Evaluation of Restoration and Flow Interactions on River Structure and Function: Channel Widening of the Thur River, Switzerland

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
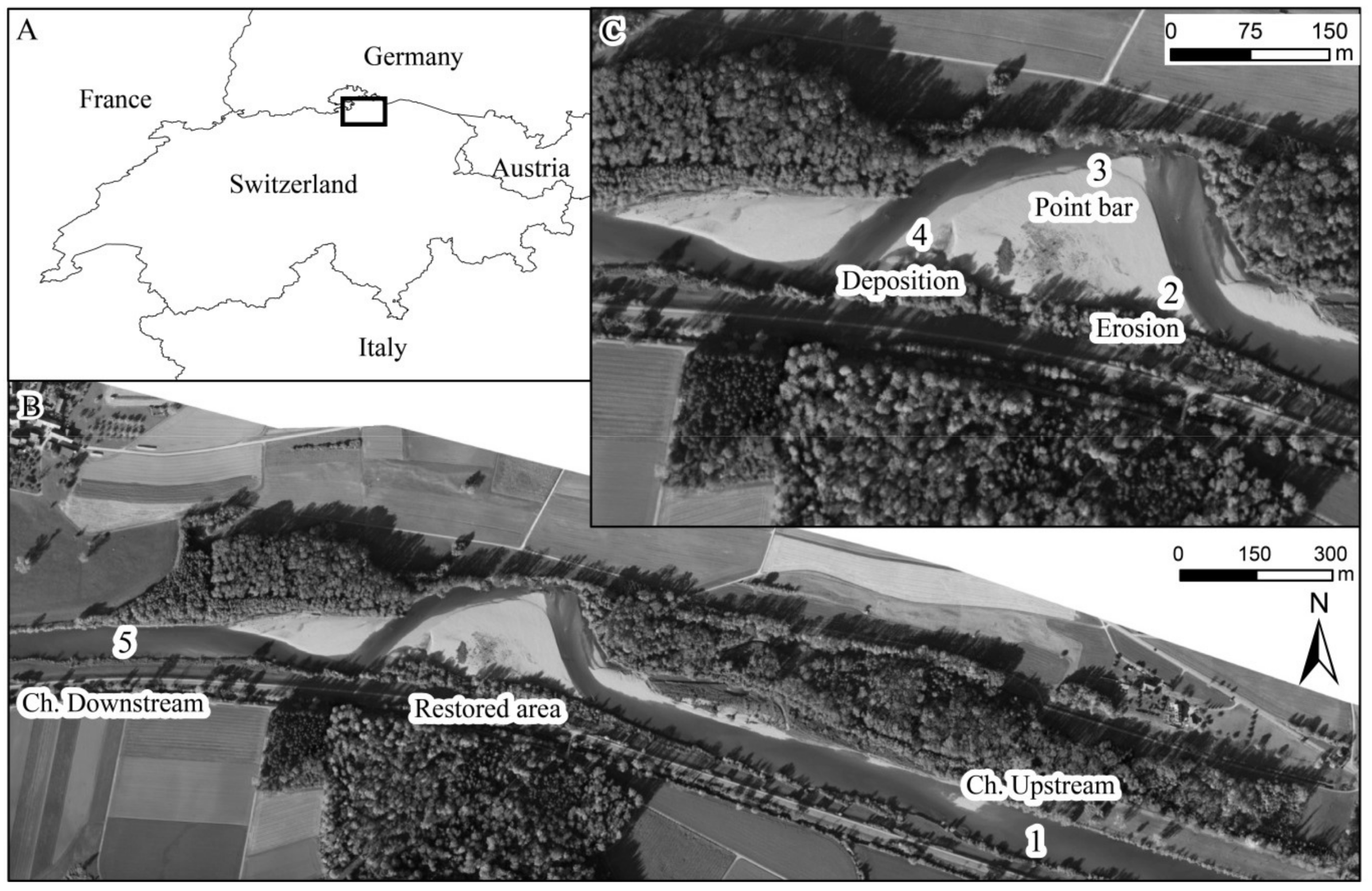
2.1. Study Site
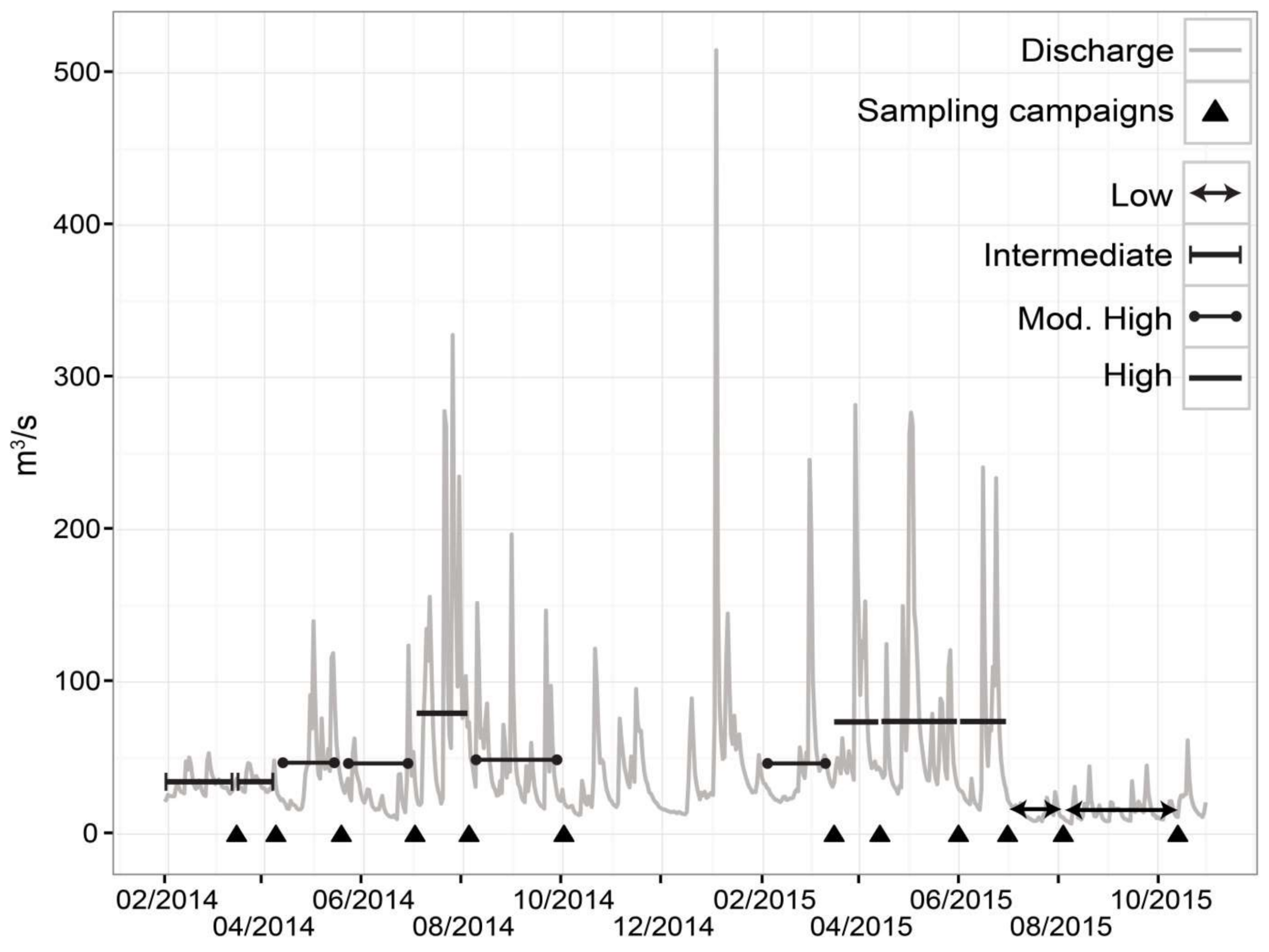
2.2. Flow Regime Classification
2.3. Physico-Chemistry and Abiotic Hyporheic Measures
2.4. Sediment Respiration and Primary Production
2.5. Macroinvertebrate Density and Taxa Richness
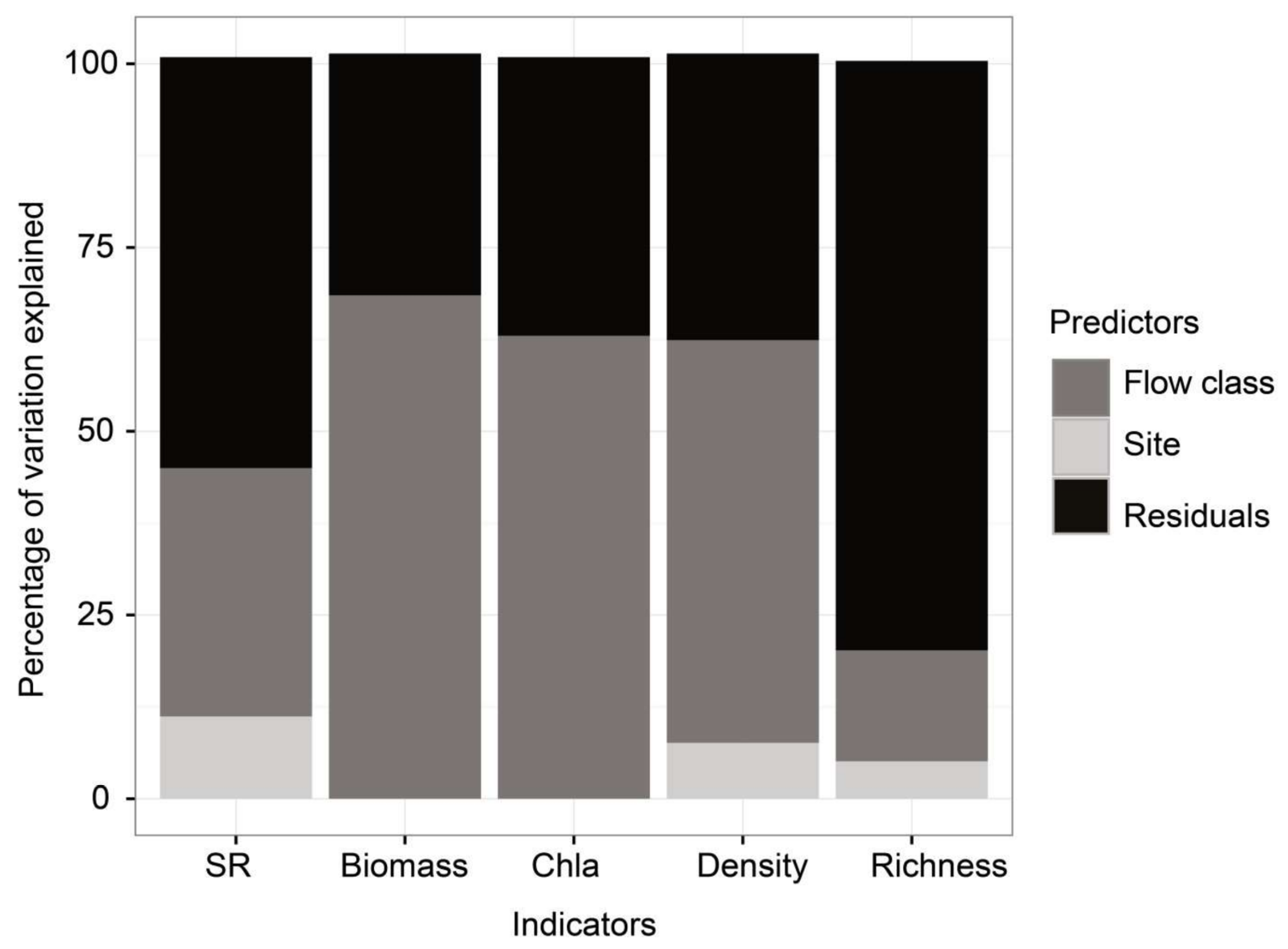
2.6. Statistical Analysis
3. Results
3.1. Flow Classes
3.2. Abiotic Factors
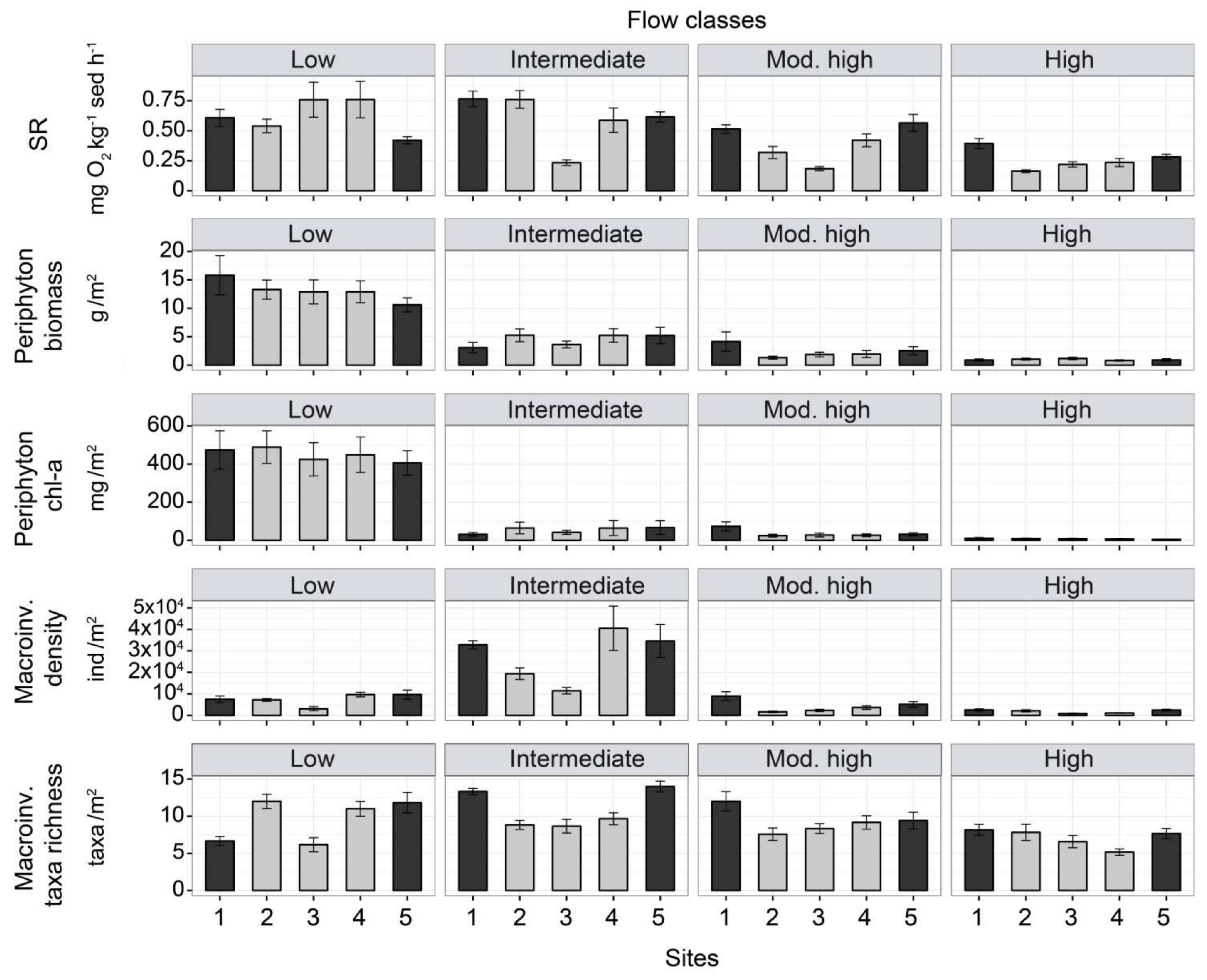
3.3. Ecosystem Function
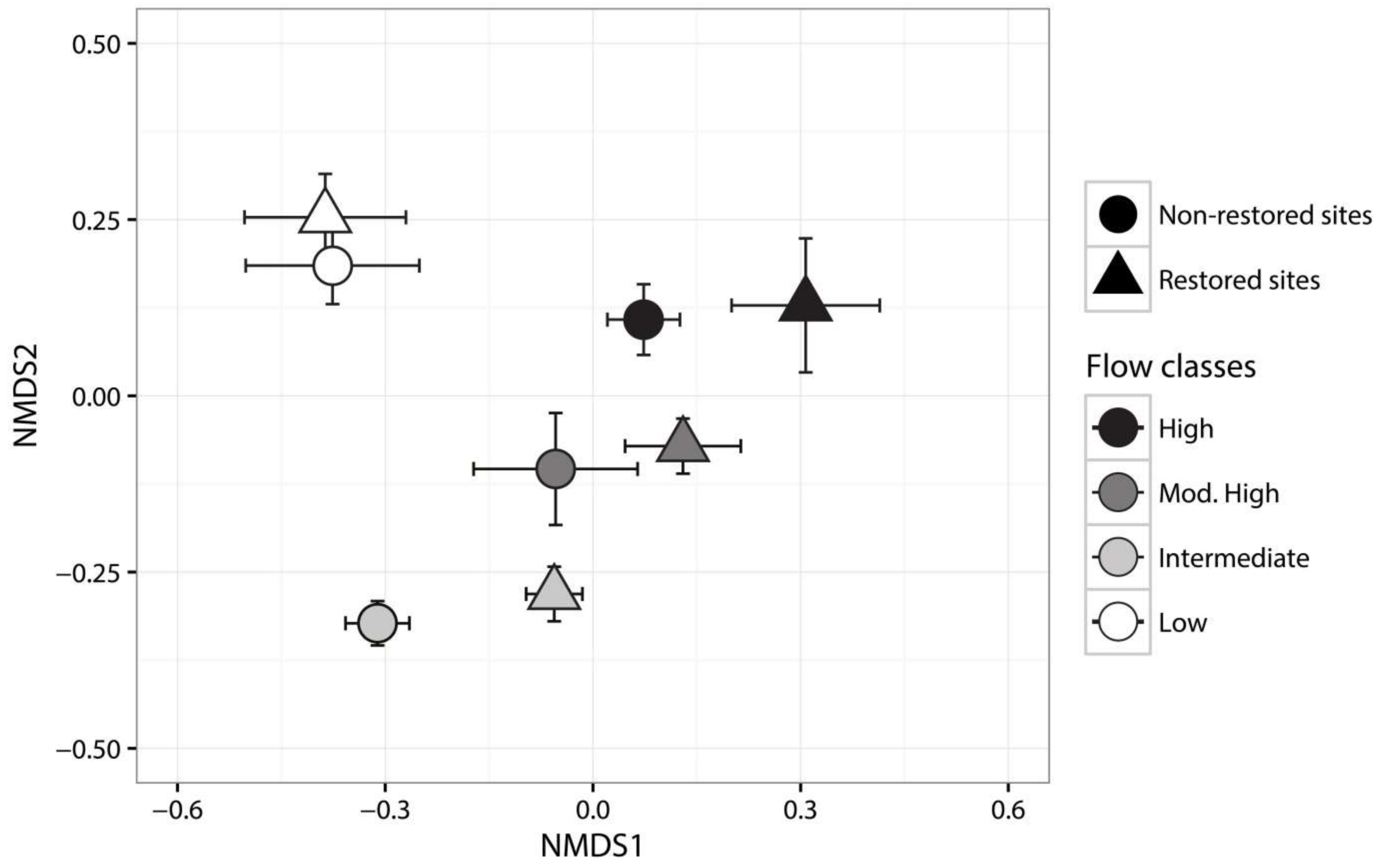
3.4. Ecosystem Structure
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Resh, V.H.; Brown, A.V.; Covich, A.P.; Gurtz, M.E.; Li, H.W.; Minshall, G.W.; Reice, S.R.; Sheldon, A.L.; Wallace, J.B.; Wissmar, R.C. The role of disturbance in stream ecology. J. N. Am. Benthol. Soc. 1988, 7, 433–455. [Google Scholar] [CrossRef]
- Poff, N.L.; Ward, J.V. Implications of streamflow variability and predictability for lotic community structure: A regional analysis of streamflow patterns. Can. J. Fish. Aquat. Sci. 1989, 46, 1805–1818. [Google Scholar] [CrossRef]
- Bunn, S.E.; Arthington, A.H. Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ. Manag. 2002, 30, 492–507. [Google Scholar] [CrossRef]
- Richter, B.D.; Baumgartner, J.V.; Powell, J.; Braun, D.P. A method for assessing hydrologic alteration within ecosystems. Conserv. Boil. 1996, 10, 1163–1174. [Google Scholar] [CrossRef]
- Lytle, D.A.; Poff, N.L. Adaptation to natural flow regimes. Trends Ecol. Evol. 2004, 19, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.J.; Armitage, P.D. The response of the macroinvertebrate community to low-flow variability and supra-seasonal drought within a groundwater dominated stream. Arch. Hydrobiol. 2004, 161, 1–20. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Ryo, M.; Sui, P.; Yoshimura, C. Evaluating the relationship between basin-scale fish species richness and ecologically relevant flow characteristics in rivers worldwide. Freshw. Biol. 2012, 57, 2173–2180. [Google Scholar] [CrossRef]
- Wohl, E.E.; Bledsoe, B.P.; Jacobson, R.B.; Poff, N.L.; Rathburn, S.L.; Walters, D.M.; Wilcox, A.C. The natural sediment regime in rivers: Broadening the foundation for ecosystem management. Bioscience 2015, 65, 358–371. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Habitat, the template for ecological strategies? J. Anim. Ecol. 1977, 46, 337–365. [Google Scholar] [CrossRef]
- Lytle, D.A.; Bogan, M.T.; Finn, D.S. Evolution of aquatic insect behaviours across a gradient of disturbance predictability. Proc. R. Soc. Lond. B Biol. Sci. 2008, 275, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.I.; Murphy, J.F.; Collins, A.L.; Sear, D.A.; Naden, P.S.; Armitage, P.D. The impact of fine sediment on macroinvertebrates. River Res. Appl. 2012, 28, 1055–1071. [Google Scholar] [CrossRef]
- Merritt, D.M. Reciprocal relations between riparian vegetation, fluvial landforms, and channel processes. In Treatise on Geomorphology; Academic Press: Cambridge, MA, USA, 2013; pp. 219–243. [Google Scholar]
- Benke, A.C. Importance of flood regime to invertebrate habitat in an unregulated river-floodplain ecosystem. N. Am. Benthol. Soc. 2001, 20, 225–240. [Google Scholar] [CrossRef]
- Surian, N.; Rinaldi, M. Morphological response to river engineering and management in alluvial channels in Italy. Geomorphology 2003, 50, 307–326. [Google Scholar] [CrossRef]
- Brandt, S.A. Classification of geomorphological effects downstream of dams. Catena 2000, 40, 375–401. [Google Scholar] [CrossRef]
- Tena, A.; Batalla, R.J. The sediment budget of a large river regulated by dams (The lower River Ebro, NE Spain). J. Soils Sediments 2013, 13, 966–980. [Google Scholar] [CrossRef]
- Zeh, W.H.; Könitzer, C.; Bertiller, A. Structures of Watercourses in Switzerland; Swiss Federal Office of Environment: Bern, Switzerland, 2009. [Google Scholar]
- Lach, J.; Wyzga, B. Channel incision and flow increase of the upper Wisłoka River, southern Poland, subsequent to the reafforestation of its catchment. Earth Surf. Process. Landf. 2002, 27, 445–462. [Google Scholar] [CrossRef]
- Tockner, K.; Stanford, J.A. Riverine floodplains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef]
- Gregory, K.J. The human role in changing river channels. Geomorphology 2006, 79, 172–191. [Google Scholar] [CrossRef]
- European-Commission. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for community action in the field of water policy. Off. J. Eur. Community 2000, L327, 1–72.
- Yarnell, S.M. Quantifying physical habitat heterogeneity in an ecologically meaningful manner: A case study of the habitat preferences of the foothill yellow-legged frog (Rana boylii). In Landscape Ecology Research Trends; Dupont, A., Jacobs, H., Eds.; Nova Science: Hauppauge, NY, USA, 2008; pp. 89–112. [Google Scholar]
- Richards, K.; Brasington, J.; Hughes, F. Geomorphic dynamics of floodplains: Ecological implications and a potential modelling strategy. Freshw. Biol. 2002, 47, 559–579. [Google Scholar] [CrossRef]
- Seidl, R.; Stauffacher, M. Evaluation of river restoration by local residents. Water Resour. Res. 2013, 10, 7077–7087. [Google Scholar] [CrossRef]
- Schirmer, M.; Luster, J.; Linde, N.; Perona, P.; Mitchell, E.; Andrew Barry, D.; Hollender, J.; Cirpka, O.; Schneider, P.; Vogt, T.; et al. Morphological, hydrological, biogeochemical and ecological changes and challenges in river restoration—The Thur River case study. Hydrol. Earth Syst. Sci. 2014, 18, 2449–2462. [Google Scholar] [CrossRef] [Green Version]
- Uehlinger, U. Resistance and resilience of ecosystem metabolism in a flood-prone river system. Freshw. Biol. 2000, 45, 319–332. [Google Scholar] [CrossRef]
- Wolman, M.G. A method of sampling coarse river-bed material. EOS Trans. Am. Geophys. Union 1954, 35, 951–956. [Google Scholar] [CrossRef]
- Tockner, K.; Malard, F.; Burgherr, P.; Robinson, C.T.; Uehlinger, U.; Zah, R.; Ward, J.V. Characteristics of channel types in a glacial floodplain ecosystem (Val Roseg, Switzerland). Arch. Hydrobiol. 1997, 140, 430–463. [Google Scholar]
- Blott, S.J.; Pye, K. GRADISTAT: A grain size distribution and statistics package for the analysis of unconsolidated sediments. Earth Surf. Process. Landf. 2001, 26, 1237–1248. [Google Scholar] [CrossRef]
- Baxter, C.V.; Hauer, F.R.; Woessner, W.W. Measuring groundwater-stream water exchange: New techniques for installing minipiezometers and estimating hydraulic conductivity. Trans. Am. Fish. Soc. 2003, 132, 493–502. [Google Scholar] [CrossRef]
- Uehlinger, U.; Naegeli, M.W.; Fisher, S.G. A heterotrophic desert stream? The role of sediment stability. West. N. Am. Nat. 2002, 62, 466–473. [Google Scholar]
- Doering, M.; Uehlinger, U.; Ackermann, T.; Woodtle, M.; Tockner, K. Spatiotemporal heterogeneity of soil and sediment respiration in a river-floodplain mosaic (Tagliamento, NE Italy). Freshw. Biol. 2011, 56, 1297–1311. [Google Scholar] [CrossRef]
- Naegeli, W.; Uehlinger, U. Contribution of the hyporheic zone to ecosystem metabolism in a prealpine gravel-bed river. J. N. Am. Benthol. Soc. 1997, 16, 794–804. [Google Scholar] [CrossRef]
- Bergey, E.A.; Getty, G.M. A review of methods for measuring the surface area of stream substrates. Hydrobiologia 2006, 556, 7–16. [Google Scholar] [CrossRef]
- Meyns, S.; Illi, R.; Ribi, B. Comparison of chlorophyll—A analysis by HPLC and spectrophotometry: Where do the differences come from? Arch. Hydrobiol. 1994, 132, 129–139. [Google Scholar]
- Vischer, D.; Hager, W.H.; Casanova, C.; Joos, B.; Lier, P.; Martini, O. Bypass tunnels to prevent reservoir sedimentation Q74 R37. In Proceedings of the 19th ICOLD Congress, Florence, Italy, 26–30 May 1997. [Google Scholar]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Muotka, T.; Paavola, R.; Haapala, A.; Novikmec, M.; Laasonen, P. Long-term recovery of stream habitat structure and benthic invertebrate communities from in-stream restoration. Biol. Conserv. 2002, 105, 243–253. [Google Scholar] [CrossRef]
- Lepori, F.; Palm, D.; Brännäs, E.; Malmqvist, B. Does restoration of structural heterogeneity in streams enhance fish and macroinvertebrate diversity? Ecol. Appl. 2005, 15, 2060–2071. [Google Scholar] [CrossRef]
- Jähnig, S.C.; Brabec, K.; Buffagni, A.; Erba, S.; Lorenz, A.W.; Ofenböck, T.; Verdonschot, P.F.M.; Hering, D. A comparative analysis of restoration measures and their effects on hydromorphology and benthic invertebrates in 26 central and southern European rivers. J. Appl. Ecol. 2010, 47, 671–680. [Google Scholar] [CrossRef]
- Palmer, M.A.; Menninger, H.L.; Bernhardt, E.S. River restoration, habitat heterogeneity and biodiversity: A failure of theory or practice? Freshw. Biol. 2010, 55, 205–222. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A. River restoration: The fuzzy logic of repairing reaches to reverse catchment scale degradation. Ecol. Appl. 2011, 21, 1926–1931. [Google Scholar] [CrossRef] [PubMed]
- Violin, C.R.; Cada, P.; Sudduth, E.B.; Hassett, B.A.; Penrose, D.L.; Bernhardt, E.S. Effects of urbanization and urban stream restoration on the physical and biological structure of stream ecosystems. Ecol. Appl. 2011, 21, 1932–1949. [Google Scholar] [CrossRef] [PubMed]
- Haase, P.; Hering, D.; Jähnig, S.C.; Lorenz, A.W.; Sundermann, A. The impact of hydromorphological restoration on river ecological status: A comparison of fish, benthic invertebrates, and macrophytes. Hydrobiologia 2012, 704, 475–488. [Google Scholar] [CrossRef]
- Robinson, C.T.; Uehlinger, U. Experimental floods cause ecosystem regime shift in a regulated river. Ecol. Appl. 2008, 18, 511–526. [Google Scholar] [CrossRef] [PubMed]
- Uehlinger, U.; Kawecka, B.; Robinson, C.T. Effects of experimental floods on periphyton and stream metabolism below a high dam in the Swiss Alps (River Spöl). Aquat. Sci. 2003, 65, 199–209. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Aristi, I.; Arroita, M.; Larrañaga, A.; Ponsatí, L.; Sabater, S.; von Schiller, D.; Elosegi, A.; Acuña, V. Flow regulation by dams affects ecosystem metabolism in Mediterranean rivers. Freshw. Biol. 2014, 59, 1816–1829. [Google Scholar] [CrossRef]
- Findlay, S. Importance of surface-subsurface exchange in stream ecosystems: The hyporheic zone. Limnol. Oceanogr. 1995, 40, 159–164. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Newbold, J.D.; Hendricks, S.P. A mixing model analysis of stream solute dynamics and the contribution of a hyporheic zone to ecosystem function. Freshw. Biol. 2003, 48, 995–1014. [Google Scholar] [CrossRef]
- Boulton, A.J.; Findlay, S.; Marmonier, P.; Stanley, E.H.; Valett, H.M. The functional significance of the hyporheic zone in streams and rivers. Annu. Rev. Ecol. Syst. 1998, 29, 59–81. [Google Scholar] [CrossRef]
- Fernald, A.G.; Landers, D.H.; Wigington, P.J. Water quality changes in hyporheic flow paths between a large gravel bed river and off-channel alcoves in Oregon, USA. River Res. Appl. 2006, 22, 1111–1124. [Google Scholar] [CrossRef]
- Poff, L.N.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stormberg, J.C. The natural flow regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Gomez-Velez, J.D.; Harvey, J.W. A hydrogeomorphic river network model predicts where and why hyporheic exchange is important in large basins. Geophys. Res. Lett. 2014, 41, 6403–6412. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flow Classes | ||||
|---|---|---|---|---|
| Indicator | High | Mod. High | Intermediate | Low |
| n | 4 | 4 | 2 | 2 |
| Mean (m3/s) | 76.4 ± 7.2 | 41.2 ± 3.9 | 32.7 ± 0.6 | 14.1 ± 0.2 |
| Median (m3/s) | 49.7 ± 7.8 | 32.4 ± 3.0 | 31.0 ± 0.4 | 12.8 ± 0.5 |
| Skewness | 1.6 ± 0.1 | 1.3 ± 0.0 | 1.1 ± 0.0 | 1.1 ± 0.0 |
| Max (m3/s) | 281.0 ± 15.5 | 175.8 ± 24.2 | 49.9 ± 1.7 | 35.5 ± 6.2 |
| Min (m3/s) | 23.5 ± 3.8 | 15.1 ± 2.0 | 23.7 ± 2.2 | 6.8 ± 0.5 |
| Days < first q | 1.8 ± 0.9 | 7.5 ± 3.0 | 0.0 ± 0.0 | 41.5 ± 8.1 |
| Days > third q | 15.5 ± 2.8 | 8.0 ± 2.3 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| CV | 0.9 ± 0.0 | 0.8 ± 0.1 | 0.2 ± 0.0 | 0.4 ± 0.1 |
| Days disrupt | 3.8 ± 1.0 | 1.0 ± 0.5 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Vel | Depth | Stream d50 | DOC | TN | NO3-N | TP | Cond | FPOM | CPOM | Hypo-d50 | VHG | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | m/s | cm | cm | mg C/L | mg N/L | mg N/L | µg P/L | µS/cm 20 °C | g/kg sed | g/kg sed | cm | cm/cm |
| 1 | 0.35 ± 0.1 | 37.8 ± 3.7 | 3.4 ± 0.3 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 37.5 ± 3.5 | 422.3 ± 13.3 | 4.0 ± 0.1 | 0.18 ± 0.02 | 4.8 ± 0.1 | 0.12 ± 0.06 |
| 2 | 0.55 ± 0.1 | 25.0 ± 2.2 | 4.2 ± 0.5 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 37.9 ± 3.8 | 423.8 ± 13.3 | 4.5 ± 0.2 | 0.10 ± 0.02 | 3.8 ± 0.2 | −0.06 ± 0.03 |
| 3 | 0.21 ± 0.1 | 27.5 ± 3.1 | 3.1 ± 0.3 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 38.3 ± 3.8 | 425.4 ± 13.1 | 4.0 ± 0.1 | 0.24 ± 0.05 | 3.2 ± 0.2 | 0.08 ± 0.03 |
| 4 | 0.27 ± 0.1 | 27.2 ± 2.7 | 1.9 ± 0.3 | 2.3 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 37.5 ± 3.8 | 423.4 ± 13.4 | 4.2 ± 0.1 | 0.16 ± 0.03 | 3.0 ± 0.2 | 0.04 ± 0.03 |
| 5 | 0.23 ± 0.1 | 18.3 ± 1.3 | 3.5 ± 0.3 | 2.2 ± 0.1 | 2.4 ± 0.1 | 2.2 ± 0.1 | 39.6 ± 3.8 | 423.9 ± 13.0 | 4.1 ± 0.1 | 0.15 ± 0.02 | 4.4 ± 0.2 | 0.22 ± 0.03 |
| Flow class | ||||||||||||
| High | 0.39 ± 0.1 | 27.1 ± 2.3 | 2.7 ± 0.2 | 1.9 ± 0.1 | 2.2 ± 0.1 | 2.1 ± 0.1 | 38.4 ± 1.8 | 421.8 ± 8.3 | 3.9 ± 0.1 | 0.14 ± 0.02 | 3.9 ± 0.2 | 0.11 ± 0.02 |
| Mod. high | n.a. | 31.2 ± 2.6 | 2.9 ± 0.2 | 2.4 ± 0.1 | 2.3 ± 0.1 | 2.2 ± 0.1 | 45.1 ± 2.3 | 421.1 ± 8.7 | 4.4 ± 0.1 | 0.15 ± 0.02 | 3.8 ± 0.2 | 0.04 ± 0.03 |
| Intermediate | n.a. | n.a. | n.a. | 2.6 ± 0.1 | 2.8 ± 0.2 | 2.5 ± 0.1 | 43.9 ± 2.8 | 421.0 ± 22.3 | 3.9 ± 0.1 | 0.10 ± 0.02 | 4.0 ± 0.2 | n.a. |
| Low | 0.22 ± 0.1 | 21.3 ± 1.7 | 4.2 ± 0.3 | 2.3 ± 0.1 | 2.5 ± 0.1 | 2.3 ± 0.1 | 18.2 ± 0.7 | 435.8 ± 12.4 | 4.6 ± 0.1 | 0.31 ± 0.05 | 3.6 ± 0.3 | 0.08 ± 0.04 |
| Site | Flow Class | Site * Flow Class | ||
|---|---|---|---|---|
| Abiotic factors | ||||
| Physical | Velocity | 0.07 | 0.04 | 0.36 |
| Depth | 0.71 | 0.71 | 0.5 | |
| Stream-d50 | <0.01 | <0.01 | 0.43 | |
| Chemical | DOC | 0.99 | <0.01 | 0.99 |
| TN | 0.99 | <0.01 | 1 | |
| NO3-N | 0.99 | 0.08 | 1 | |
| TP | 0.98 | <0.01 | 0.99 | |
| Cond | 0.99 | 0.86 | 1 | |
| Hyporheic | FPOM | 0.01 | <0.01 | 0.91 |
| CPOM | 0.01 | <0.01 | <0.01 | |
| Hypo-d50 | <0.01 | 0.55 | <0.01 | |
| VHG | <0.01 | 0.06 | 0.03 | |
| Ecosystem function | ||||
| SR | <0.01 | <0.01 | <0.01 | |
| Periphyton biomass | 0.93 | <0.01 | 0.49 | |
| Chl-a | 0.39 | <0.01 | 0.82 | |
| Ecosystem structure | ||||
| Density | <0.01 | <0.01 | 0.32 | |
| Richness | <0.01 | <0.01 | 0.02 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín, E.J.; Ryo, M.; Doering, M.; Robinson, C.T. Evaluation of Restoration and Flow Interactions on River Structure and Function: Channel Widening of the Thur River, Switzerland. Water 2018, 10, 439. https://doi.org/10.3390/w10040439
Martín EJ, Ryo M, Doering M, Robinson CT. Evaluation of Restoration and Flow Interactions on River Structure and Function: Channel Widening of the Thur River, Switzerland. Water. 2018; 10(4):439. https://doi.org/10.3390/w10040439
Chicago/Turabian StyleMartín, Eduardo J., Masahiro Ryo, Michael Doering, and Christopher T. Robinson. 2018. "Evaluation of Restoration and Flow Interactions on River Structure and Function: Channel Widening of the Thur River, Switzerland" Water 10, no. 4: 439. https://doi.org/10.3390/w10040439
APA StyleMartín, E. J., Ryo, M., Doering, M., & Robinson, C. T. (2018). Evaluation of Restoration and Flow Interactions on River Structure and Function: Channel Widening of the Thur River, Switzerland. Water, 10(4), 439. https://doi.org/10.3390/w10040439