Biogeochemical Cycle of Lanthanides in a Light Rare Earth Element-Enriched Geological Area (Quebec, Canada)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of the Study Area and Geological Description
2.2. Sampling Procedures
2.3. Physicochemical Extractions and Analyses
2.4. Data Analyses
3. Results and Discussion
3.1. REE Distribution in the Parent Material
3.2. Soil–Plant Distribution of REEs
3.3. Sediment–Water Interphase Distribution of REEs
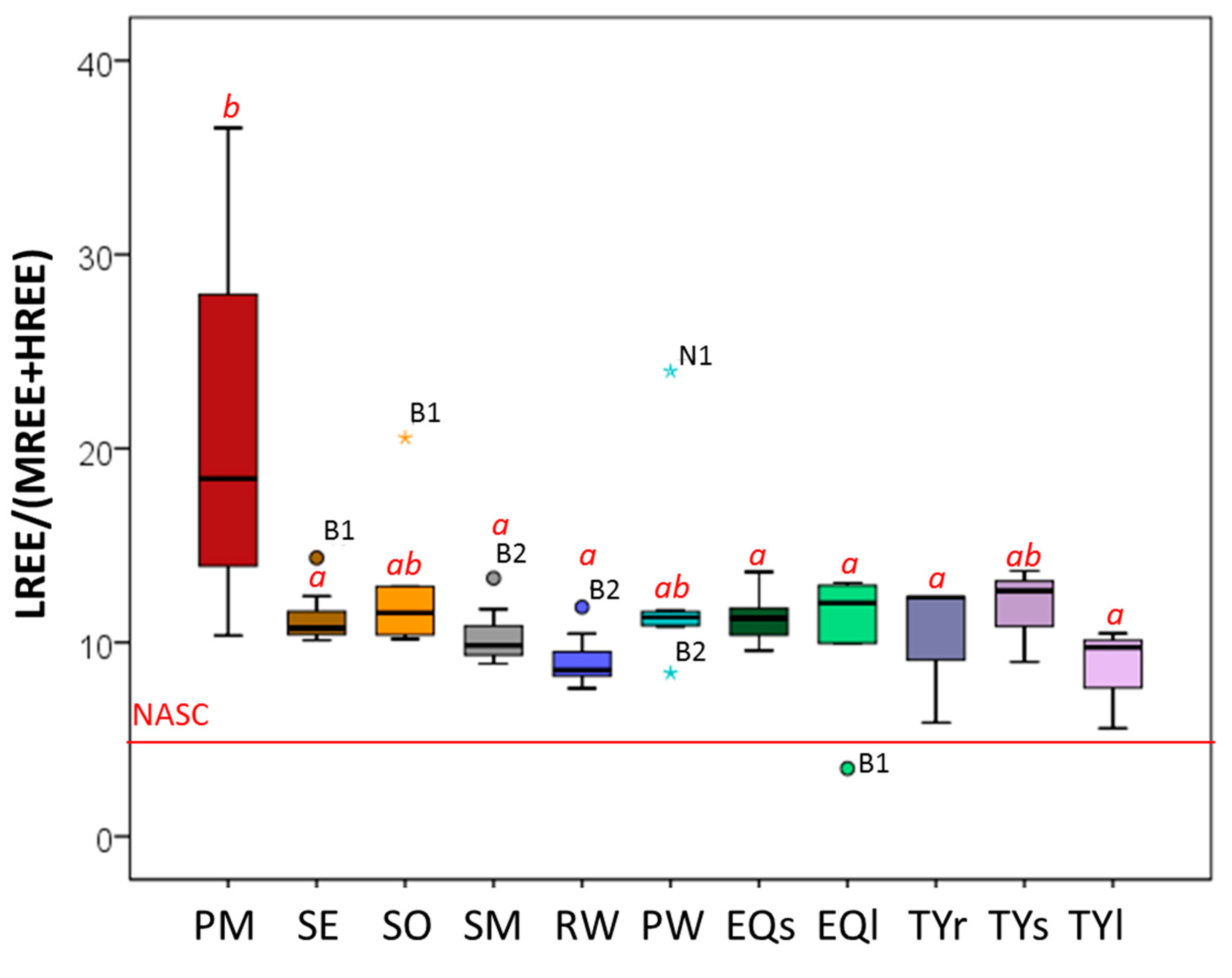
3.4. Distribution of REEs along the Study Ecosystem
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fisher, A.; Kara, D. Determination of rare earth elements in natural water samples—A review of sample separation, preconcentration and direct methodologies. Anal. Chimica. Acta 2016, 935, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhu, G.; Li, Y. Toxicological effects of rare earth yttrium on wheat seedlings (Triticum aestivum). J. Rare Earths 2013, 31, 1214–1220. [Google Scholar] [CrossRef]
- Turlin, F.; André-Mayer, A.-S.; Moukhsil, A.; Vanderhaeghe, O.; Gervais, F.; Solgadi, F.; Groulier, P.-A.; Poujol, M. Unusual LREE-rich, peraluminous, monazite- or allanite-bearing pegmatitic granite in the central Grenville Province, Québec. Ore Geol. Rev. 2017, 89, 627–667. [Google Scholar] [CrossRef]
- Chakhmouradian, A.R.; Wall, F. Rare earth elements: Minerals, mines, magnets (and more). Elements 2012, 8, 333–340. [Google Scholar] [CrossRef]
- Zepf, V. Rare Earth Elements: What and Where They Are. In Rare Earth Elements: A New Approach to the Nexus of Supply, Demand and Use: Exemplified along the Use of Neodymium in Permanent Magnets; Springer: Berlin/Heidelberg, Germany, 2013; pp. 11–39. [Google Scholar]
- Geagea, M.L.; Stille, P.; Millet, M.; Perrone, T. REE characteristics and Pb, Sr and Nd isotopic compositions of steel plant emissions. Sci. Total Environ. 2007, 373, 404–419. [Google Scholar] [CrossRef] [PubMed]
- Brioschi, L.; Steinmann, M.; Lucot, E.; Pierret, M.C.; Stille, P.; Prunier, J.; Badot, P.M. Transfer of rare earth elements (REE) from natural soil to plant systems: Implications for the environmental availability of anthropogenic REE. Plant Soil 2013, 366, 143–163. [Google Scholar] [CrossRef]
- Rare Earths Statistics and Information. U.S. Geological Survey, 2018. Available online: http://minerals.usgs.gov/minerals/pubs/commodity/rare_earths/ (accessed on 11 January 2019).
- Aubert, D.; Probst, A.; Stille, P. Distribution and origin of major and trace elements (particularly REE, U and Th) into labile and residual phases in an acid soil profile (Vosges Mountains, France). Appl. Geochem. 2004, 19, 899–916. [Google Scholar] [CrossRef] [Green Version]
- Öhlander, B.; Land, M.; Ingri, J.; Widerlund, A. Mobility of rare earth elements during weathering of till in northern Sweden. Appl. Geochem. 1996, 11, 93–99. [Google Scholar] [CrossRef]
- Chevis, D.A.; Johannesson, K.H.; Burdige, D.J.; Cable, J.E.; Martin, J.B.; Roy, M. Rare earth element cycling in a sandy subterranean estuary in Florida, USA. Mar. Chem. 2015, 176, 34–50. [Google Scholar] [CrossRef]
- Armand, R.; Cherubini, C.; Tuduri, J.; Pastore, N.; Pourret, O. Rare earth elements in French stream waters —Revisiting the geochemical continental cycle using FOREGS dataset. J. Geochem. Explor. 2015, 157, 132–142. [Google Scholar] [CrossRef]
- Amyot, M.; Clayden, M.G.; MacMillan, G.A.; Perron, T.; Arscott-Gauvin, A. Fate and trophic transfer of rare earth elements in temperate lake food webs. Environ. Sci. Technol. 2017, 51, 6009–6017. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, D.B.; Fairbrother, A. Examination of rare earth element concentration patterns in freshwater fish tissues. Chemosphere 2015, 120, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, O.; Cayer, A.; Pelletier, M.; Stevenson, R.; Jébrak, M. The Paleoproterozoic Montviel carbonatite-hosted REE-Nb deposit, Abitibi, Canada: Geology, mineralogy, geochemistry and genesis. Ore Geol. Rev. 2015, 67, 314–335. [Google Scholar] [CrossRef]
- Desharnais, G.; Duplessis, C. Montviel Core zone REE Mineral Resource Estimate Technical Report; SGS Canada Inc.: Blainville, QC, Canada, 2011; pp. 43–101. [Google Scholar]
- Barker, A.L. Rapport statutaire déposé au ministère des Ressources Naturelles et de la Faune, Québec. Summary Report on Exploration Work. 1975, p. 133. Available online: http://sigeom.mines.gouv.qc.ca (accessed on 17 April 2019).
- Corta, H.; Berthelot, P. Rapport D’une Campagne de Sondages. Propriété Montviel; Resources Nomans Inc.: Québec, QC, Canada, 2002. [Google Scholar]
- NI 43-101 Technical Report Montviel Rare Earth Project; GéoMégA Resources Inc.: Montreal, QC, Canada, 2015; p. 186.
- David, J.; Dion, C.; Goutier, J.; Roy, P.; Bandyayera, D.; Legault, M.; Rhéaume, P. Datations U-Pb effectuées dans la Sous-province de l’Abitibi à la suite des travaux de 2004–2005. Ministère des Ressources Naturelles et de la Faune, Québec 2006, 4, 22. [Google Scholar]
- Nadeau, O.; Stevenson, R.; Jébrak, M. Evolution of Montviel alkaline-carbonatite complex by coupled fractional crystallization, fluid mixing and metasomatism—Part I: Petrography and geochemistry of metasomatic aegirine-augite and biotite: Implications for REE-Nb mineralization. Ore Geol. Rev. 2016, 72, 1143–1162. [Google Scholar] [CrossRef]
- Nadeau, O.; Stevenson, R.; Jébrak, M. Evolution of Montviel alkaline-carbonatite complex by coupled fractional crystallization, fluid mixing and metasomatism—Part II: Trace element and Sm-Nd isotope geochemistry of metasomatic rocks: Implications for REE-Nb mineralization. Ore Geol. Rev. 2016, 72, 1163–1173. [Google Scholar] [CrossRef]
- Nadeau, O.; Stevenson, R.; Jébrak, M. Interaction of mantle magmas and fluids with crustal fluids at the 1894 Ma Montviel alkaline-carbonatite complex, Canada: Insights from metasomatic and hydrothermal carbonates. Lithos 2018, 296–299, 563–579. [Google Scholar] [CrossRef]
- Romero-Freire, A.; Minguez, L.; Pelletier, M.; Cayer, A.; Caillet, C.; Devin, S.; Gross, E.M.; Guérold, F.; Pain-Devin, S.; Vignati, D.A.L.; et al. Assessment of baseline ecotoxicity of sediments from a prospective mining area enriched in light rare earth elements. Sci. Total Environ. 2018, 61, 831–839. [Google Scholar] [CrossRef]
- McDonough, W.F.; Sun, S.-S. The composition of the Earth. Chem. Geol. 1995, 120, 223–253. [Google Scholar] [CrossRef]
- Gromet, L.P.; Dymek, R.F.; Haskin, L.A.; Korotev, R.L. The “North American Shale Composite” its compilation, major and trace element characteristic. Geochim. Cosmochim. Acta 1984, 48, 2469–2482. [Google Scholar] [CrossRef]
- Li, L.; Wu, J.; Lu, J.; Min, X.; Xu, J.; Yang, L. Distribution, pollution, bioaccumulation, and ecological risks of trace elements in soils of the northeastern Qinghai-Tibet Plateau. Ecotoxicol. Environ. Saf. 2018, 166, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Zaharescu, D.G.; Burghelea, C.I.; Dontsova, K.M.; Presler, J.K.; Maier, R.M.; Huxman, T.; Domanik, K.J.; Hunt, E.A.; Amistadi, M.K.; Gaddis, E.E.; et al. Ecosystem composition controls the fate of rare earth elements during incipient soil genesis. Sci. Rep. 2017, 7, 43208. [Google Scholar] [CrossRef] [PubMed]
- Edahbi, M.; Plante, B.; Benzaazoua, M. Environmental challenges and identification of the knowledge gaps associated with REE mine wastes management, J. Clean. Prod. 2019, 212, 1232–1241. [Google Scholar] [CrossRef]
- MacMillan, G.A.; Chételat, J.; Heath, J.; Mickpegak, R.; Amyot, M. Rare earth elements (REE) in freshwater, marine, and terrestrial ecosystems in the eastern Canadian Arctic Environ. Environ. Sci. Process. Impacts 2017, 19, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Morais, C.A.; Ciminelli, V.S.T. Recovery of europium by chemical reduction of a commercial solution of europium and Gadolinium chlorides. Hydrometallurgy 2001, 60, 247–253. [Google Scholar] [CrossRef]
- Soares Dinali, G.S.; Root, R.A.; Amistadi, M.K.; Chorover, J.; Lopes, G.; Guimarães Guilherme, L.R. Rare earth elements (REE) sorption on soils of contrasting mineralogy and texture. Environ. Int. 2019, 128, 279–291. [Google Scholar] [CrossRef]
- Liang, T.; Ding, S.; Song, W.; Chong, Z.; Zhang, C.; Li, H. A review of fractionations of rare earth elements in plants. J. Rare Earths 2008, 26, 7–15. [Google Scholar] [CrossRef]
- Ding, S.-M.; Liang, T.; Zhang, C.-S.; Wang, L.-J.; Sun, Q. Accumulation and fracitonation of rare earth elements in soil-wheat system. Pedosphere 2006, 16, 82–90. [Google Scholar] [CrossRef]
- Censi, P.; Cibella, F.; Falcone, E.E.; Cuttitta, G.; Saiano, F.; Inguaggiato, C.; Latteo, V. Rare earths and trace elements contents in leaves: A new indicator of the composition of atmospheric dust. Chemosphere 2017, 169, 342–350. [Google Scholar] [CrossRef]
- Brito, P.; Malvar, M.; Galinha, C.; Caçador, I.; Canário, J.; Araújo, M.F.; Raimundo, J. Yttrium and rare earth elements fractionation in salt marsh halophyte plants. Sci. Total Environ. 2018, 643, 1117–1126. [Google Scholar] [CrossRef]
- Casse, M.; Montero-Serrano, J.-C.; St-Onge, G.; Poirier, A. REE distribution and Nd isotope composition of estuarine waters and bulk sediment leachates tracing lithogenic inputs in eastern Canada. Mar. Chem. 2019, 211, 117–130. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Units | Sampling Site | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N0 | N1 | N2 | A1 | A2 | B1 | B2 | ||||
| SO | REE | mg kg−1 | 85.1 | 96.4 | - | 155.2 | 191.3 | 152.3 | - | |
| EQ | REE | Stem | mg kg−1 | 3.21 | 10.41 | - | 7.68 | 7.72 | 4.45 | - |
| REE | Leaf | mg kg−1 | 2.26 | 1.15 | - | 1.73 | 0.54 | 0.16 | - | |
| SO | La[N]/Yb[N] | 7.3 | 4.2 | 3.9 | 4.5 | 15.4 | ||||
| EQ | La[N]/Yb[N] | Stem | 15.0 | 11.1 | 3.6 | 6.0 | 6.0 | |||
| La[N]/Yb[N] | Leaf | 35.1 | 25.2 | 4.0 | 10.0 | 10.9 | ||||
| SO | Eu/Eu* | 1.16 | 0.95 | 0.91 | 1.13 | 1.16 | ||||
| EQ | Eu/Eu* | Stem | 4.93 | 2.24 | 1.25 | 1.48 | 2.57 | |||
| Eu/Eu* | Leaf | 5.32 | 6.60 | 1.25 | 3.23 | n.c. | ||||
| SO | Nd/Nd* | 1.45 | 1.53 | 1.38 | 1.59 | 1.24 | ||||
| EQ | Nd/Nd* | Stem | 1.28 | 1.25 | 1.20 | 1.22 | 1.25 | |||
| Nd/Nd* | Leaf | 1.18 | 1.22 | 1.05 | 1.12 | 1.28 | ||||
| EQ/SO | BAF | EQ stem/SO | - | 0.038 | 0.108 | - | 0.049 | 0.040 | 0.029 | - |
| BAF | EQ leaf/SO | - | 0.027 | 0.012 | - | 0.011 | 0.003 | 0.001 | - | |
| EQ | TF1 | EQ leaf/stem | - | 0.70 | 0.11 | - | 0.23 | 0.07 | 0.04 | - |
| Units | Sampling Site | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N0 | N1 | N2 | A1 | A2 | B1 | B2 | ||||
| SE | mg kg−1 | 183.6 | 181.0 | 101.2 | 184.9 | 110.5 | 89.2 | 71.3 | ||
| PW | µg L−1 | 99.4 | 13.2 | 49.8 | 44.3 | 25.8 | 14.0 | 20.8 | ||
| RW | µg L−1 | 4.0 | 4.1 | 3.7 | 3.5 | 4.2 | 3.0 | 4.3 | ||
| SM | REE | mg kg−1 | 1.3 | 5.7 | 4.2 | 1.4 | 1.4 | 0.4 | 0.3 | |
| TY | Root | mg kg−1 | 1.85 | - | 2.35 | |||||
| Stem | mg kg−1 | 0.39 | - | 1.20 | ||||||
| Leaf | mg kg−1 | 0.67 | 0.73 | 0.20 | ||||||
| SE | 3.67 | 3.27 | 3.78 | 3.35 | 5.93 | 6.43 | 4.28 | |||
| PW | 4.38 | 3.57 | 3.80 | 5.09 | 3.88 | 6.49 | 3.82 | |||
| RW | 2.10 | 2.49 | 3.17 | 2.11 | 2.40 | 3.77 | 4.80 | |||
| SM | La[N]/Yb[N] | 3.59 | 3.28 | 3.03 | 3.61 | 4.11 | 5.19 | 5.68 | ||
| TY | Root | 2.40 | 6.58 | |||||||
| Stem | 4.14 | 7.51 | ||||||||
| Leaf | 3.87 | 4.44 | 9.16 | |||||||
| SE | 0.96 | 1.04 | 0.92 | 0.89 | 1.04 | 1.14 | 0.97 | |||
| PW | 0.99 | 2.15 | 0.95 | 0.96 | 0.96 | 1.32 | 1.19 | |||
| RW | 1.13 | 0.94 | 0.97 | 1.04 | 1.02 | 1.77 | 1.71 | |||
| SM | Eu/Eu* | 0.98 | 0.97 | 0.95 | 1.14 | 0.95 | 1.88 | 1.34 | ||
| TY | Root | 1.20 | 2.95 | |||||||
| Stem | 3.20 | 3.41 | ||||||||
| Leaf | 3.98 | 1.62 | 26.16 | |||||||
| SE | 1.85 | 1.86 | 1.66 | 1.74 | 1.66 | 1.55 | 1.35 | |||
| PW | 1.22 | 10.74 | 0.65 | 0.39 | 1.12 | 0.24 | 0.19 | |||
| RW | 1.22 | 1.10 | 1.17 | 1.30 | 1.15 | 1.20 | 1.21 | |||
| SM | Nd/Nd* | 1.08 | 1.12 | 1.08 | 1.11 | 1.05 | 1.03 | 1.13 | ||
| TY | Root | 1.16 | 1.23 | |||||||
| Stem | 1.11 | 1.36 | ||||||||
| Leaf | 1.25 | 1.19 | 1.19 | |||||||
| TY/SE | TY root/SE | - | 0.018 | - | 0.026 | |||||
| BAF | TY stem/SE | - | 0.004 | - | 0.013 | |||||
| TY leaf/SE | - | 0.007 | 0.007 | 0.002 | ||||||
| TY/PW | TY root/PW | L kg−1 | 37 | - | 169 | |||||
| BCF | TY stem/PW | L kg−1 | 8 | - | 86 | |||||
| TY leaf/PW | L kg−1 | 13 | 28 | 14 | ||||||
| TY/RW | TY root/RW | L kg−1 | 503 | - | 772 | |||||
| BCF | TY stem/RW | L kg−1 | 106 | - | 392 | |||||
| TY leaf/RW | L kg−1 | 181 | 173 | 66 | ||||||
| TY | TF1 | TY leaf/stem | 1.70 | - | 0.17 | |||||
| TF2 | TY stem/root | 0.21 | - | 0.51 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Freire, A.; Turlin, F.; André-Mayer, A.-S.; Pelletier, M.; Cayer, A.; Giamberini, L. Biogeochemical Cycle of Lanthanides in a Light Rare Earth Element-Enriched Geological Area (Quebec, Canada). Minerals 2019, 9, 573. https://doi.org/10.3390/min9100573
Romero-Freire A, Turlin F, André-Mayer A-S, Pelletier M, Cayer A, Giamberini L. Biogeochemical Cycle of Lanthanides in a Light Rare Earth Element-Enriched Geological Area (Quebec, Canada). Minerals. 2019; 9(10):573. https://doi.org/10.3390/min9100573
Chicago/Turabian StyleRomero-Freire, Ana, François Turlin, Anne-Sylvie André-Mayer, Mia Pelletier, Alain Cayer, and Laure Giamberini. 2019. "Biogeochemical Cycle of Lanthanides in a Light Rare Earth Element-Enriched Geological Area (Quebec, Canada)" Minerals 9, no. 10: 573. https://doi.org/10.3390/min9100573