
Comparative Inactivation of the RNA of the Delta and Omicron Variants of SARS-CoV-2 in Wastewater of Five Municipalities in Southeast Germany


Institute of Medical Microbiology and Virology, University Hospital Carl Gustav Carus, Technische Universität Dresden, Fetscherstrasse 74, 01307 Dresden, Germany
*
Author to whom correspondence should be addressed.
Water 2024, 16(8), 1193; https://doi.org/10.3390/w16081193
Submission received: 25 March 2024
/
Revised: 16 April 2024
/
Accepted: 18 April 2024
/
Published: 22 April 2024
(This article belongs to the Section Water Quality and Contamination)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:During the COVID-19 pandemic, wastewater-based epidemiology has been extensively used as a helpful tool for evaluation of the epidemic situation in catchments of wastewater treatment plants. In this context, knowledge about the rate of virus inactivation in wastewater is important for characterization of the influence of retention times in sewers on virus concentrations, of the storage periods before analysis in the laboratory, on virus elimination during treatment and for modeling approaches. In the present study, we genotyped SARS-CoV-2 in the respiratory tract specimens of infected patients and spiked the raw wastewater of five treatment plants with aliquots of these samples. The test preparations were incubated under practically relevant temperature conditions (14 °C) over 31 days. The linear decay rates resulted in comparable T90 values (30.5 d) for the Delta and the Omicron BA1 variants of SARS-CoV-2 without differences in using the E or S gene of the virus as the target for amplification. In contrast, the origin of wastewater influenced the inactivation rate of both variants significantly, with the mean T90 values varying between 24.3 and 53.1 days in the wastewater from the five plants. The data suggest that the inactivation rate of SARS-CoV-2 in wastewater is more strongly determined by the composition of the water than by the virus variant.
1. Introduction
The quantitative and qualitative detection of SARS-CoV-2 in the influents of wastewater treatment plants (WWTPs) is an excellent example to demonstrate the value of wastewater-based epidemiology (WBE) as an additional approach to monitoring the circulation of infectious diseases in the human community [1]. Beyond WWTPs, wastewater from institutions housing vulnerable persons (e.g., retirement homes, hospitals) can be monitored. In contrast to clinical approaches, WBE is independent of the kind and extent of the test procedures in the population and includes infected patients with mild symptoms. Based on the fecal excretion of viruses by many infected patients [2], numerous studies have confirmed the suitability of SARS-CoV-2 detection in wastewater for following or even predicting the incidence of infections in the population served by the WWTP [3,4,5]. Recently, the research on wastewater surveillance of SARS-CoV-2 has been focused on optimizing the methods of virus concentration and virus detection and the normalization of data [6,7,8,9]. In contrast, relatively little is known about the persistence of viruses in wastewater. Understanding their inactivation processes and determining virus stability is not only important for risk and modeling approaches but also for evaluation of the influence of the time between the excretion of and the arrival of feces at a WWTP, of the transport and storage of samples before analysis on the virus concentrations detected in the laboratory [10] and of virus removal in wastewater treatment processes [11].
In recent years, the evolution of SARS-CoV-2 after countless infections of immunized and non-immunized persons worldwide has resulted in the emergence of variants that have temporarily dominated in human populations [12]. Whereas the follow-up of clinically common variants in wastewater has been documented in many reports [13], data on the differences between variants regarding their quantitative potential for fecal excretion are not available. In addition, studies investigating the differences in the environmental stability of SARS-CoV-2 variants are rare. Inactivation experiments on surfaces have demonstrated conflicting results relating to the persistence of SARS-CoV-2 variants [14,15,16,17,18,19,20]. To our knowledge, comparative investigations of the decay rates of variant strains in wastewater do not exist, and the genotype of the virus is often not specified in many reports.
Viruses in aquatic environments are exposed to many inactivating factors. Whereas particular mechanisms of inactivation can be neglected in sewers (e.g., UV light), mainly temperature and the activity of accompanying organisms (bacteria, protozoa) have been shown as important for virus persistence [21]. Furthermore, sewer biofilms have an influence on RNA stability and distribution [22,23]. Due to different temperatures of incubation, comparison of the results of previous SARS-CoV-2 persistence studies is difficult. In most cases, 4 °C and 20/25 °C have been, used which are only partially comparable to the conditions in the sewers of countries with a temperate climate. In long-term experiments (at least 30 days), the time to achieve a 90% reduction (one log) in SARS-CoV-2 (T90) in untreated wastewater ranged between 2.8 and 52 days (4 °C), while it took approximately 13 days at 20 °C, respectively [24,25]. It is important to note that different spike material was added to the wastewater (gamma-irradiated virus after propagation in cell culture, endogenous virus, aliquots of clinical samples), which might have influenced the decay rates in different reports. In addition, the results of infectivity tests have confirmed that, in comparison to RNA, the T90 of infectious viruses was strongly reduced to values between 10.4 h and 5.5 days (4–24 °C), probably depending on the virus titer [26,27,28]. Whereas the persistence of SARS-CoV-2 in generally different water matrices (like wastewater, autoclaved wastewater, treated wastewater, river and seawater, tap water) has been investigated in several studies [24,27,29], data on virus stability in raw wastewater from different locations are limited.
Based on the current gaps in our understanding of the inactivation of SARS-CoV-2, the aim of the present study was to assess the stability of the RNA of the Delta and Omicron variants in wastewaters from five treatment plants in Germany. All the assays were performed at 14 °C to simulate approximately the actual temperature of wastewater in the local sewers [30] and were repeated to investigate the variability in the virus decay in the influents of all the plants included in the study. The use of two targets in the genome of SARS-CoV-2 for amplification allowed us to draw conclusions about the influence of the real-time quantitative PCR method on the results, which was found to be an important aspect of virus persistence data in previous studies [25,31,32]. Parallel detection of crAssphages can enable a comparison of the decay rates of SARS-CoV-2, and they are a common indicator of fecal pollution in water [33].
2. Materials and Methods
2.1. Wastewater Samples
Between October 2022 and May 2023, composite (24 h) untreated wastewater samples were collected from five WWTPs in Saxony, Germany, receiving wastewater from different population sizes. The main characteristics of the plants and sewer systems are summarized in Table S1. The concentration of SARS-CoV-2 was determined (background) as reported [5], and the samples were stored at 4 °C and used for the inactivation experiments within 24 h.
2.2. Inactivation Experiments
Randomly selected SARS-CoV-2-positive throat swab fluids from patients with symptoms of respiratory tract infections were aliquoted immediately after testing and used for whole genome sequencing, as described recently [34], to determine the virus variant present in the samples. After aliquoting, the remaining samples were frozen (−80 °C) and were not further thawed until the start of the inactivation experiments. Each swab fluid was used only once. According to the measured Ct values in the real-time PCR, the throat samples were diluted with PBS (phosphate-buffered saline) to obtain comparable numbers of SARS-CoV-2 genome copies. The diluted samples (25 µL each) were used to spike the wastewater (4 mL, plastic tubes), which was incubated using over-head shaking (60×/min) at 14 °C in dark in a cooled incubator. During an incubation period of four weeks, each wastewater was sampled six times on the same days (0, 3, 6, 12, 24 and 31).
2.3. RNA Extraction and Virus Detection and Quantification
The nucleic acids of the viruses in the wastewater samples (200 µL each) were prepared using RNeasy columns as recommended by the manufacturer (QIAGEN) and treated to remove the PCR inhibitors (Zymo Research, Irvine, CA, USA). After storage at 4 °C, detection of SARS-CoV-2 using real-time quantitative PCR (RTqPCR) was performed within 3 h after sampling, as reported [5]. Briefly, a commercial kit (Altona, Hamburg, Germany) amplifying the E and S genes of SARS-CoV-2 (RealStar SARS-CoV-2 RT-PCR kit 1.0) was used as recommended by the manufacturer. The amplifications were carried out in triplicate using a QuantStudio5 thermocycler. Positive and negative controls were included in all the runs. Samples were considered positive if amplification was positive in at least two of the three replicates with Ct values ≤ 40. The virus concentrations were calculated with the standard curve of a commercial RNA standard of SARS-CoV-2 (Wuhan strain; Twist Bioscience, San Francisco, CA, USA). Detection and quantification of the naturally occurring crAssphages in WWTP 1 were performed as described recently [35]. All the steps of sample processing and virus detection were strictly separated in different laboratories.
2.4. Inactivation Rate, Calculation of T90 and Statistical Analysis
The measured mean concentrations of the genome copies (E- or S-gene-based) were linearized using logarithmic transformation, and the time for a one log reduction (90%) in the SARS-CoV-2-specific RNA concentration (T90) was calculated as described [36]. The goodness of fit of the linear regression was characterized by the coefficient of determination (r2). A paired t-test was used to compare the mean T90 values obtained from different experimental runs (variables: virus variants, E and S gene, origin of wastewater, SARS-CoV-2 and crAssphages), considering α < 0.05 as significant.
3. Results
The maximal background of SARS-CoV-2 in all the wastewaters used in the inactivation experiments was below 0.2% of the concentration in the spiked samples and thus can be neglected for evaluation of the decay rates of the specified virus variants used for spiking. Overall, 38 independent stability experiments with the wastewaters of five WWTPs were performed in this study (19 for each Delta and Omicron variant, Table S2). The virus concentrations at the start of the inactivation experiments (day 0) varied between 4.6 × 105 to 8.8 × 106/mL (Delta variant) and 2.7 × 105 to 7.2 × 106/mL (Omicron variant), respectively. Linearization of the time-dependent concentrations of SARS-CoV-2 resulted in a mean coefficient of determination of 0.924 ± 0.062 (range: 0.712 to 0.996) and first-order decay rate constants between 0.024 and 0.148 (mean of experiments with the Delta variant: 0.089 ± 0.029; Omicron variant: 0.084 ± 0.024), respectively (Table S2). Based on the averaged constants from all the experiments, mean T90 values of 30.5 ± 17.6 (Delta variant) and 30.5 ± 11.5 days (Omicron variant) were calculated (Figure 1), demonstrating a comparable persistence of both variants in the wastewater. However, the relatively high standard deviations of the T90 values indicate a broad range of stability data at the same temperature. One reason for this result might be a locally and time-dependent variable influence of the substances in the five wastewater sources used in the study. For WWTPs 1–5 under comparable experimental conditions (the same aliquots of a clinical sample added to the water of all the WWTPs), mean T90 values of 26.1, 24.3, 53.1, 36.6 and 26.1 days were measured (Figure 2), showing statistically significant differences in some cases (e.g., WWTP 3). These differences are independent of the SARS-CoV-2 variant used to spike the aliquots of the same wastewater sample (Figure 3). The variant-specific decay and its variability at any treatment plant corresponded largely to the averaged T90 values in the wastewater of the different WWTPs (Figure 2). Parallel detection of SARS-CoV-2 and crAssphages was performed in some of the samples from treatment plant 1. The decay kinetics resulted in mean T90 values of 21.1 ± 2.6 (SARS-CoV-2) and 61.9 ± 14.9 days (crAssphages), demonstrating a statistically significant difference in the persistence of both viruses (Figure 4). After the use of the E and S genes as targets for amplification in the investigated wastewater samples, slightly lower T90 values for the S gene (30.5 ± 14.7 vs. 28.6 ± 14.1 days without statistical significance, n = 38) and the same differences between the samples of the WWTPs as described were found. This result was independent of the virus variant.
4. Discussion
In principle, the use of fresh SARS-CoV-2-positive stool samples can be considered the best raw material to use to spike wastewater for inactivation experiments. In the present study, SARS-CoV-2 from the clinical material of infected patients was used to calculate the decay of viral RNA in the wastewater. It cannot be excluded that the single freezing/thawing of the swab fluids might have had an influence on the integrity of the structures of the virus surface in comparison with particles obtained immediately after propagation in cell cultures. This aspect is equally important for comparison with the virus persistence rates measured in different studies. In general, the loss of infectivity of viruses is faster than the degradation of RNA. As an example, the strain hCoV-19/USA-WA1/2020, NR-52281 (lineage A) was used to spike wastewater samples, and a decay of infectivity (T90) of 10.8 h was determined [28]. It is of note that the virus aliquots were also frozen before use in these inactivation experiments. Furthermore, the results of the cultivation assays confirmed that the rate of infectious viruses in stool samples of COVID-19 patients is relatively low, and so far, only non-infectious virus particles have been detected in raw wastewater [4,37,38], indicating a partial degradation of viruses during the intestinal passage of SARS-CoV-2 and its transport to WWTPs. After using pasteurization, freeze–thaw and detergent treatments on viruses in wastewater, Robinson et al. [38] have suggested the presence of intact but non-infectious virus particles in wastewater. Thus, it can be assumed that the spike material in the present study is largely representative of the SARS-CoV-2 data measured in wastewater monitoring.
Here, statistically significant differences between the T90 values in the wastewater of the five WWTPs were demonstrated. A couple of in-sewer factors have been identified that may determine virus concentrations and, as a consequence, virus stability [39]. Among them, temperature is the most important parameter, but further, more locally specific factors, like the pH, chlorination or discharge of industrial wastewater, may play a role in virus inactivation processes. Due to the complex composition of wastewater and the variety of multifactorial interactions between viruses and the water matrix, it can be hypothesized that experimental differentiation between the influencing parameters is difficult to determine. Additionally, the time-dependent fluctuation in the concentrations of substances in wastewater further complicates their evaluation for virus stability. The relatively high standard deviation of the mean T90 values in WWTPs 3 and 4 (Figure 3 and Figure 4) could be discussed in this context. Comparable studies addressing this aspect of variation in SARS-CoV-2 persistence are rare. Using ddPCR for detection, Beattie et al. [40] added bovine coronavirus to the wastewater of three plants and found no differences in their concentrations in unpasteurized samples. In contrast, Roldan-Hernandez et al. [32] confirmed changes in T90 values by a factor of up to 6 in primary settled solids from two WWTPs, which was independent of the incubation temperature and virus species (SARS-CoV-2 and the non-enveloped pepper mild mottle virus (PMMoV)). Further studies are necessary to determine the practical range of virus inactivation rates in natural wastewaters from various catchments incubated at environmentally relevant temperatures.
With a mean T90 value of 30.5 days, the decay rate measured here is largely consistent with a previous study. Using amplification of SARS-CoV-2 RNA (not infectivity) and incubation at a comparable temperature (15 °C), a T90 of 20.4 days for gamma-irradiated SARS-CoV-2 (Beta variant) was determined [24]. Besides variable spiking material, the experimental design of other reports differs greatly in relation to temperature [25,41,42,43], the type of water [29], the performance of the infectivity assays [26,27,28,29,44], the duration of incubation [45] or the use of primary settled solids as a matrix in their stability experiments [32]. Additionally, the initial concentration of the virus copies at the start of the incubation experiments might be of importance to T90 values [36]. In the present report, we focused on spiked but otherwise unmodified wastewater samples incubated at a defined temperature typical for a moderate climate and detected changes in the concentration of SARS-CoV-2 RNA, as undertaken in monitoring programs. Despite an optimized quantification of E gene copies using triplicate amplification of the RNA in each sample, linearization of the decay resulted in a broad range of T90 values. This is in accordance with the variability in measured persistence in other studies [31,36] and demonstrates the fundamental difficulty of comparing virus stabilities between different experiments. According to the many factors determining the persistence process, it must be assumed that the calculated T90 values reflect the result of a combined influence of the specific conditions in an experimental approach on viral RNA and can only be conditionally transferred to the stability of the same virus/RNA in other wastewaters.
In the present study, a statistically significant difference (factor of 2.9 of mean T90 values) between the mean decay rate of SARS-CoV-2 and the crAssphages was found. Data on the differences in persistence between SARS-CoV-2 and indicator viruses are inconsistent. Ahmed et al. [24] confirmed comparable inactivation of SARS-CoV-2 and murine hepatitis virus (MHV) in various types of water. The origins of both viruses were different (gamma-irradiated stock of SARS-CoV-2 and pooled fecal samples of MHV-infected mice), which could have had an influence on decay. Here, we used endogenous crAssphages in their naturally occurring state in wastewater. After incubation at a temperature of 12 °C, Burnet et al. [31] measured T90 values between 5.7 and 7.0 days (depending on the target used for amplification) for SARS-CoV-2, whereas the concentration of the common fecal indicator PMMoV remained constant over a period of 27 days. In a further study, the relation between PMMoV and SARS-CoV-2 stability in settled solids of raw wastewater depends on the incubation temperature and the origin of the wastewater sample [32]. Similar results were found in river water and seawater, demonstrating stable concentrations of both virus species at 4 °C but higher T90 values for PMMoV at 20 °C [29]. Using non-spiked raw sewage from university dormitories, Li et al. [42] reported strongly different ratios of T90 values for SARS-CoV-2 and PMMoV. These findings indicate that the persistence of the viral indicators of fecal water contamination does not completely correspond to the inactivation rates of SARS-CoV-2 and requires further investigations.
The results of this study suggest that the typical retention time of wastewater in sewer systems (<24 h) does not significantly affect the concentration of excreted SARS-CoV-2 RNA measured at a WWTP. This is independent of the virus variant present in the sample. Furthermore, many reports have confirmed the prolonged stability of viruses (including coronavirus) at lower temperatures, such as 14 °C. If short-term storage of the samples cannot be avoided due to limited laboratory capacities and/or logistic problems, the combined effect of the retention time in the sewer and of refrigerated storage seems of limited importance to the significance of the results of SARS-CoV-2 wastewater monitoring [26,40]. Regarding the circulation of different SARS-CoV-2 variants in wastewater, the comparable stability of the RNA of the Delta and Omicron variants indicates the suitability of WBE for evaluating the epidemiological situation in the catchment of WWTPs, and it can be expected that reliable data during further virus evolutions could be collected. Studies investigating the comparable persistence of different variants in wastewater are very rare. In a recent report, Sherchan et al. [44] used an infectivity assay and found T90 values of 17.7 (Delta variant) and 15.3 h (Omicron variant) at room temperature. Despite the strong methodological difference (infectivity vs. RNA decay) in comparison to the present study, the similar stability of both variants was confirmed. However, future inactivation experiments under defined and practically relevant conditions will be helpful to characterize the persistence of newly developed variants with dominant occurrence in the human population.
5. Conclusions
After incubation at 14 °C, the RNA persistence of the Delta and Omicron variants of SARS-CoV-2 in untreated wastewater from different treatment plants was compared. In the same water samples, calculation of the T90 values of both variants as well as the use of the E or S gene as an amplification target resulted in comparable decay rates for both variants. In contrast, statistically significant differences in the RNA stability in water from different plants were found, demonstrating a remarkable influence of local/temporal changes in the composition of the wastewater matrix on virus persistence.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w16081193/s1. Table S1: Characteristics of wastewater treatment plants, Table S2: Details of SARS-CoV-2 inactivation experiments performed in the study.
Author Contributions
Both authors contributed to the study conception and design. The material preparation, data collection and analysis were performed by R.D. and M.G. The first draft of the manuscript was written by R.D. M.G. commented on all versions of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The study was partly supported by a grant from the Sächsisches Staatsministerium für Wissenschaft, Kultur und Tourismus (FKZ: 100535976).
Institutional Review Board Statement
The clinical specimens were used in accordance with the Declaration of Helsinki, and the investigations were approved by the Ethics Committee of Medical Faculty of TU Dresden (IRB00001473).
Informed Consent Statement
All the subjects gave their informed consent for inclusion and to publish this paper before they participated in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Keshaviah, A.; Diamond, M.B.; Wade, M.J.; Scarpino, S.V. Wastewater monitoring can anchor global disease surveillance systems. Lancet Glob. Health 2023, 11, e976–e981. [Google Scholar] [CrossRef]
- Zhou, J.Q.; Liu, G.X.; Huang, X.L.; Gan, H.T. The importance of fecal nucleic acid detection in patients with coronavirus disease (COVID-19): A systematic review and meta-analysis. J. Med. Virol. 2022, 94, 2317–2330. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Gwee, S.X.W.; Ng, J.Q.X.; Lau, N.; Koh, J.; Pang, J. Wastewater surveillance to infer COVID-19 transmission: A systematic review. Sci. Total Environ. 2022, 804, 150060. [Google Scholar] [CrossRef]
- Tiwari, A.; Lipponen, A.; Hokajärvi, A.M.; Luomala, O.; Sarekoski, A.; Rytkönen, A.; Österlund, P.; Al-Hello, H.; Juutinen, A.; Miettinen, I.T.; et al. Detection and quantification of SARS-CoV-2 RNA in wastewater influent in relation to reported COVID-19 incidence in Finland. Water Res. 2022, 215, 118220. [Google Scholar] [CrossRef]
- Helm, B.; Geissler, M.; Mayer, R.; Schubert, S.; Oertel, R.; Dumke, R.; Dalpke, A.; El-Armouche, A.; Renner, B.; Krebs, P. Regional and temporal differences in the relation between SARS-CoV-2 biomarkers in wastewater and estimated infection prevalence—Insights from long-term surveillance. Sci. Total Environ. 2023, 857, 159358. [Google Scholar] [CrossRef]
- Ahmed, W.; Smith, W.J.M.; Tiwari, A.; Bivins, A.; Simpson, S.L. Unveiling indicator, enteric, and respiratory viruses in aircraft lavatory wastewater using adsorption-extraction and Nanotrap Microbiome a Particles workflows. Sci. Total Environ. 2023, 896, 165007. [Google Scholar] [CrossRef] [PubMed]
- Akter, J.; Smith, W.J.M.; Liu, Y.; Kim, I.; Simpson, S.L.; Thai, P.; Korajkic, A.; Ahmed, W. Comparison of adsorption-extraction (AE) workflows for improved measurements of viral and bacterial nucleic acid in untreated wastewater. Sci. Total Environ. 2024, 908, 167966. [Google Scholar] [CrossRef] [PubMed]
- Atoui, A.; Cordevant, C.; Chesnot, T.; Gassilloud, B. SARS-CoV-2 in the environment: Contamination routes, detection methods, persistence and removal in wastewater treatment plants. Sci. Total Environ. 2023, 881, 163453. [Google Scholar] [CrossRef]
- Mazumder, P.; Dash, S.; Honda, R.; Sonne, C.; Kumar, M. Sewage surveillance for SARS-CoV-2: Molecular detection, quantification, and normalization factors. Curr. Opin. Environ. Sci. Health 2022, 28, 100363. [Google Scholar] [CrossRef]
- Babler, K.M.; Sharkey, M.E.; Abelson, S.; Amirali, A.; Benitez, A.; Cosculluela, G.A.; Grills, G.S.; Kumar, N.; Laine, J.; Lamar, W.; et al. Degradation rates influence the ability of composite samples to represent 24-hourly means of SARS-CoV-2 and other microbiological target measures in wastewater. Sci. Total Environ. 2023, 867, 161423. [Google Scholar] [CrossRef]
- Foladori, P.; Cutrupi, F.; Cadonna, M.; Manara, S. Coronaviruses and SARS-CoV-2 in sewerage and their removal: Step by step in wastewater treatment plants. Environ. Res. 2022, 207, 112204. [Google Scholar] [CrossRef]
- Volz, E. Fitness, growth and transmissibility of SARS-CoV-2 genetic variants. Nat. Rev. Genet. 2023, 24, 724–734. [Google Scholar] [CrossRef]
- Itarte, M.; Bofill-Mas, S.; Martinez-Puchol, S.; Torrell, H.; Cereto, A.; Carrasco, M.; Fores, E.; Canela, N.; Girones, R.; Rusinol, M. Looking for a needle in a haystack. SARS-CoV-2 variant characterization in sewage. Curr. Opin. Environ. Sci. Health 2021, 24, 100308. [Google Scholar] [CrossRef]
- Bobrin, V.A.; Chen, S.R.; Grandes Reyes, C.F.; Smith, T.; Purcell, D.F.J.; Armstrong, J.; McAuley, J.L.; Monteiro, M.J. Surface inactivation of highly mutated SARS-CoV-2 Variants of Concern: Alpha, Delta, and Omicron. Biomacromolecules 2022, 23, 3960–3967. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.W.H.; Lai, A.M.Y.; Peiris, M.; Man Poon, L.L. Increased stability of SARS-CoV-2 Omicron variant over ancestral strain. Emerg. Infect. Dis. 2022, 28, 1515–1517. [Google Scholar] [CrossRef]
- Hirose, R.; Itoh, Y.; Ikegaya, H.; Miyazaki, H.; Watanabe, N.; Yoshida, T.; Bandou, R.; Daidoji, T.; Nakaya, T. Differences in environmental stability among SARS-CoV-2 variants of concern: Both omicron BA.1 and BA.2 have higher stability. Clin. Microbiol. Infect. 2022, 28, 1486–1491. [Google Scholar] [CrossRef] [PubMed]
- Meister, T.L.; Fortmann, J.; Todt, D.; Heinen, N.; Ludwig, A.; Brüggemann, Y.; Elsner, C.; Dittmer, U.; Steinmann, J.; Pfaender, S.; et al. Comparable environmental stability and disinfection profiles of the currently circulating SARS-CoV-2 Variants of Concern B.1.1.7 and B.1.351. J. Infect. Dis. 2021, 224, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Onianwa, O.; Garratt, I.; Carter, J.; Spencer, A.; Verlander, N.Q.; Pottage, T.; Bennett, A.M. Comparison of surface persistence of SARS-CoV-2 Alpha and Delta variants on stainless steel at 4 °C and 24 °C. Appl. Environ. Microbiol. 2022, 88, e0076422. [Google Scholar] [CrossRef]
- Pottage, T.; Onianwa, O.; Atkinson, B.; Spencer, A.; Bennett, A.M. Stability of SARS-CoV-2 variants of concern (Delta and Omicron) on surfaces at room temperature. Virology 2023, 583, 27–28. [Google Scholar] [CrossRef]
- Takeda, Y.; Nikaido, M.; Jamsransuren, D.; Matsuda, S.; Ogawa, H. Virucidal activities of acidic electrolyzed water solutions with different pH values against multiple strains of SARS-CoV-2. Appl. Environ. Microbiol. 2023, 89, e0169922. [Google Scholar] [CrossRef]
- Pinon, A.; Vialette, M. Survival of viruses in water. Intervirology 2018, 61, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ahmed, W.; Metcalfe, S.; Smith, W.J.M.; Choi, P.M.; Jackson, G.; Cen, X.; Zheng, M.; Simpson, S.L.; Thomas, K.V.; et al. Impact of sewer biofilms on fate of SARS-CoV-2 RNA and wastewater surveillance. Nat. Water 2023, 1, 272–280. [Google Scholar] [CrossRef]
- Zhang, S.; Sharma, E.; Tiwari, A.; Chen, Y.; Sherchan, S.P.; Gao, S.; Zhou, X.; Shi, J.; Jiang, G. The reduction of SARS-CoV-2 RNA concentration in the presence of sewer biofilms. Water 2023, 15, 2132. [Google Scholar] [CrossRef]
- Ahmed, W.; Bertsch, P.M.; Bibby, K.; Haramoto, E.; Hewitt, J.; Huygens, F.; Gyawali, P.; Korajkic, A.; Riddell, S.; Sherchan, S.P.; et al. Decay of SARS-CoV-2 and surrogate murine hepatitis virus RNA in untreated wastewater to inform application in wastewater-based epidemiology. Environ. Res. 2020, 191, 110092. [Google Scholar] [CrossRef] [PubMed]
- Hokajärvi, A.M.; Rytkönen, A.; Tiwari, A.; Kauppinen, A.; Oikarinen, S.; Lehto, K.M.; Kankaanpää, A.; Gunnar, T.; Al-Hello, H.; Blomqvist, S.; et al. The detection and stability of the SARS-CoV-2 RNA biomarkers in wastewater influent in Helsinki, Finland. Sci. Total Environ. 2021, 770, 145274. [Google Scholar] [CrossRef] [PubMed]
- Bivins, A.; Greaves, J.; Fischer, R.; Yinda, K.C.; Ahmed, W.; Kitajima, M.; Munster, V.J.; Bibby, K. Persistence of SARS-CoV-2 in water and wastewater. Environ. Sci. Technol. Lett. 2020, 7, 937–942. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, L.C.; Torres-Franco, A.F.; Lopes, B.C.; Santos, B.S.A.D.S.; Costa, E.A.; Costa, M.S.; Reis, M.T.P.; Melo, M.C.; Polizzi, R.B.; Teixeira, M.M.; et al. Viability of SARS-CoV-2 in river water and wastewater at different temperatures and solids content. Water Res. 2021, 195, 117002. [Google Scholar] [CrossRef] [PubMed]
- Sherchan, S.; Thakali, O.; Ikner, L.A.; Gerba, C.P. Survival of SARS-CoV-2 in wastewater. Sci. Total Environ. 2023, 882, 163049. [Google Scholar] [CrossRef] [PubMed]
- Sala-Comorera, L.; Reynolds, L.J.; Martin, N.A.; O’Sullivan, J.J.; Meijer, W.G.; Fletcher, N.F. Decay of infectious SARS-CoV-2 and surrogates in aquatic environments. Water Res. 2021, 201, 117090. [Google Scholar] [CrossRef]
- Figueroa, A.; Hadengue, B.; Leitao, J.P.; Blumensaat, F. A framework for modelling in-sewer thermal-hydraulic dynamic anomalies driven by stormwater runoff and seasonal effects. Water Res. 2023, 229, 119492. [Google Scholar] [CrossRef]
- Burnet, J.B.; Cauchie, H.M.; Walczak, C.; Goeders, N.; Ogorzaly, L. Persistence of endogenous RNA biomarkers of SARS-CoV-2 and PMMoV in raw wastewater: Impact of temperature and implications for wastewater-based epidemiology. Sci. Total Environ. 2023, 857, 159401. [Google Scholar] [CrossRef] [PubMed]
- Roldan-Hernandez, L.; Graham, K.E.; Duong, D.; Boehm, A.B. Persistence of endogenous SARS-CoV-2 and pepper mild mottle virus RNA in wastewater-settled solids. ACS ES&T Water 2022, 2, 1944–1952. [Google Scholar]
- Sabar, M.A.; Honda, R.; Haramoto, E. CrAssphage as an indicator of human-fecal contamination in water environment and virus reduction in wastewater treatment. Water Res. 2022, 221, 118827. [Google Scholar] [CrossRef]
- Stadtmüller, M.; Laubner, A.; Rost, F.; Winkler, S.; Patrasova, E.; Simunkova, L.; Reinhardt, S.; Beil, J.; Dalpke, A.H.; Yi, B. Emergence and spread of a sub-lineage of SARS-CoV-2 Alpha variant B.1.1.7 in Europe, and with further evolution of spike mutation accumulations shared with the Beta and Gamma variants. Virus Evol. 2022, 8, veac010. [Google Scholar] [CrossRef]
- Dumke, R.; Geissler, M.; Skupin, A.; Helm, B.; Mayer, R.; Schubert, S.; Oertel, R.; Renner, B.; Dalpke, A. Simultaneous detection of SARS-CoV-2 and influenza virus in wastewater of two cities in southeastern Germany, January to May 2022. Int. J. Environ. Res. Public Health 2022, 19, 13374. [Google Scholar] [CrossRef] [PubMed]
- Mahlknecht, J. Presence and persistence of SARS-CoV-2 in aquatic environments: A mini-review. Curr. Opin. Environ. Sci. Health 2022, 29, 100385. [Google Scholar] [CrossRef]
- Albert, S.; Ruiz, A.; Peman, J.; Salavert, M.; Domingo-Calap, P. Lack of evidence for infectious SARS-CoV-2 in feces and sewage. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2665–2667. [Google Scholar] [CrossRef]
- Robinson, C.A.; Hsieh, H.Y.; Hsu, S.Y.; Wang, Y.; Salcedo, B.T.; Belenchia, A.; Klutts, J.; Zemmer, S.; Reynolds, M.; Semkiw, E.; et al. Defining biological and biophysical properties of SARS-CoV-2 genetic material in wastewater. Sci. Total Environ. 2022, 807, 150786. [Google Scholar] [CrossRef]
- Bertels, X.; Demeyer, P.; van den Bogaert, S.; Boogaerts, T.; van Nuijs, A.L.N.; Delputte, P.; Lahousse, L. Factors influencing SARS-CoV-2 RNA concentrations in wastewater up to the sampling stage: A systematic review. Sci. Total Environ. 2022, 820, 153290. [Google Scholar] [CrossRef]
- Beattie, R.E.; Blackwood, A.D.; Clerkin, T.; Dinga, C.; Noble, R.T. Evaluating the impact of sample storage, handling, and technical ability on the decay and recovery of SARS-CoV-2 in wastewater. PLoS ONE 2022, 17, e0270659. [Google Scholar] [CrossRef]
- Islam, G.; Gedge, A.; Lara-Jacobo, L.; Kirkwood, A.; Simmons, D.; Desaulniers, J.P. Pasteurization, storage conditions and viral concentration methods influence RT-qPCR detection of SARS-CoV-2 RNA in wastewater. Sci. Total Environ. 2022, 821, 153228. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ash, K.T.; Joyner, D.C.; Williams, D.E.; Alamilla, I.; McKay, P.J.; Iler, C.; Green, B.M.; Kara-Murdoch, F.; Swift, C.M.; et al. Decay of enveloped SARS-CoV-2 and non-enveloped PMMoV RNA in raw sewage from university dormitories. Front. Microbiol. 2023, 14, 1144026. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Dong, Q.; Li, S.; Cheng, Z.; Kang, X.; Ren, D.; Xu, C.; Zhou, X.; Liang, P.; Sun, L.; et al. Persistence of SARS-CoV-2 RNA in wastewater after the end of the COVID-19 epidemics. J. Hazard. Mater. 2022, 429, 128358. [Google Scholar] [CrossRef] [PubMed]
- Sherchan, S.P.; Thakali, O.; Ikner, L.A.; Gerba, C.; Haramoto, E. Survivability of Delta and Omicron variants of SARS-CoV-2 in wastewater. Water Res. 2023, 246, 120644. [Google Scholar] [CrossRef]
- Weidhaas, J.; Aanderud, Z.T.; Roper, D.K.; VanDerslice, J.; Gaddis, E.B.; Ostermiller, J.; Hoffman, K.; Jamal, R.; Heck, P.; Zhang, Y.; et al. Correlation of SARS-CoV-2 RNA in wastewater with COVID-19 disease burden in sewersheds. Sci. Total Environ. 2021, 775, 145790. [Google Scholar] [CrossRef]
Figure 1.
Mean decay of SARS-CoV-2 variants Delta and Omicron in wastewater of five treatment plants (measured by amplification of E gene, mean of 19 individual experiments for each variant, 14 °C).
Figure 1.
Mean decay of SARS-CoV-2 variants Delta and Omicron in wastewater of five treatment plants (measured by amplification of E gene, mean of 19 individual experiments for each variant, 14 °C).

Figure 2.
Comparison of mean T90 values obtained from wastewater of treatment plants (WWTPs) 1–5 spiked with Delta or Omicron variant of SARS-CoV-2 (WWTP 1–4: n = 6, WWTP 5: n = 4). Data are results of virus detection through amplification of E gene. Dotted lines indicate statistically significant differences (α < 0.05).
Figure 2.
Comparison of mean T90 values obtained from wastewater of treatment plants (WWTPs) 1–5 spiked with Delta or Omicron variant of SARS-CoV-2 (WWTP 1–4: n = 6, WWTP 5: n = 4). Data are results of virus detection through amplification of E gene. Dotted lines indicate statistically significant differences (α < 0.05).
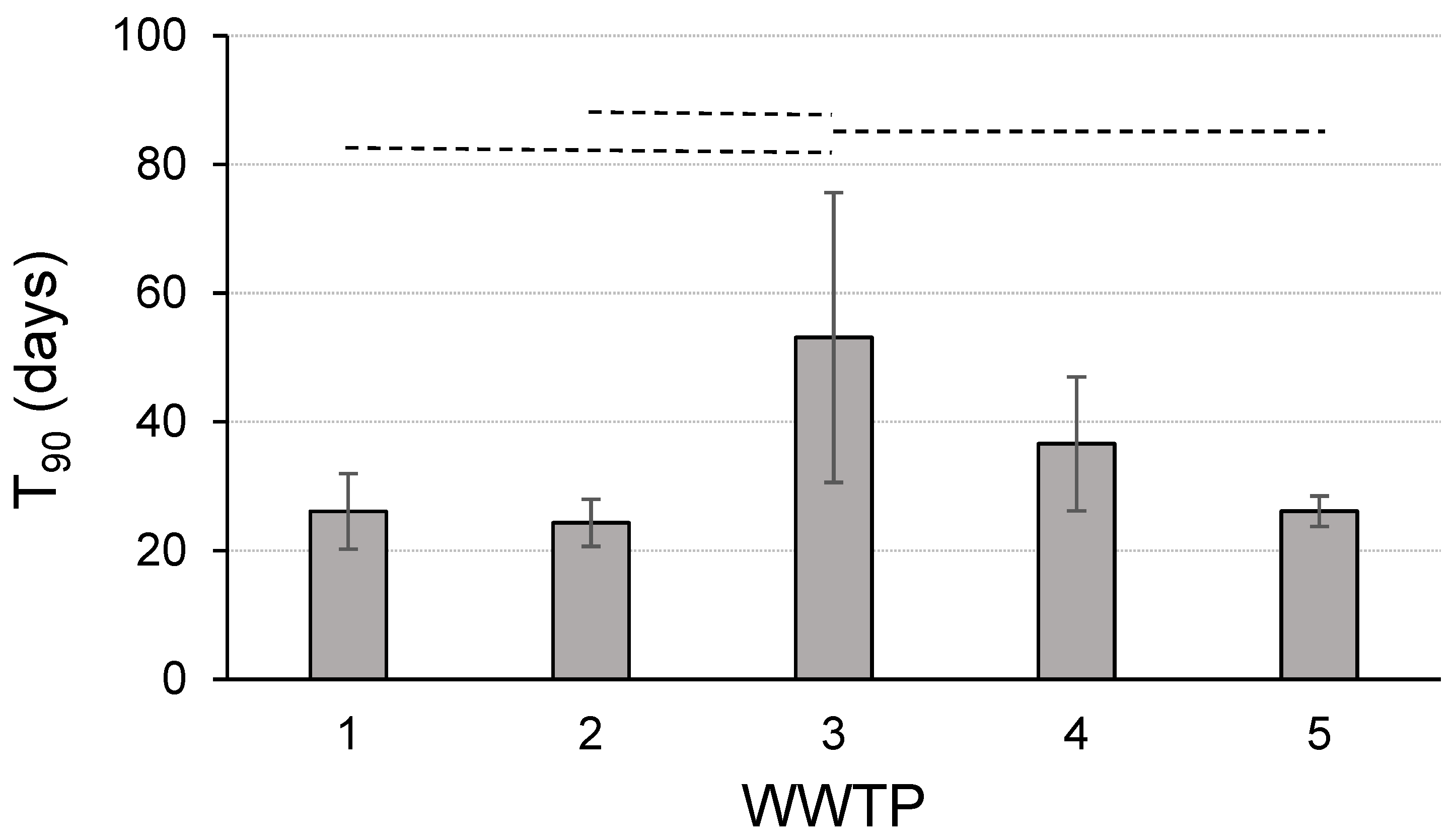
Figure 3.
Mean inactivation of Delta and Omicron variants of SARS-CoV-2 in wastewater of different treatment plants (WWTPs). In parentheses: number of independent experiments.
Figure 3.
Mean inactivation of Delta and Omicron variants of SARS-CoV-2 in wastewater of different treatment plants (WWTPs). In parentheses: number of independent experiments.
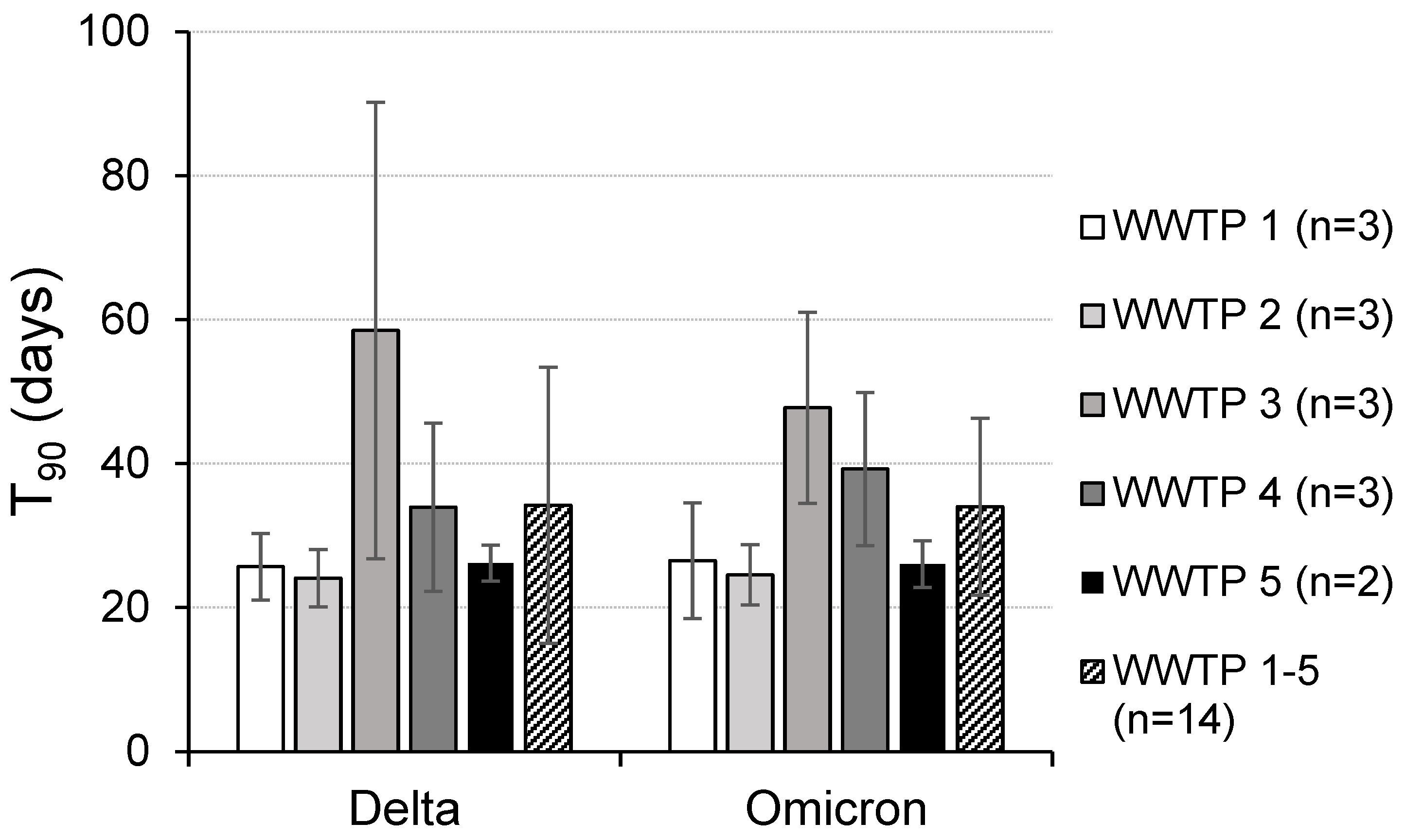
Figure 4.
Mean decay of SARS-CoV-2 (both variants) and crAssphages in wastewater of treatment plant 1 (n = 8).
Figure 4.
Mean decay of SARS-CoV-2 (both variants) and crAssphages in wastewater of treatment plant 1 (n = 8).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Geissler, M.; Dumke, R. Comparative Inactivation of the RNA of the Delta and Omicron Variants of SARS-CoV-2 in Wastewater of Five Municipalities in Southeast Germany. Water 2024, 16, 1193. https://doi.org/10.3390/w16081193
AMA Style
Geissler M, Dumke R. Comparative Inactivation of the RNA of the Delta and Omicron Variants of SARS-CoV-2 in Wastewater of Five Municipalities in Southeast Germany. Water. 2024; 16(8):1193. https://doi.org/10.3390/w16081193
Chicago/Turabian StyleGeissler, Michael, and Roger Dumke. 2024. "Comparative Inactivation of the RNA of the Delta and Omicron Variants of SARS-CoV-2 in Wastewater of Five Municipalities in Southeast Germany" Water 16, no. 8: 1193. https://doi.org/10.3390/w16081193
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.