major life-threatening event in MFS. In most cases, MFS is due to mutation in the
FBN1 gene encoding fibrillin-1, an extracellular matrix protein. To date, over 3000 mutations have been reported in the
FBN1 database [
1,
2].
The syndrome displays great clinical variability, regarding age of onset and number and severity of the symptoms that cannot be explained by the few genotype-phenotype correlations identified between
FBN1 mutations and MFS. Furthermore, this variability is observed not only between families, but also among relatives. Genotype-phenotype correlation [
3,
4,
5] could explain interfamilial variability but not the intrafamilial variability, which suggests the existence of genetic factors underlying phenotypic variability.
Recently, our team investigated the hypothesis that phenotype severity could be related to the variable expression level of
FBN1 synthesized from the wildtype (WT) allele [
6].
FBN1 gene expression levels were assessed in culture of skin fibroblasts from 80 controls and from 80 MFS patients with premature termination codon (PTC) mutations. Results in controls showed a 3.9-fold variation in
FBN1 messenger RNA (mRNA) synthesis level between controls. A similar 4.4-fold variation was found in the MFS population, but the mean level of
FBN1 expression level was half of the control population. Differential allelic expression analysis in MFS fibroblasts showed that no residual expression of the mutated allele was detected. Moreover, a low level of residual WT
FBN1 mRNA accounted for a high risk of ectopia lentis and pectus abnormality and tended to increase the severity of aortic dilatation. In the control population, independently of the expression level of
FBN1, we observed a steady-state equilibrium between the two allelic expressions, suggesting that the
FBN1 expression level mainly depends on trans-acting regulators [
6].
An expression quantitative trait loci (eQTL) analysis was performed to identify trans-acting regulators of
FBN1 expression, in PTC-mutation carriers. A significant signal that reached genome-wide significant threshold was identified on chromosome 11 (
p = 6 × 10
−8) at 11q22.3 (gMod-M3) [
5]. This trans-eQTL was located in a linkage disequilibrium (LD) block and delineating a 40 kb region containing only one expressed gene,
SLN (encoding sarcolipin), and a single pseudogene,
SNX7-ps1 (CTD-2651C21.3) [
5]. Since the lead single nucleotide polymorphism (SNP) had never been highlighted in any reported genome-wide study, we first investigated the region and then looked for association between the genes in the region and
FBN1 expression.
2. Materials and Methods
2.1. Patient and Control Samples
Marfan syndrome patients were recruited in the ‘Centre National Maladies Rares—Syndrome de Marfan et apparentés’, the French National Reference Centre located at Bichat Hospital (Paris, France). Clinical diagnosis and systemic score were established according to the revised Ghent nosology as already reported in Aubart et al [
5]. For the purpose of this study, skin fibroblasts from 40 MFS patients carrying a PTC-
FBN1 mutation were selected according to their genotype at rs11212346:20 with [CC] genotype, 19 with [CT] genotype, and 1 with [TT] genotype. All patients with a [CT] genotype and [TT] genotype were selected and matched with 20 [CC] genotype patients. To develop and validate the various tests, three control skin fibroblasts were used.
All patients originated from the French National Reference Centre for Marfan Syndrome and related disorders and gave their written informed consent for participation in this clinical and genetic study in agreement with the requirements of French regulations (Accepted by “Comite’ de Protection des Personnes CPP Ile de France XI”, 78105 St Germain en Laye; with the registration number #11008).
2.2. Cell Culture and RNA Purification
Patient skin fibroblasts were cultivated in DMEM (Thermo Scientific, Villebon sur Yvette, France) supplemented with 4.5 g/L glucose, 15% Fetal Bovine Serum (FBS, PAA Laboratory, Villacoublay, France), and antibiotics (50 U/mL of penicillin, streptomycin, and amphotericin B) (PAA Laboratory, Villacoublay, France) as previously described [
6].
From fibroblast culture, total RNAs were extracted with the miRNeasy kit® (Qiagen S.A., Courtaboeuf, France) according to the manufacturer’s instructions. After purification, RNAs were eluted in 20 µL of RNase-free water and RNA concentrations were estimated by measuring absorbance at 260 nm, using Nanodrop 2000/2000 c system (Thermo Scientific, Villebon sur Yvette, France).
2.3. Digital Droplet PCR (ddPCR)
To assess expression level of low-expressed genes, a digital droplet PCR (ddPCR) was performed. Reverse transcription was performed using miScript II RT kit (Qiagen S.A., Courtaboeuf, France), according to the manufacturer’s recommendations. Droplet digital PCR was performed at a concentration of 50 ng/µL per well. For each assay, the gene of interest (SLN or SNX7-ps1) and control gene (SDHA) were run in multiplex using 6-carboxyfluoresceine (FAM) and hexachloro-fluorescein (HEX) labeling, respectively. Reaction mixtures were prepared using ddPCR™ Supermix for Probes (BioRad, Marnes-la-Coquette, France), DNA, and PrimePCR Probe Assay (BioRad, Marnes-la-Coquette, France) according to the manufacturer’s recommendations. Droplets were generated from the mixtures using Droplet Generation Oil for Probes (BioRad, Marnes-la-Coquette, France) on the QX100 Droplet Generator (BioRad, Marnes-la-Coquette, France) according to the manufacturer’s instructions. PCR was carried out on a thermal cycler for 40 cycles using a 55 °C annealing temperature and fluorescent level of each droplet was read using the QX100 Droplet Reader (BioRad, Marnes-la-Coquette, France). Data were analyzed and the concentration of mRNA levels for both genes and the ratio of gene of interest/reference gene were calculated using QuantaSoft software (BioRad, Marnes-la-Coquette, France). Each DNA sample was run in two technical replicates and three biological replicates.
2.4. Real-Time Quantitative PCR (RT-qPCR)
Reverse transcription was performed using miScript II RT kit, according to the manufacturer’s recommendations. The final concentration of complementary DNA (cDNA) was 100 ng/µL for each sample. RT-qPCR was performed using ABI PRISM 7300 (Applied Biosystems, Thermo Scientific, Waltham, MA, USA), according to the manufacturer’s instructions. Absolute Blue qPCR SYBR Green supermix (Thermo Scientific, Villebon sur Yvette, France) and specific primers (
Table S1) were used to quantify
FBN1,
SNX7,
FOS,
SDHA, and
GAPDH gene expression. Results were then normalized with the expression of the two housekeeping genes (
SDHA and
GAPDH) by calculating the geometric means [
7].
2.5. Cell Culture and Pharmacological Treatment with Calcium Ionophore, A23187
For the pharmacological treatment, 7 × 10
5 skin fibroblasts were seeded in each well of 4-well plates (21.8 cm
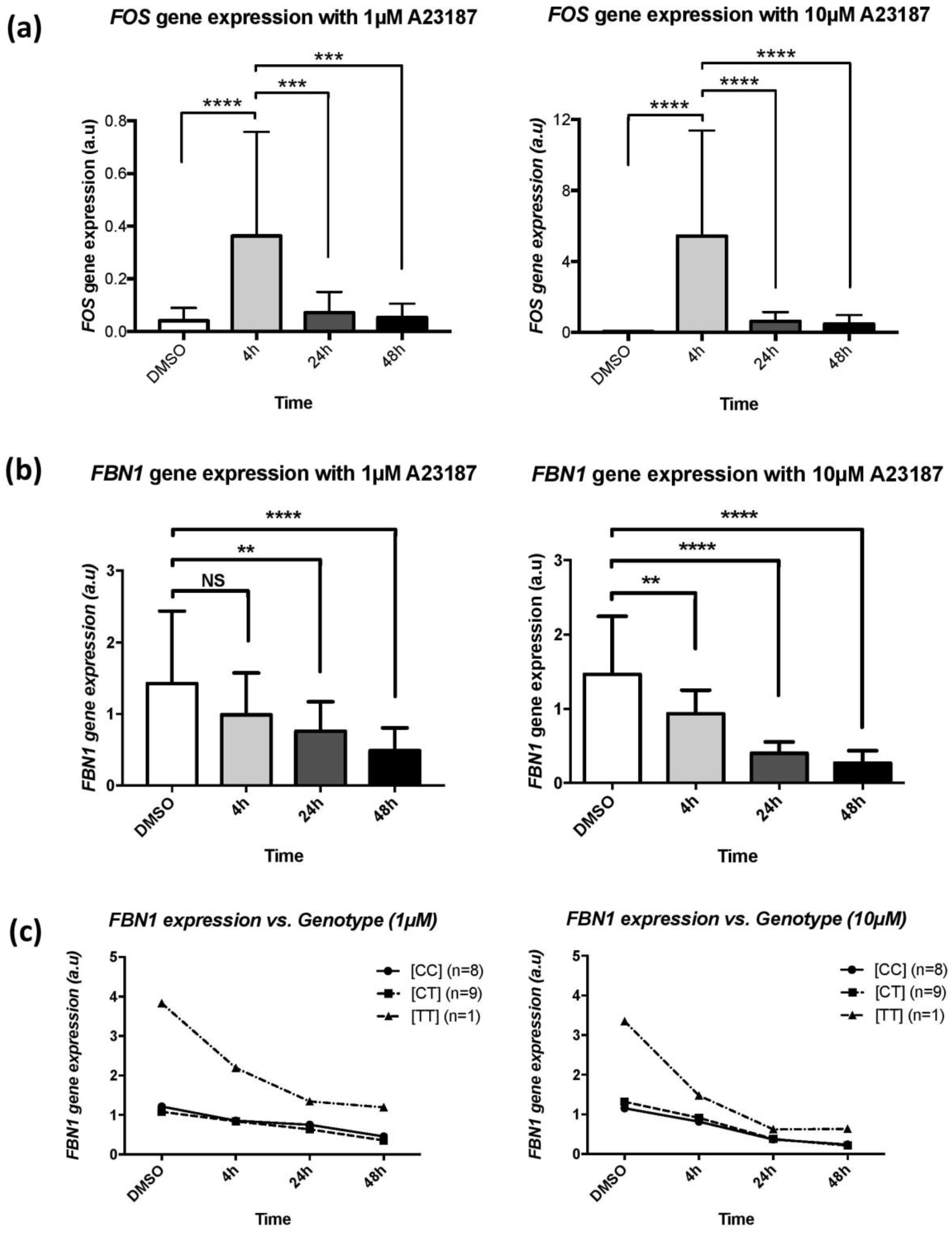
2/well). Cells were incubated with A23187, a calcium ionophore (Sigma-Aldrich, Darmstadt, Germany). Each cell preparation was treated with two doses of A23187, 1 µM and 10 µM, in duplicates and at three different times of incubation (4, 24, and 48 h). Skin fibroblasts were deprived of FBS and antibiotics 24 h before treatment. A23187 was solubilized in DMSO and diluted in FBS-free DMEM before its addition to cells. In control wells, a similar volume of DMSO, diluted in FBS-free DMEM, was added. Expression levels of
FOS gene were quantified as a positive control for the entry of calcium in the cells [
8].
2.6. Mimic and micro RNA (miRNA) Transfection
TargetScan [
9] was used to predict biological targets of micro RNAs (miRNAs) by searching for the presence of conserved sequenced corresponding to miRNA binding site. P
CT (probability of conserved targeting) estimates the probability of the site being preferentially conserved because it is targeted by the cognate miRNA [
10].
For the transfection assay, 6 × 105 skin fibroblasts were seeded in each well of 4-well plates (21.8 cm2/well). Transfection was performed using Hiperfect Transfection Reagent (Qiagen S.A., Courtaboeuf, France), 150 ng miScript miRNA Mimic (Syn-hsa-miR-9-5) (Qiagen S.A., Courtaboeuf, France) or 1500 ng miScript miRNA Inhibitor (Anti-hsa-miR-9-5) (Qiagen S.A., Courtaboeuf, France), according to the manufacturer’s recommendation. For controls, transfection control (Hiperfect Transfection Reagent only) and a negative control (1500 ng miScript Inhibitor Negative Control; Qiagen S.A., Courtaboeuf, France) had been tested. Each condition had been tested in three biological replicates.
Total RNA were extracted 72 h after transfection with the miRNeasy kit® according to the manufacturer’s instructions. After purification, RNAs were eluted in 15 µL of RNase-free water and the concentrations of RNA were estimated by measuring absorbance at 260 nm, using Nanodrop 2000/2000 c system. Reverse transcription was performed as above.
2.7. Statistical Analysis
Statistical analysis was performed using GraphPad Prism version 7.0 software. The statistical difference between two groups was tested using nonparametric Mann–Whitney U test, and between three groups using One Way ANOVA or Kruskall–Wallis test. The coefficient of correlation (r2) was calculated by linear regression and the statistical significance by Spearman correlation.
4. Discussion
This work was aimed at investigating the mechanism underlying the trans-eQTL signal (
p = 6 × 10
−8) for
FBN1 expression in fibroblast on chromosome 11. This trans-eQTL is located in an LD block, which contains only one expressed gene,
SLN, and a single pseudogene,
SNX7-ps1. Since the lead SNP (rs11212346) had never been highlighted in any reported genome-wide study, we first investigated the region and then looked for association between the genes in the region and
FBN1 expression. We showed that the
SLN gene is weakly expressed in skin fibroblasts in controls and in MFS subjects. We also showed that the pseudogene is expressed and that expressions of
SLN and
SNX7-ps1 are correlated. To our knowledge, this has never been reported before for these two genes even if the mechanism has been described [
13]. Furthermore, since we have shown that skin fibroblasts are a surrogate for adventitial fibroblasts [
6], it is highly probable that this observation extends to this cell type.
The presence of a potential functional variant in SLN responsible for the variability of FBN1 expression was investigated but not found. One of the SNPs in the LD block, rs7104725, is located in the pseudogene coding sequence and could be involved in a gene expression regulatory mechanism.
We confirmed in skin fibroblasts that
SNX7-ps1 is an expressed pseudogene with a positive correlation between the
SLN gene and
SNX7-ps1, suggesting the role of a common regulatory element on their expression. Functional implication of pseudogenes in physiological processes is still poorly understood. However, they are known to possibly act as regulatory antisense transcripts or miRNA decoys and we investigated these two possible mechanisms [
14].
Pseudogenes can function as antisense transcript, either by direct hybridization on its parent gene or by generating siRNA leading to the repression of the targeted gene expression [
14]. The absence of homologous sequences and no correlation of expression between
SNX7-ps1 and the
FBN1 gene exclude
FBN1 as a direct target of
SNX7-ps1. However, downstream target of a regulatory element such as transcription factor can establish eQTL association with the same locus if the effect is strong enough [
15]. Therefore, a downstream target of
SNX7-ps1 could be responsible for
FBN1 gene regulation. We hypothesized that the
SNX7 gene could be an
FBN1 gene regulator. In our cohort, we found no correlation between
SNX7-ps1 and
SNX7 gene expression, suggesting that
SNX7-ps1 has no regulation function on
SNX7. Moreover, the absence of correlation between rs11212346 genotype and
SNX7 as well as between
SNX7 and
FBN1 expression did not support a role of
SNX7 in
FBN1 gene regulation.
Another potential mechanism is the ability of processed pseudogenes to interfere with factors of mRNA stability. They can act as miRNA decoy: due to sequence homology between pseudogenes and miRNA target, pseudogenes can serve as bait and can compete with mRNA target for miRNA silencing [
14,
16]. In silico analysis demonstrated complementarity binding site between miR9,
SNX7,
SNX7-ps1, and
FBN1, placing miR9 as a potential common regulator. Therefore, we investigated a potential role of miR9 in
FBN1 gene regulation. The absence of correlation between miR9 and
SNX7,
SNX7-ps1 or
FBN1 expression did not support the role of miR9. However, miR9 is weakly expressed in skin fibroblasts and this could prevent us from observing any correlation. To counter this limitation, we transfected patients’ skin fibroblasts with a synthetic miR9 or an siRNA-targeting miR9. No difference of expression was observed for
SNX7 or
SNX7-ps1 in the different transfection conditions. Regarding
FBN1 gene expression, no significant difference was observed, but a trend with a decreased expression in the presence of miR9 and an increased expression in the absence of miR9 was observed. Moreover, a recent study showed that a down-regulation of miR9 has been associated with an increased expression level of
MMP13, a matrix metalloproteinase that degrades collagen and other matrix components [
17]. This study implicates miR9 in the extracellular matrix stability, known to be important for
FBN1 microfibrils’ proper assembly [
18]. Therefore, miR9 may have an impact on
FBN1 gene regulation independently of the eQTL region.
Sarcolipin is a 31-amino-acid protein, highly expressed in skeletal muscle and at a lower extent in the atria [
11]. Sarcolipin inhibits the activity of SERCA1 pumps in cardiac and skeletal muscle by decreasing its apparent affinity for Ca
2+ [
12]. During Ca
2+ transport, SERCA transfers two molecules of Ca
2+ per molecule of ATP hydrolysed and this process contributes to muscle thermogenesis [
19,
20]. Sarcolipin is a regulator of Ca
2+ homeostasis, playing a critical role in cardiac contractility and cardiac pathophysiology [
21]. Based on all of this information, the absence of a direct action of
SLN on
FBN1 gene expression was not completely surprising. Indeed, there is no evidence in the literature that sarcolipin has a transcription factor activity giving it the ability to directly regulate gene expression. However, due to sarcolipin’s critical role on Ca
2+ bioavailability, we hypothesized that sarcolipin could indirectly impact
FBN1 gene expression by altering intracellular Ca
2+ homeostasis. To support the idea, Lannoy et al. showed that an increase of intracellular free calcium concentration, [Ca
2+]
i, had an effect on gene expression of several genes encoding for constitutive proteins of elastic fibers or involved in elastic fiber stability [
22]. By using a Ca
2+ ionophore, A23187, they increased [Ca
2+]
i in vascular smooth muscle cells of Brown Norway rats and assessed its impact on the gene expression pattern. A decrease in elastic mRNA content was associated with the increase of [Ca
2+]
i and the same pattern was observed for several elastic fiber-associated proteins, such as fibrillin-1. Indeed, the increase of [Ca
2+]
i decreased
FBN1 gene expression in a significant manner. Moreover, Doyle et al [
23] investigated the impact of calcium channel blockers (CCB), an antihypertensive compound, on TAA expansion. They showed that
Fbn1C1039G/− mice (modelling MFS) treated with CCB had accelerated TAA expansion and rupture through a protein kinase C-mediated pathway. In a retrospective study on patients with MFS treated with CCB, they showed that MFS patients displayed an increased risk of aortic dissection compared to patients treated with other antihypertensive compounds. These results support a cross-talk between calcium signalling, TGFβ, and aneurysms [
23]. Our findings in human dermal fibroblasts extracted from MFS patients support the results of Lannoy et al [
22]. Indeed, we observed a significant decrease of
FBN1 expression, in a time- and dose-dependant manner, confirming the role of calcium on
FBN1 gene regulation.
The modification of FBN1 expression was also observed for all MFS patients with comparable pattern for the rs11212346 [CC] and [CT] genotype subjects. Interestingly, the homozygous [TT] patient displayed a much stronger response to calcium cellular influx. Therefore, it could be speculated that homozygote [TT] subjects display a possible inappropriate answer to calcium intracellular modifications. A targeted recruitment of patients with this specific genotype is warranted to confirm this result. Overall, we were able to confirm the impact of [Ca2+]i on FBN1 gene expression but we could not conclude regarding the role of sarcolipin and/or the eQTL locus in this regulation.
In conclusion, our data provide insight into the expression pattern of the SLN and SNX7-ps1 genes at 11q22.3 but further work is necessary to unravel the mechanism underlying the trans-eQTL signal for FBN1 expression in fibroblasts. Indeed, there is the possibility that the lead SNP could be in LD with functional variants or a regulatory SNP outside of the LD block. Finally, another avenue of research could be the investigation at the chromatin level of the area.

 ,
, 



{kind=link}
{kind=link}
{kind=link}