Lethal and Sub-Lethal Effects of Organic-Production-Approved Insecticides and Fungicides on the Predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae)

Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Experimental Setup
2.3. Statistical Analyses
3. Results
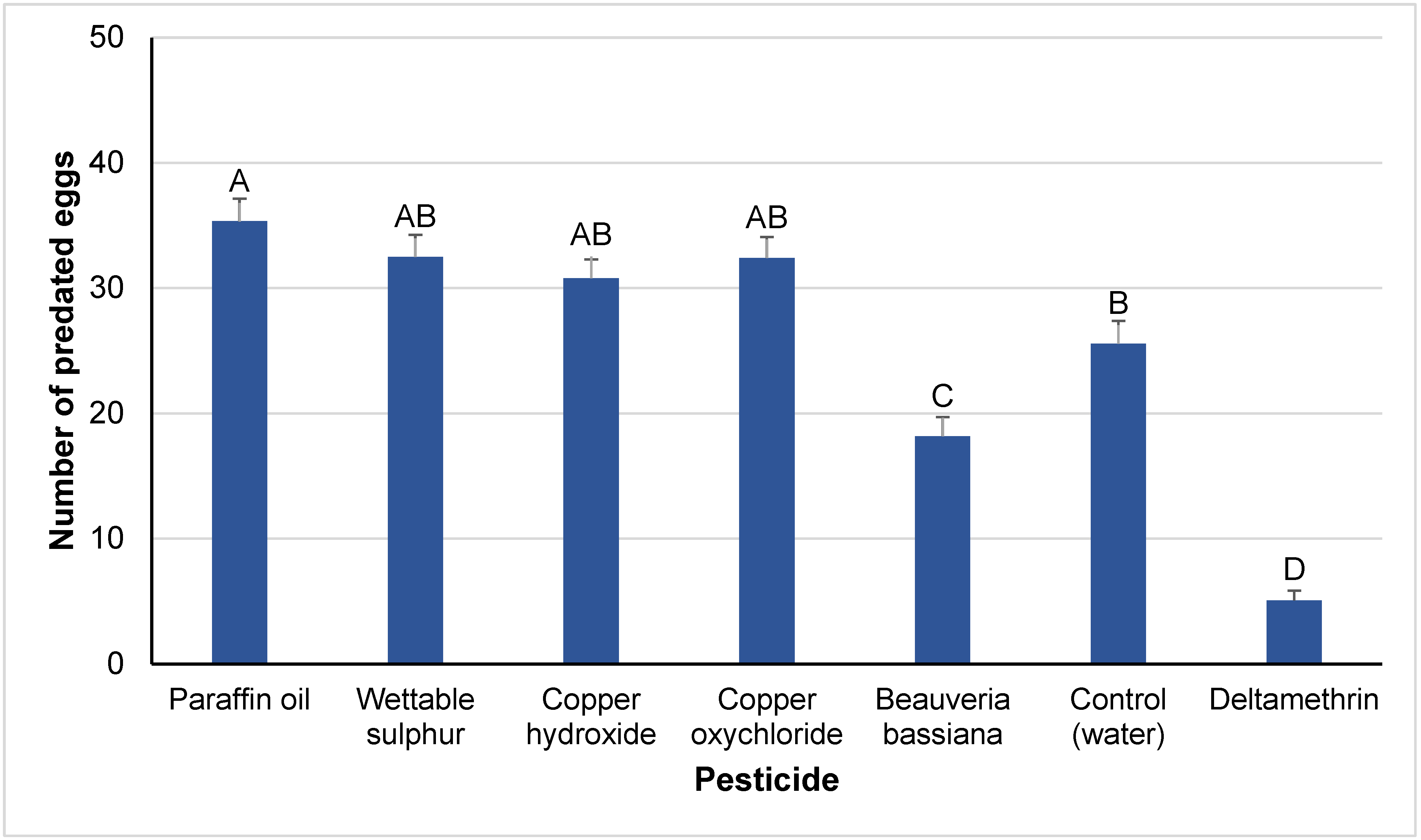
3.1. Lethal Effects
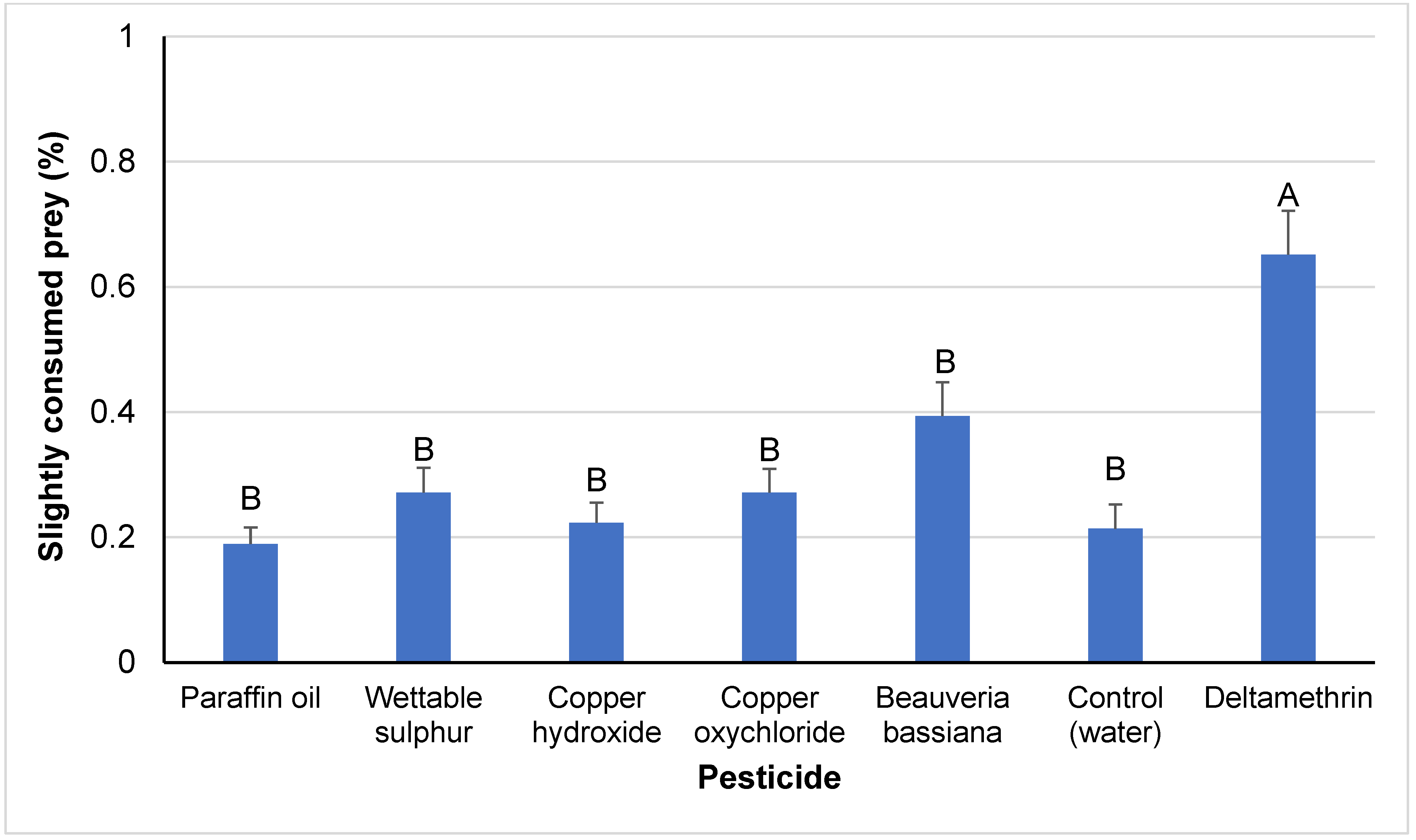
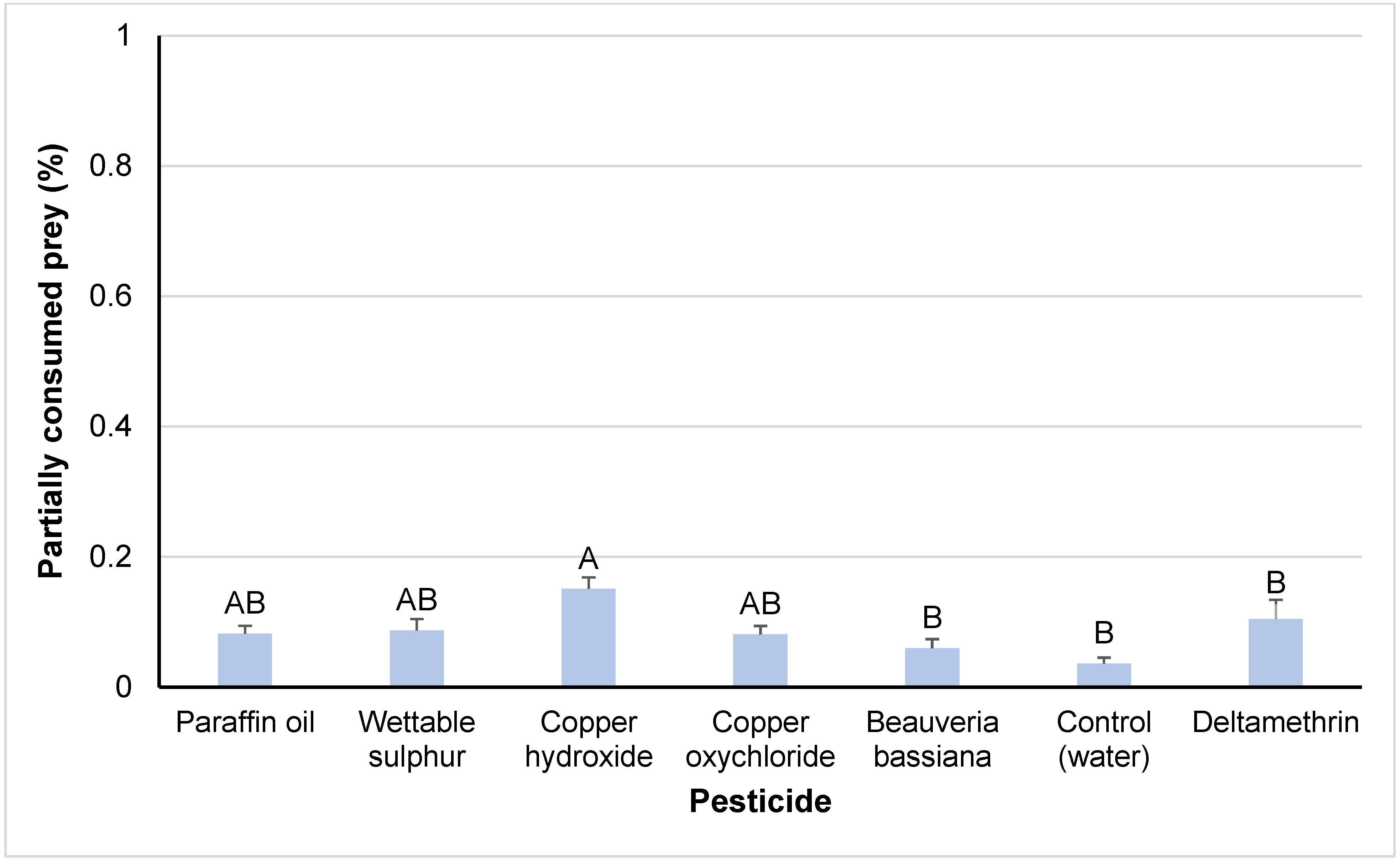
3.2. Sub-Lethal Effects
4. Discussion
4.1. Lethal Effects
4.2. Sub-Lethal Effects
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jacobson, R.J. IPM program for tomato. In Biocontrol in Protected Culture; Heinz, K.M., van Driesche, R.G., Parrella, M.P., Eds.; Ball Publishing: Batavia, IL, USA, 2004; pp. 457–471. [Google Scholar]
- Perdikis, D.; Kapaxidi, E.; Papadoulis, G. Biological control of insect and mite pests in greenhouse solanaceous crops. Eur. J. Plant Sci. Biotechnol. 2008, 2, 125–144. [Google Scholar]
- Richardson, E.B.; Troczka, B.J.; Gutbrod, O.; Davies, E.T.G.; Nauen, R. Diamide resistance: 10 years of lessons from lepidopteran pests. J. Pest Sci. 2020, 93, 911–928. [Google Scholar] [CrossRef]
- Biondi, A.; Guedes, R.N.C.; Wan, F.H.; Desneux, N. Ecology, worldwide spread and management of the invasive South American tomato pinworm, Tuta absoluta: Past, present and future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; Tsagarakou, A.; Vontas, J.; Nauen, R. Insecticide resistance in the tomato pinworm Tuta absoluta: Patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Buragohain, P.; Saikia, D.K.; Sotelo-Cardona, P.; Srinivasan, R. Development and validation of an integrated pest management strategy against the invasive South American tomato leaf miner, Tuta absoluta in South India. Crop Prot. 2021, 139, 105348. [Google Scholar] [CrossRef]
- Desneux, N.; Han, P.; Mansour, R.; Arno, J.; Brévault, T.; Campos, M.R.; Chailleux, A.; Guendes, R.; Karimi, J.; Konan, K.; et al. Integrated pest management of Tuta absoluta: Practical implementations across different world regions. J. Pest Sci. 2022, 95, 17–39. [Google Scholar] [CrossRef]
- Bahlai, C.A.; Xue, Y.; McCreary, C.M.; Schaafsma, A.W.; Hallett, R.H. Choosing organic pesticides over synthetic pesticides may not effectively mitigate environmental risk in soybeans. PLoS ONE 2010, 5, e11250. [Google Scholar] [CrossRef] [PubMed]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere 2012, 87, 803–812. [Google Scholar] [CrossRef]
- Niedobová, J.; Skalský, M.; Fric, Z.F.; Hula, V.; Brtnický, M. Effects of so-called “environmentally friendly” agrochemicals on the harlequin ladybird Harmonia axyridis (Coleoptera: Coccinelidae). Eur. J. Entomol. 2019, 116, 173–177. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Martinou, A.; Seraphides, N.; Stavrinides, M. Lethal and behavioral effects of pesticides on the insect predator Macrolophus pygmaeus. Chemosphere 2014, 96, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.E.S.; Alves, F.M.; Pereira, R.C.; Aquino, L.A.; Fernandes, F.L.; Zanuncio, J.C. Lethal and sublethal effects of seven insecticides on three beneficial insects in laboratory assays and field trials. Chemosphere 2016, 156, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Depalo, L.; Lanzoni, A.; Masetti, A.; Pasqualini, E.; Burgio, G. Lethal and sub-lethal effects of four insecticides on the aphidophagous coccinellid Adalia bipunctata (Coleoptera: Coccinellidae). J. Econ. Entomol. 2017, 110, 2662–2671. [Google Scholar] [CrossRef] [PubMed]
- Perdikis, D.; Psaroudaki, S.; Papadoulis, G. Compatibility of Nesidiocoris tenuis and Iphiseius degenerans with insecticides, miticides and fungicides used in tomato crops. Bull. Insectol. 2020, 73, 181–192. [Google Scholar]
- Desneux, N.; Pham-Delègue, M.H.; Kaiser, L. Effects of sub-lethal and lethal doses of lambda-cyhalothrin on oviposition experience and host-searching behaviour of a parasitic wasp, Aphidius ervi. Pest Manag. Sci. 2004, 60, 381–389. [Google Scholar] [CrossRef]
- Perdikis, D.; Fantinou, A.; Lykouressis, D. Enhancing pest control in annual crops by conservation of predatory Heteroptera. Biol. Control 2011, 59, 13–21. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Riahi, C.; Urbaneja, A. Use of zoophytophagous mirid bugs in horticultural crops: Current challenges and future perspectives. Pest Manag. Sci. 2021, 77, 33–42. [Google Scholar] [CrossRef]
- Martinou, A.F.; Stavrinides, M.C. Effects of sublethal concentrations of insecticides on the functional response of two mirid generalist predators. PLoS ONE 2015, 10, e0144413. [Google Scholar] [CrossRef]
- Perdikis, D.C.; Arvaniti, K.; Paraskevopoulos, A.; Grigoriou, A. Pre-plant release enhanced the earlier establishment of Nesidiocoris tenuis in open field tomato. Entomol. Hellen. 2015, 24, 11–21. [Google Scholar] [CrossRef]
- Dáder, B.; Colomer, I.; Adán, Á.; Medina, P.; Viñuela, E. Compatibility of early natural enemy introductions in commercial pepper and tomato greenhouses with repeated pesticide applications. Insect Sci. 2020, 27, 1111–1124. [Google Scholar] [CrossRef]
- Ziaei Madbouni, M.A.; Samih, M.A.; Qureshi, J.A.; Biondi, A.; Namvar, P. Compatibility of insecticides and fungicides with the zoophytophagous mirid predator Nesidiocoris tenuis. PLoS ONE 2017, 12, e0187439. [Google Scholar] [CrossRef] [PubMed]
- Fantinou, A.A.; Perdikis, D.; Maselou, D.; Labropoulos, P. Prey killing without consumption: Does Macrolophus pygmaeus show adaptive foraging behaviour? Biol. Control 2008, 47, 187–193. [Google Scholar] [CrossRef]
- Lykouressis, D.; Perdikis, D.; Mandarakas, I. Partial consumption of different species of aphid prey by the predator Macrolophus pygmaeus (Hemiptera: Miridae). Eur. J. Entomol. 2016, 113, 345–351. [Google Scholar] [CrossRef]
- Prieto, J.D.; Trotta, V.; Fanti, P.; Castañé, C.; Battaglia, D. Predation by Macrolophus pygmaeus (Hemiptera: Miridae) on Acyrthosiphon pisum (Hemiptera: Aphididae): Influence of prey age/size and predator’s intraspecific interactions. Eur. J. Entomol. 2016, 113, 37–43. [Google Scholar] [CrossRef]
- Mahdian, K.; Van Leeuwen, T.; Tirry, L.; De Clercq, P. Susceptibility of the predatory stinkbug Picromerus bidens to selected insecticides. BioControl 2007, 52, 765–774. [Google Scholar] [CrossRef]
- FRAC Code List 2018: Fungicides Sorted by Mode of Action. Available online: http://www.phi-base.org/images/fracCodeList.pdf (accessed on 10 September 2022).
- IRAC Mode of Action Classification Scheme (Version 10.1); IRAC International MoA Working Group: 2021. Available online: https://www.frac.info/knowledge-database/downloads (accessed on 10 September 2022).
- Amarasekare, K.G.; Shearer, P.W. Laboratory bioassays to estimate the lethal and sublethal effects of various insecticides and fungicides on Deraeocoris brevis (Hemiptera: Miridae). J. Econ. Entomol. 2013, 106, 776–785. [Google Scholar] [CrossRef]
- SAS Institute. JMP, Version 14.0; SAS Institute Inc.: Cary, NC, USA, 2016.
- Sterk, G.; Hassan, S.A.; Baillod, M.; Bakker, F.; Bigler, F.; Blumel, S.; Bogenschutz, H.; Boller, E.; Bromand, B.; Brun, J.; et al. Results of the seventh joint pesticide testing programme. IOBC/WPRS working group ‘‘pesticides and beneficial organisms’’. BioControl 1999, 44, 99–117. [Google Scholar] [CrossRef]
- Hamdi, F.; Fargues, J.; Ridray, G.; Jeannequin, B.; Bonato, O. Compatibility among entomopathogenic hyphocreales and two beneficial insects used to control Trialeurodes vaporariorum (Hemiptera: Aleurodidae) in Mediterranean greenhouses. J. Invertebr. Pathol. 2011, 108, 22–29. [Google Scholar] [CrossRef]
- Malekan, N.; Hatami, B.; Akhavan, A.; Ebadi, R.; Radjabi, R. Evaluation of entomopathogenic fungi Beauveria bassiana and Lecanicillium muscarium on different nymphal stages of greenhouse whitefly Trialeurodes vaporariorum in greenhouse conditions. Biharean Biologist 2015, 9, 108–112. [Google Scholar]
- Angeli, G.; Baldessari, M.; Maines, R.; Duso, C. Side-effects of pesticides on the predatory bug Orius laevigatus (Heteroptera: Anthocoridae) in the laboratory. Biocontrol Sci. Technol. 2005, 15, 745–754. [Google Scholar] [CrossRef]
- DeLong, D.M. Studies of Methods and Materials for the Control of the Leafhopper Empoasca fabae as a Bean Pest. Techn. Bull. 1940, 740. Available online: https://handle.nal.usda.gov/10113/CAT86200734 (accessed on 10 September 2022).
- Zappalá, L.; Siscaro, G.; Biondi, A.; Mollá, O.; Gonzalez-Cabrera, J.; Urbaneja, A. Efficacy of sulphur on Tuta absoluta and its side effects on the predator Nesidiocoris tenuis. J. Appl. Entomol. 2012, 136, 401–409. [Google Scholar] [CrossRef]
- van de Veire, M.; Sterk, G.; Van der Staaij, M.; Ramakers, P.M.J.; Tirry, L. Sequential testing scheme for the assessment of the side-effects of plant protection products on the predatory bug Orius laevigatus. BioControl 2002, 47, 101–113. [Google Scholar] [CrossRef]
- Wanumen, A.C.; Sanchez-Ramos, I.; Vinuela, E.; Medina, P.; Andán, A. Impact of feeding on contaminated prey on the life parameters of Nesidiocoris tenuis (Hemiptera: Miridae) adults. J. Insect Sci. 2016, 16, 103. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Norum, U.; Friberg, N.; Jensen, M.R.; Pedersen, J.M.; Bjerregaard, P. Behavioural changes in three species of freshwater macroinvertebrates exposed to the pyrethroid lambda-cyhalothrin: Laboratory and stream microcosm studies. Aquat. Toxicol. 2010, 98, 328–335. [Google Scholar] [CrossRef]
- Labbè, R.M. Intraguild Interactions of the Greenhouse Whitefly Natural Enemies, Predator Dicyphus hesperus, Pathogen Beauveria bassiana and Parasitoid Encarsia formosa. Master’s Thesis, Faculté des Sciences de l’Agriculture et de l’Alimentation Université, Laval Québec, Québec, QC, Canada, 2005. Available online: http://hdl.handle.net/20.500.11794/18015 (accessed on 3 September 2022).
- Wu, S.; Xing, Z.; Sun, W.; Xu, X.; Meng, R.; Lei, Z. Effects of Beauveria bassiana on predation and behavior of the predatory mite Phytoseiulus persimilis. J. Invertebr. Pathol. 2018, 153, 51–56. [Google Scholar] [CrossRef]
- Alma, C.R.; Gillespie, D.R.; Roitberg, B.D.; Goettel, M.S. Threat of infection and threat-avoidance behavior in the predator Dicyphus hesperus feeding on whitefly nymphs infected with an entomopathogen. J. Insect Behav. 2010, 23, 90–99. [Google Scholar] [CrossRef]
- Meyling, N.V.; Pell, J.K. Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol. Entomol. 2006, 31, 162–171. [Google Scholar] [CrossRef]
- Herrick, N.J.; Cloyd, R.A. Direct and indirect effects of pesticides on the insidious flower bug (Hemiptera: Anthocoridae) under laboratory conditions. J. Econ. Entomol. 2017, 110, 931–940. [Google Scholar] [CrossRef]
- Sahid, A.; Natawigena, W.D.; Hersanti; Sudarjat. Laboratory rearing of Sycanus annulicornis (Hemiptera: Reduviidae) on two species of prey: Differences in its biology and efficiency as a predator of the nettle caterpillar pest Setothosea asigna (Lepidoptera: Limacodidae). Eur. J. Entomol. 2018, 115, 208–216. [Google Scholar] [CrossRef]
- Matioli, T.F.; da Silva, M.R.; de Bastos Pazini, J.; Barroso, G.; Vieira, J.G.A.; Yamamoto, P.T. Risk assessment of insecticides used in tomato to control whitefly on the predator Macrolophus basicornis (Hemiptera: Miridae). Insects 2021, 12, 1092. [Google Scholar] [CrossRef] [PubMed]
- Roda, A.; Castillo, J.; Allen, C.; Urbaneja, A.; Pérez-Hedo, M.; Weihman, S.; Stansly, P.A. Biological control potential and drawbacks of three zoophytophagous mirid predators against Bemisia tabaci in the United States. Insects 2020, 11, 670. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name/a.i. | Mode of Action | Chemical Class | Brand Name/Formulation | Manufacturer | Highest Label Rate g a.i./L |
|---|---|---|---|---|---|
| Paraffin oil-Mineral oil | NC | Diverse | Nitropol O EW | W. Neudorff, Athens, Emmerthal, Germany | 5.46 |
| Sulfur | M2 | Inorganic | Frame 80 WG | Agrostulln, Stulln, Germany | 2.4 |
| Copper hydroxide | M1 | Inorganic | Champ 36 SC | Nufarm, Melbourne, Austria | 1.27 |
| Copper oxychloride | M1 | Inorganic | Cuprachlor 50 WP | Industrias Quimicas del Vallès, S.A, Barcelona, Spain | 1.25 |
| Beauveria bassiana | UNF | Entomopathogen | Metab SL | Microspore-Sacom, Greece | 11.5 × 107 conidia |
| Deltamethrin | 3A | Pyrethroid | Decis 2.5 EC | Bayer S.A.S, Lyon, France | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betsi, P.C.; Perdikis, D.C. Lethal and Sub-Lethal Effects of Organic-Production-Approved Insecticides and Fungicides on the Predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae). Insects 2023, 14, 866. https://doi.org/10.3390/insects14110866
Betsi PC, Perdikis DC. Lethal and Sub-Lethal Effects of Organic-Production-Approved Insecticides and Fungicides on the Predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae). Insects. 2023; 14(11):866. https://doi.org/10.3390/insects14110866
Chicago/Turabian StyleBetsi, Petri C., and Dionysios Ch Perdikis. 2023. "Lethal and Sub-Lethal Effects of Organic-Production-Approved Insecticides and Fungicides on the Predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae)" Insects 14, no. 11: 866. https://doi.org/10.3390/insects14110866