

Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone

1
Guangdong Province Key Laboratory of Biotechnology Drug Candidates, School of Life Sciences and Biopharmaceuticals, Guangdong Pharmaceutical University, Guangzhou 510006, China
2
School of Health Sciences, Guangzhou Xinhua University, 19 Huamei Road, Tianhe District, Guangzhou 510520, China
3
State Key Laboratory of Biocontrol, Guangdong Key Laboratory of Pharmaceutical Functional Genes, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Insects 2023, 14(11), 871; https://doi.org/10.3390/insects14110871
Submission received: 15 October 2023
/
Revised: 6 November 2023
/
Accepted: 10 November 2023
/
Published: 12 November 2023
(This article belongs to the Special Issue Insect Hormones, Metabolism, Immunity and Development)
Abstract
:Simple Summary
Insects rely on a sterol hormone known as 20-Hydroxyecdysone to regulate vital cellular activities. This study delves deep into the biosynthesis of this hormone, its signal transduction, and the mechanisms by which it regulates autophagy (cellular self-cleaning) and apoptosis (programmed cell death). Our primary goal was to unravel the intricate interplay between 20-Hydroxyecdysone and these cellular processes. Through our research, we uncovered the intricate pathways and mechanisms that this hormone influences in cell renewal and controlled cell death. These insights not only hold significance for comprehending insect development and survival but also have broader implications in fields like agriculture and pest control. By shedding light on these processes, we aim to provide valuable knowledge that can be leveraged for the betterment of our environment and society.
Abstract
20E (20-Hydroxyecdysone) is a central steroid hormone that orchestrates developmental changes and metamorphosis in arthropods. While its molecular mechanisms have been recognized for some time, detailed elucidation has primarily emerged in the past decade. PCD (Programmed cell death), including apoptosis, necrosis, efferocytosis, pyroptosis, ferroptosis, and autophagy, plays a crucial role in regulated cell elimination, which is vital for cells’ development and tissue homeostasis. This review summarizes recent findings on 20E signaling regulated autophagy and apoptosis in insects, including Drosophila melanogaster, Bombyx mori, Helicoverpa armigera, and other species. Firstly, we comprehensively explore the biosynthesis of the sterol hormone 20E and its subsequent signal transduction in various species. Then, we focus on the involvement of 20E in regulating autophagy and apoptosis, elucidating its roles in both developmental contexts and bacterial infection scenarios. Furthermore, our discussion unfolds as a panoramic exposition, where we delve into the fundamental questions with our findings, anchoring them within the grander scheme of our study in insects. Deepening the understanding of 20E-autophagy/apoptosis axis not only underscores the intricate tapestry of endocrine networks, but also offers fresh perspectives on the adaptive mechanisms that have evolved in the face of environmental challenges.
1. Introduction
Ecdysteroids, acknowledged as the principal steroid hormones in arthropods, are commonly known as “molting hormones” in insect physiology [1]. Ecdysone serves as a prohormone crucial for synthesizing the primary insect molting hormone, 20E. Once ecdysone is released, it undergoes conversion into the more active form, 20E. This hormone is renowned for its central role in regulating various life stages in insects, facilitating the smooth transition from larval stages to pupation and eventually to maturity. It exerts a significant influence on cellular processes such as autophagy and apoptosis, which are essential for the developmental changes that occur during molting [2]. These cellular events are integral to insect development, growth, and survival, especially in species like D. melanogaster (Drosophila melanogaster), B. mori (Bombyx mori), H. armigera (Helicoverpa armigera), and others [3,4,5,6].
PCD is a vital cellular mechanism in insects, ensuring the proper development, maintenance, and survival [7,8]. Processes such as apoptosis, necrosis, and autophagy play pivotal roles in tissue remodeling and the removal of damaged cells [9]. In advanced eukaryotes, three main autophagy types have been identified: macroautophagy, microautophagy, and chaperone-mediated autophagy. Both microautophagy and chaperone-mediated autophagy direct cytosol segments and proteins to lysosomes, either autonomously or with assistance by chaperones [10]. Macroautophagy, often referred to as autophagy, is a recycling mechanism crucial for cellular homeostasis, guiding developmental shifts and addressing environmental stresses, especially in insects [11]. Apoptosis emerges as a pivotal process-shaping development, homeostasis, and defense [12]. Autophagy and apoptosis in insects have been extensively studied, not only in the midgut epithelium of Acheta domesticus (Insecta, Orthoptera, and Gryllidae), but also in the context of insect infections [13,14]. This indicates that both autophagy and apoptosis play indispensable roles in cellular development. Furthermore, 20E emerges as a key regulator of autophagy and apoptosis. In the silk-producing B. mori, a lepidopteran species, 20E’s regulation of autophagy and apoptosis are vital for its unique life cycle stages [15]. Similarly, the malaria vector, Anopheles gambiae, as an Diptera species depends on the processes regulated by 20E for its rapid larval development [16]. On the other hand, in the cotton bollworm, H. armigera, the interplay between 20E and other signaling pathways modulates PCD, which is crucial for its adaptability to diverse environments [6]. Collectively, 20E regulates autophagy, apoptosis, immune function, and the central nervous system, indicating high antioxidant activities and other functions [17]. This evolutionary pattern highlights the unique biological responses across different life phases. Therefore, it is crucial to assess regulatory mechanisms through the lens of age and developmental transitions.
The understanding between 20E, autophagy, and apoptosis holds profound implications beyond basic biology. In agriculture, a comprehensive understanding of this triad could pave the way for innovative pest control methods, reducing the reliance on environmentally harmful pesticides. This synergistic connection between 20E and autophagy/apoptosis in insects not only underscores the complexity of nature but also presents a fertile ground for potential advancements in both sustainable agriculture and medicine.
2. Sterol Hormone 20E Biosynthesis
From a biochemical perspective, the sterol hormone 20E, which belongs to the ecdysteroid family, plays a pivotal role in orchestrating insect growth, development, and reproduction [18,19]. Its biosynthesis begins with the assimilation of dietary cholesterol, serving as the foundational precursor. The process of 20E biosynthesis in insects holds a central process governing the development and metamorphosis [20]. Biosynthesis of 20E involves a series of enzymatic reactions that convert cholesterol into the active hormone. Initially, cholesterol undergoes a hydroxylation process, which is catalyzed by several cytochrome P450 enzymes, leading to the production of precursors such as ecdysone. Subsequent enzymatic steps, including further hydroxylation and oxidation, result in the formation of 20E. Upon synthesis, 20E targets specific nuclear receptors, mainly the EcR (ecdysone receptor), to regulate gene expression essential for developmental processes [21]. This bioactive form of ecdysone then participates in the regulation of gene expression by forming a heterodimer with USP (ultraspiracle protein), which is essential for initiating the developmental processes [22]. The genes EcR-B1 (ecdysone receptor B1 isoform) and USP1 (ubiquitin specific peptidase 1) have been predominantly associated with 20E regulation [23,24]. Intriguingly, other pivotal factors such as E75B and Ha-eIF5c have also been identified to influence 20E regulation. It was elucidated that the genes modulated by 20E, namely EcR-B1, USP1, E75B, BR-CZ2, HHR3, and Ha-eIF5c, are influenced by the suppressive actions of both Ha-Ntf2 and Ha-Ran [25,26,27]. Taken together, these evidences suggest that the regulation of 20E exhibits conserved mechanisms across diverse species. These information provide a coherent framework, tracing from its formation, through its process, and to its ultimate effects. Therefore, a comprehensive understanding of 20E biosynthesis yields profound insights into its subsequent roles in biological metabolism.
E (Ecdysone), which is the immediate precursor of 20E, is synthesized and released via specialized structures in insects, such as the prothoracic glands or ovaries. After secretion, it circulates in the hemolymph and is transformed into the biologically active 20E in peripheral tissues. This conversion primarily occurs in the fat body, midgut, and malpighian tubules, particularly during the larval stage [28]. Illustratively, in Drosophila and other members of the Diptera order, ecdysone is synthesized and secreted via the prothoracic glands, acting as the seminal trigger for the initiation of the molting cascade [29]. Subsequently, E undergoes hydroxylated at the 20th position by the enzyme, shade, leading to the formation of the biologically activated 20E being released from the fat body and transported to different cells and tissues as needed. In the adults of some insects, the presence of ecdysteroids in the ovary was discovered 40 years ago, and now it is well established that ovarian follicular cells synthesize ecdysone de novo [30,31]. Intriguingly, in certain lepidopteran species, it has been documented that male gonads release significant quantities of ecdysone in vitro [32].
The biosynthetic cascade is triggered when the PTTH (neuropeptide Prothoracicotropic Hormone) binds to its receptor, Torso, on prothoracic gland cells [33]. This interaction initiates the Ras/Raf/ERK signaling pathway, activating genes crucial for ecdysone biosynthesis [34]. Simultaneously, the Insulin/IIS (IGF Signaling) pathway, responsive to nutritional cues, enhances ecdysone synthesis by increasing the expression of biosynthetic enzymes and by augmenting the prothoracic glands’ sensitivity to PTTH [35]. The biosynthetic process is further modulated by biogenic amines, particularly octopamine, and neuropeptides such as B. mori, although their roles may vary across different species. Significantly, external factors such as photoperiodicity and ambient temperature subtly influence this biosynthetic pathway [36,37,38,39]. This biosynthesis involves cytochrome P450 enzymes, with six identified as playing roles in the 20E pathway [40]. While the cytochrome P450 Cyp6t3 has been shown not to be essential for ecdysone biosynthesis in D. melanogaster, CYP18A1 is still crucial for its metamorphosis [41,42]. The biosynthesis of 20E involves multiple cytochrome P450 enzymes, including CYP306A1, which facilitates the production of the arthropod molting hormone [43]. These findings provide a new perspective, suggesting that P450 may have a more pronounced role in regulating the sterol hormone related to cellular processes.
Moreover, JH (Juvenile Hormone) adds an additional layer of regulation, often acting antagonistically to 20E. Elevated JH concentrations typically suppress 20E synthesis [44]. In the captivating world of beetles, as part of the Coleoptera order, the JH takes center stage, driving vitellogenesis within the fat body [45,46]. Vitellogenesis involves the synthesis and accumulation of yolk proteins in maturing oocytes, which are crucial for nourishing the embryo post-fertilization [47]. This crucial process primarily occurs in the fat body, an organ comparable to the mammalian liver. Interestingly, in the adult females of Hemimetabola (Dictyoptera to Hemiptera) and Coleoptera, JH serves as the main regulator, exerting pleiotropic controls over various aspects of female reproduction [48,49]. Given the diversity of Coleoptera, there is variability in how different beetle species respond to JH, reflecting their specific reproductive strategies and ecological niches. Furthermore, the coordinated interplay of these PCD forms, observed in the ovarian tropharia of certain beetles, underscores the delicate equilibrium insects establish to ensure their survival and adaptability amidst constantly evolving environments [50]. While ecdysone, 20E, and JH each govern insect growth and reproduction, an intriguing question arises: Could 20E also have a pivotal role in the insect reproductive processes? If so, what are the underlying mechanisms through which these hormones operate? Unraveling these questions could provide fresh insights into the multifaceted roles of steroid hormones in insect biology.
In the silkworm B. mori, the enzymatic activity of ecdysteroid-phosphate phosphatase plays a crucial role in the 20E biosynthetic pathway within eggs, catalyzing the conversion of maternal-conjugated ecdysteroids [5]. This process is paralleled in the species Drosophila, belonging to Diptera, where studies have uncovered a reciprocal regulatory relationship between 20E and JH in larvae [51]. Each hormone intricately influences the synthesis of the other, showcasing the complexity of hormonal interactions during development. Recent studies have revealed that 20E, which is specifically derived from cholesterol via the catalytic activity of a series of cytochrome P450 enzymes that hydroxylate E at carbon 20, particularly CYP314a1, is essential for insect molting and metamorphosis [52]. This underscores the significance of Drosophila E-20-monooxygenase, which is identified as the product of the shade locus. In a recent investigation focused on H. armigera, a representative of the Lepidoptera order, four cytochrome P450 homologs-HarmCYP302A1, HarmCYP306A1, HarmCYP314A1, and HarmCYP315A1, were identified. These homologs demonstrated evolutionary conservation across lepidopterans. Intriguingly, while HarmCYP302A1 and HarmCYP315A1 were predominantly expressed in the larval prothoracic glands, such consistent expression patterns were absent for HarmCYP306A1 and CYP314A1 [53]. It is evident that, at different times, the biological activity and expression of different enzymes can serve as indicators of biological activity to some extent. However, the question remains: Is the emergence of biological activity necessarily related to these enzymes?
In conclusion, the intricate process of 20E biosynthesis serves as a cornerstone in the field of insect physiology and development. A comprehensive grasp of this pathway not only sheds light into the evolutionary intricacies of insect life, but also offers promising opportunities for transformative applications in pest management, medicine, and biotechnology. As we unravel the steps and intricacies of 20E biosynthesis, we gain a deeper comprehension of insect biology and open avenues for sustainable and innovative solutions to challenges in agriculture and health.
3. 20E Signal Transduction
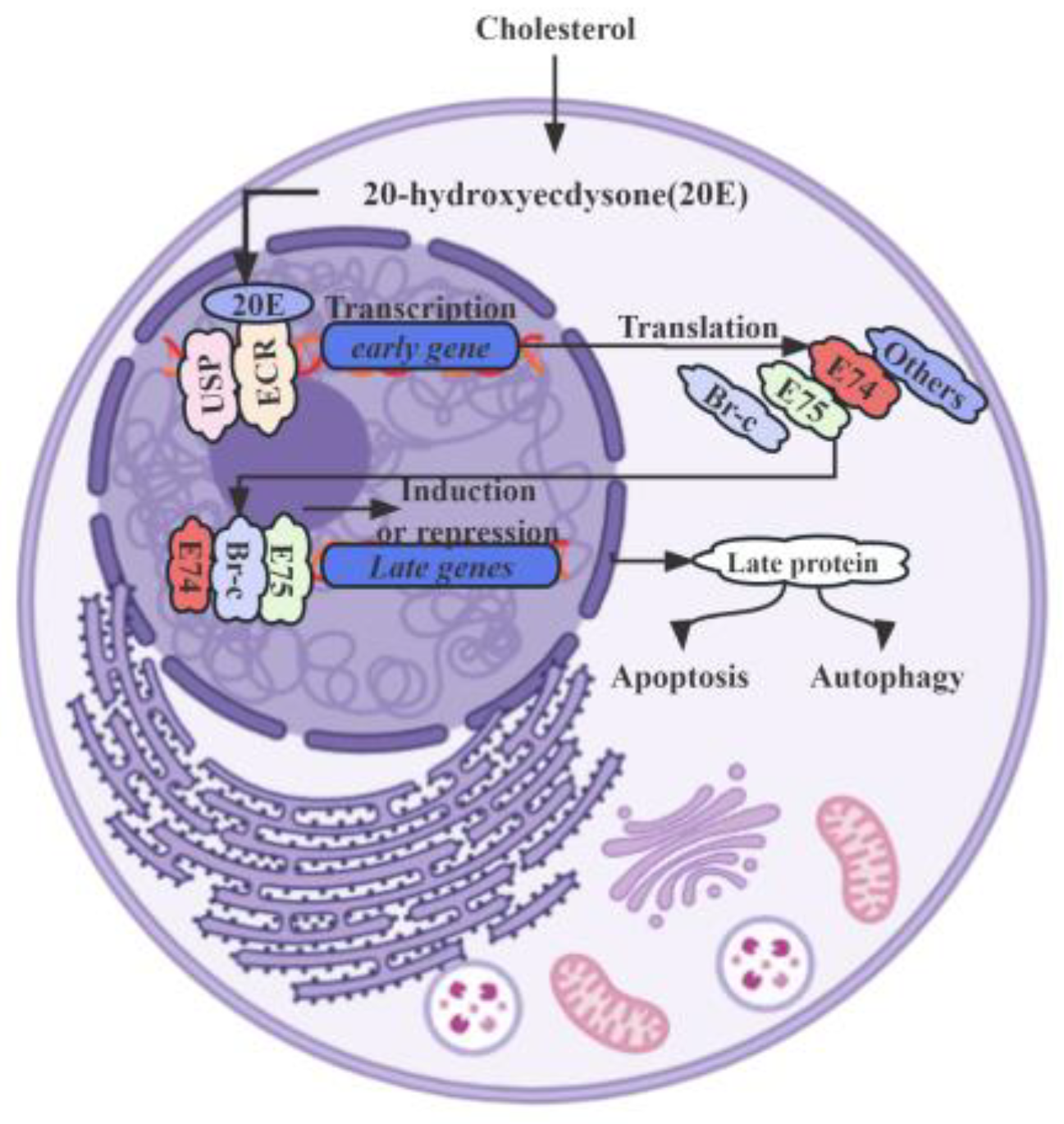
Specifically, 20E counteracts IIS through the AMPK (AMP-activated protein kinase) and PP2A (protein phosphatase 2A) axis within the insect fat body, leading to a suppression of growth rates. Increased levels of 20E activate AMPK, a pivotal molecular sensor responsible for maintaining energy equilibrium in the insect fat body, of which the subsequently activated AMPK stimulates PP2A, which in turn dephosphorylates the insulin receptor and AKT (protein kinase B), culminating in the inhibition of the IIS signaling pathway [15]. Moreover, E93 (Ecdysone-induced protein 93), encoded by a member of the helix-turn-helix transcription factor family, also plays a crucial role in the crosstalk of 20E signaling with JH signaling, mediated by the JH primary response gene Kr-h1 [54]. These transcription factor genes, described as “primary response genes”, are activated and transcribed within minutes after 20E stimulation, without requiring de novo protein synthesis. Upon binding to its receptor complex, EcR-USP, 20E rapidly and strongly induces the expression of primary-response genes. These genes, including nuclear receptors and transcription factors like E75 (Ecdysone-induced protein 75), E93, Br-C (Broad-Complex), and E74 (Ecdysone-induced protein 74), play vital roles in the larval–pupal metamorphosis of insects (Figure 1) [54,55,56].
The genes responsible for E biosynthesis are essential for producing the precursor hormone 20E, exhibiting distinct expression patterns in the tubular accessory glands of adult male insects [57]. In the remodeling fat body of Drosophila, which serves as an in vivo model, research has revealed a complex molecular interplay between autophagy and caspase activity. Despite initially seeming to be opposing processes, both autophagy and caspase activity are triggered via the common stimulus of 20E. Although they counterbalance each other, obstruction in one pathway amplifies the other. This dynamic interplay is further highlighted via the observed progressive augmentation of both autophagy and caspase activity in the remodeling fat body, triggered via a surge of the molting hormone, 20E, during the pivotal larval–prepupal transition [58]. Additionally, research has elucidated the antagonistic relationship between 20E and the insulin/IIS pathway.
In B. mori, a thorough analysis of the EcR-A and EcR-B1 expression during the larval–pupal transition has revealed their upregulation by 20E, primarily in the lateral neurosecretory cells of the larval brain, which are known as prothoracicotropic hormone-producing cells [23]. Significantly, 20E signaling predominantly mediates the occurrence of autophagy in larval tissues and organs during larval molting and larval–pupal metamorphosis [59]. The 20E-EcR-USP complex initially induces the expression of a small set of primary response genes, including transcription factors such as Br-C, E74, E75, and E93, which are responsible for the upregulation of a larger set of downstream secondary response genes and crucial for successful molting or metamorphosis (Table 1). In a distinct study, it was demonstrated that 20E plays a role in the elongation of genital disks. The precursor hormone 20E significantly induced the expression of three different BmE75 isofoms in Bombyx, which consequentially coordinated feedback to 20E biosynthesis [55]. Both BIGFLP (Bombyx imp-L2 gene-enhancer flippase protein) and 20E hormones were discovered to synergistically stimulate protein synthesis, where the modulatory effects were mediated via the IIS pathway and the mitogen-activated protein kinase pathway [60]. This coordination between BIGFLP and 20E underscores their joint regulation of genital disk growth and development. Notably, RNA interference have revealed that the downregulation of BmADK leads to the decreased expression of ATG-8, Caspase-9, Ec-R, E74A, and Br-C [61]. This suggests BmADK’s potential involvement in 20E signaling, apoptosis, and autophagy, all of which are essential for silk gland degeneration and silkworm metamorphosis. However, the precise role that 20E plays in these vital processes remains insufficiently elucidated.
Additionally, studies focusing on HaP60 (H. armigera P60) have revealed its phosphorylation under the regulation of insulin-like peptides during larval growth stages. In metamorphosis, the dephosphorylation of HaP60, coupled with its upregulation in response to 20E, orchestrates a series of protein phosphorylations, which leads to the cytosolic localization of forkhead box protein O subsequently promoting cellular proliferation, indicating a subtle connection that 20E has an impact on the additional regulation of autophagy and apoptosis [62]. Simultaneously, there was a significant downregulation of several 20E-responsive genes, such as LdEcR, LdUSP, LdHR3, and LdFTZ-F1. Intriguingly, the silencing of LdTorso also led to the upregulation of the JH biosynthesis gene larval diapause hormone acid methyltransferase, leading to elevated JH titers and the subsequent activation of the early-inducible JH gene, LdKr-h1 [63].
Collectively, the cytochrome P450 enzymes and primary-response genes play a crucial role in the biosynthesis of 20E across different species (Table 1). The precise modulation of gene expression and cellular processes by 20E signaling underscores its evolutionary significance and its pivotal role in insect biology. Delving into the complexities of 20E signal transduction not only sheds light on the molecular mechanisms that drive insect development and metamorphosis, but also reveals potential strategies for targeted pest management and other practical applications.

{kind=link}
{kind=link}
Table 1.
The cytochrome P450 enzymes and primary-response genes in 20E signal transduction.
| Species | Cytochrome P450 Enzymes | The Primary-Response Genes |
|---|---|---|
| D. melanogaster | DmCYP18a1 [41] DmCyp6a8/DmCyp6a2 [64] DmCyp6g1 [65] DmCYP302A1, DmCYP315A1 and DmCYP314A1 [66] | DmE75A [67] DmE75B, DmE75C and DmE75D [68] DmE93, DmE74, Br-C, and β-FTZ-F1 [69] |
| B. mori | BmCyp450, BmCYP314a1, BmCYP315a1, BmCYP302a1 and BmCYP306a1 [70] BmCYP4 and CYP6 [71] BmCYP18A1 [72] | BmE75A, BmE75B and BmE75C [73] BmE93 and BR-C [74] EcR-A and BHR3 [75] |
| H. armigera | HarmCYP302A1, HarmCYP306A1, HarmCYP314A1, and HarmCYP315A1 [53] HarmCYP6b2, HarmCYP18a1 and HarmCYP18b1 [76] | Br-C, E74, Ha-Ntf2, Ha-Ran, E75B, HHR3, and Ha-eIF5c [25] |
4. 20E Regulates Autophagy and Apoptosis in Insects
While the precise mechanisms underlying how 20E influences PCD are not yet fully elucidated, preliminary discoveries suggest that the potential involvement of the enzymatic machinery are responsible for 20E in the transformation of dietary sterols into ecdysteroids [4]. The steroid hormone 20E, with its diverse functions in regulating autophagy, apoptosis, and various cellular processes, is gradually emerging as a key player in the orchestration of insect growth and metamorphic transitions.
The study suggested that specific neurons, particularly RP2s, provide valuable insights into the molecular dynamics of neuronal demise during insect metamorphosis [77]. This process is likely regulated by steroid hormones, with 20E assuming a crucial role in coordinating both autophagy and apoptosis in these organisms. Moreover, it has been observed that 20E can influence intracellular calcium levels, consequently shifting from autophagic cell survival to apoptotic cell death [17,78]. Another research has illuminated the role of 20E in upregulating the transcription factor Klf15 (Krüppel-like factor) by enhancing Klf15 transcription in the metamorphic fat body through its nuclear receptor EcR, thereby promoting autophagy, apoptosis, and gluconeogenesis during metamorphosis [79]. Furthermore, studies have emphasized the reduction in glycolysis and PGK1 expression levels during metamorphosis under the regulation of 20E. While insulin promotes glycolysis and cell proliferation via PGK1 phosphorylation, 20E counteracted this by dephosphorylating PGK1 via PTEN (phosphatase and tensin homolog), thereby inhibiting glycolysis [80]. Moreover, a separate study highlighted that even in the absence of E or 20E synthesis, specific mutants still exhibit transcriptional enhancement of glycolytic genes during embryogenesis [81]. This suggests that these hormones may not be crucial for this particular metabolic transition, further emphasizing the intricate role 20E plays in regulating PCD. During insect metamorphosis, cell death artfully transforms larval forms, paving the way for adulthood. Modern research into Lepidoptera and Diptera metamorphosis unveils the intricate ballet of tissue change and its governing regulations [82]. In the midgut, apoptotic cells are cast into the lumen, initially dispersed but eventually forming a cohesive layer beneath the peritrophic membrane. Intriguingly, only the youthful midgut epithelium shows apoptosis. As it matures, necrosis emerges alongside apoptosis, eventually taking precedence and ultimately overshadowing apoptosis completely [83]. Within the intricate cellular machinery of mammals, apoptosis is marked by the coordinated movement of Cyt-c (cytochrome C) from its protected location within the mitochondria to the expansive environment of the cytoplasm [84]. Understanding the roles of 20E can potentially inform pest control strategies, given the hormone’s influence on insect development and survival.
4.1. Regulation in D. melanogaster
In the developmental stages of Drosophila, the steroid hormone E plays a crucial role in guiding the transition from the embryo to the larva and subsequently to the pupa. It is notably during the larval–pupal metamorphosis that a significant wave of PCD occurs, efficiently eliminating redundant larval tissues. E’s periodic surges regulate the activation of distinct transcription factors, which, in turn, promote the expression of crucial genes associated with cell death [85]. This ensures that PCD is orchestrated both spatially and temporally. Furthermore, E governs cell death in specific tissues during both the larval and adult phases. Basically, a surge of 20E at the conclusion of the larval stage initiates metamorphosis. This process entails the removal of redundant larval tissues and the repurposing of molecular materials to form adult structures [30]. In the fat body, E93 serves as a primary mediator for 20E signaling, directing the onset of both autophagy and caspase activity. This activation is triggered via a surge of the molting hormone, 20E, transmitted through the 20E nuclear receptor complex, EcR-USP [86]. In a separate study delving into the intricate process of Drosophila salivary gland degradation, researchers have elucidated that the transcriptional upregulation of mitochondrial apoptosis factors, including Cyt-c and genes encoding the death-associated APAF1-related killer, precedes the upregulation of genes encoding the initiator and effector caspases [87]. It was reported that the accumulation of transcripts from both reaper and grim is effectively curtailed by this steroid hormone, highlighting the intricate connection between 20E and the apoptotic process [88]. As the degradation progresses, there is a noticeable and gradual increase in the presence of the Cyt-c protein and active caspase 3 in the cytoplasm. Interestingly, the Cyt-c protein exhibits colocalization with mito-GFP, a definitive marker for cytoplasmic mitochondria. Concurrently, alternations in the mitochondrial membrane potential align with the presence of Cyt-c in the cytoplasm [87]. This series of events highlights the emergence of cytoplasmic Cyt-c prior to the initiation of apoptosis in the degradation of the Drosophila salivary gland. It sheds light on an aspect of the conserved apoptotic process shared between insects and mammals. In a captivating exploration of the genetic and hormonal orchestration of neuronal death within the Drosophila central nervous system, it has been clarified that apoptosis is triggered via the genes’ expression of reaper, grim, or head involution defective. This neuronal demise is intrinsically linked to the decline in the titer of the steroid hormone 20E, a phenomenon characteristic of the metamorphic culmination. As previously mentioned, the pivotal role of E93 in upregulating downstream secondary response genes has been further underscored. This significance becomes particularly evident when its RNAi knockdown or mutation leads to the inhibition of both autophagy and caspase activity. Conversely, overexpressing E93 not only activates both processes but also more effectively counters the inhibition of autophagy compared to caspase activity under EcR overexpression. E93 inhibits PI3K-TORC1 signaling, thereby initiating autophagy [54]. In a subsequent investigation, it was observed that the removal of the tissue-specific transcription factor Fkh (Fork head) not only triggers a death response to 20E within the larval salivary glands, but it is also sufficient for this initiation. Intriguingly, this loss of Fkh is identified as a steroid-regulated event, orchestrated by the 20E-induced BR-C gene. Consequently, this sensitizes the pivotal death regulators, Hid (head involution defective) and reaper to hormonal signals. These findings accentuate the multifaceted role of the D. melanogaster FOXA orthologue Fkh, positioning it in a previously uncharted context as a crucial competence factor in the domain of steroid-governed cellular demise [89]. Furthermore, 20E inhibits the PI3K/mTOR signaling to activate the ATG1/ATG13 complex, initiating autophagosome formation in the Drosophila fat body [90].
Considering the multifaceted functions of steroid hormone 20E signaling, a more comprehensive exploration of its physiological roles and precise mechanisms in autophagy is warranted.
4.2. Regulation in B. mori
The intricate interplay between 20E signaling and autophagy in insects, particularly in Bombyx, holds great scientific importance. 20E-induced autophagy has been observed during the larval–pupal transition in various insects, with particular emphasis on Bombyx. It is noteworthy that 20E is both necessary and sufficient to induce autophagy in larval tissue, accomplished by upregulating the expression of several Atgs (autophagy-related genes), implying E-induced autophagy is a vital process in the degradation of larval tissues [91]. In Bombyx Bm-12 cells, both 20E treatment and starvation induced cell death, where autophagy preceded apoptosis. Upon exposure to 20E or starvation, BmATG8 underwent rapid cleavage, conjugating with PE (phosphatidylethanolamine) to form BmATG8-PE, followed by the cleavage of BmATG5 and BmATG6 into BmATG5-tN and BmATG6-C, respectively [2]. Studies have revealed that 20E treatment enhanced the transcription of BmV-ATPase and this effect was diminished by RNAi targeting the 20E receptor BmUsp. The hormone precursor 20E upregulated the transcription of BmV-ATPases by inducing the Bombyx transcription factor EB and facilitating its nuclear translocation. Additionally, 20E suppressed mTOR signaling, thereby promoting the transcription and assembly of BmV-ATPase subunits [92]. Notably, injecting 20E markedly induced apoptosis and elevated the gene expression of apoptotic within 6 h. When larval fat body tissues were exposed to 20E in vitro, a substantial upregulation was observed in a suite of eight genes integral to the apoptotic pathway. These genes, which include Apaf-1, Nedd2 like1, Nedd2 like2, ICE1, ICE3, ICE5, Arp, and IAP (inhibitor of apoptosis), showed heightened expression levels during critical periods of molting and pupation. This pattern of gene expression underscores the significant role of 20E in enhancing apoptotic pathways, thereby promoting PCD within the Bombyx fat body throughout the larval molting process and the transition into pupation [93]. More importantly, the 20E signaling pathway has been identified to enhance autophagy, primarily via the induction of the Atg expression and the simultaneous inhibition of the mTOR pathway. Intriguingly, early-response transcription factors to 20E, including Br-C, E74, E75, and E93, play pivotal roles in regulating this autophagy process [94]. Specifically, Bombyx E75 has been demonstrated to be essential for autophagy induction during the larval–pupal metamorphic transition [55]. The primary response gene to 20E, E93, functions through GAGA-containing motifs. It coordinates with the 20E-EcR-USP complex, playing a crucial role in the remodeling of larval tissues and the formation of adult tissues during the Bombyx larval–pupal metamorphosis [56]. Research has revealed that the expression of the E93 gene in mosquitoes is suppressed by JH and stimulated by 20E. This ecdysone-induced protein, E93, plays a crucial role in overseeing the gonadotrophic cycles of adult female mosquitoes, particularly in Aedes aegypti. It is worth noting that when E93 RNAi was applied to silence the gene prior to the initiation of the first gonadotrophic cycle, it led to a disruption in the subsequent cycle’s normal progression [95]. This result indicates that 20E can exert diverse effects on the regulation of E93 in different species. Furthermore, 20E has been shown to simulate a starvation-like state in Bombyx, primarily by diminishing food intake, thereby inducing autophagy [94]. Previous research has emphasized that BmATG5 and BmATG6 undergo cleavage into BmATG5-tN and BmATG6-C, respectively. These cleaved forms play a pivotal role in mediating the transition from autophagy to apoptosis. Notably, this shift is prompted by common stimuli, specifically 20E and starvation [2,96]. Subtle variations in BmAtg13 have been intricately associated with autophagy, activated either by 20E or via starvation. Interestingly, both the knockdown and overexpression of BmAtg13 inhibit autophagy. While the 20E treatment significantly elevates BmAtg13 gene expression, blocking 20E signaling transduction—specifically via the knockdown of BmUsp—leads to a reduction in both the gene expression and protein levels of BmAtg13. This emphasizes the pivotal role of BmAtg13 in 20E- and starvation-induced autophagy in B. mori, paving the way for more comprehensive studies [97]. It was also discovered that among the Atg genes, five exhibited primary responsiveness to 20E. This was highlighted by the identification of a distinct 20E response element within the Atg1 promoter region, an ortholog of human ULK1. The complexity of this dynamic was further unveiled when the RNAi-mediated knockdown of four key genes—Br-C, E74, HR3, and βftz-F1, all integral to the 20E-induced transcriptional cascade—resulted in diminished autophagy and the varied downregulation of Atg genes. From these findings, it was deduced that 20E’s influence is not confined to merely inhibiting TORC1 activity for autophagosome creation; it significantly boosts Atg’s genes expression, thereby promoting autophagy in the Bombyx fat body [94]. However, the exact underlying mechanism remains to be thoroughly elucidated. Furthermore, our findings illuminate that 20E adeptly dephosphorylates the histone deacetylase BmRpd3, orchestrating the deacetylation modification of ATG proteins, culminating in its nucleo-cytoplasmic translocation and the subsequent amplification of autophagy [98]. The potential parallels drawn from 20E signaling in insects may offer foundational knowledge applicable to other organisms, including vertebrates. lncRNAs (Long non-coding RNAs) have been implicated in the regulation of autophagy [99]. A recent investigation identified LNC_000560 and its putative target gene as being associated with 20E-regulated autophagy in B. mori [100]. These investigations highlight a profound connection between 20E and the regulation of both autophagy and apoptosis.
In essence, 20E signaling augments autophagy via various pathways, encompassing the induction of Atg and V-ATPases gene expression, the inhibition of mTOR pathway, and deacetylation modifications of ATG proteins.
4.3. Regulation in H. armigera
Beyond the induction of Atg gene transcription and mTOR signaling inhibition, 20E’s involvement in autophagy regulation extends even further. Its role in regulating autophagosome formation has been established and research has additionally elucidated the mechanism by which the regulation of ATG12-ATG5 conjugation influences autophagy. Within the context of H. armigera, 20E adeptly orchestrates the ATG12–ATG5 conjugation in a manner that is both concentration and time sensitive, thereby promoting autophagy and apoptosis in the insect midgut, a phenomenon that will be elaborated upon subsequently [78]. In a detailed study, it was found that during lepidopteran metamorphosis, the roles of CTSD (cathepsin D), a lysosomal aspartic protease, in apoptosis and cell proliferation are intricately regulated. This is achieved via its varied tissue-specific expression and the precise autophagy-mediated maturation process. Specifically, CTSD boosts cell proliferation by releasing its pro-enzyme as an external ligand, while its mature intracellular form encourages apoptosis. This balance is further emphasized by the 20E, which increases CTSD expression, and the orchestrated autophagy process that facilitates its maturation, ultimately culminating in the activation of caspase 3 and the promotion of apoptosis [101]. Recent findings have underscored the central role of 20E in reshaping amino acid metabolism. A novel study indicates that during metamorphic development, 20E intricately orchestrates amino acid metabolism. Notably, arginine, α-KG (alpha-ketoglutarate), and Glu (glutamate) are identified as signature metabolites for feeding larvae, migrating larvae, and pupae stages, respectively. This metamorphic shift involves a 20E-mediated reduction in arginine levels, achieved by inhibiting arginosuccinate synthetase and amplifying arginase expression. In the larval midgut, Glu transforms into α-KG via GDH (glutamate dehydrogenase), a conversion restrained by 20E. Fascinatingly, in the pupal fat body, the process is reversed: α-KG is reverted to Glu by GDH-like, an enzyme reinforced by 20E. These findings underscore 20E’s significant influence in the metabolic reconfiguration, ensuring a seamless insect metamorphosis [6]. While the steroid hormone 20E is known to increase the expression of calcium release-activated calcium channel modulator 1, thereby inducing apoptosis in the midgut of H. armigera, a contrasting study offers a more nuanced perspective, illustrating that insulin and 20E are involved in a delicate antagonism in modulating PDK1 (phosphoinositide-dependent kinase-1) expression during insect pupation. This delicate balance is evident, as the dsRNA-mediated PDK1 knockdown in H. armigera larvae not only postpones pupation, resulting in diminutive pupae, but also attenuates Akt/protein kinase B expression while enhancing FoxO (Forkhead Box O) expression, suggesting that the PDK1 knockdown impedes midgut remodeling and reduces 20E levels in the larvae, a phenomenon consistent with the shared regulatory mechanisms of autophagy and apoptosis [102,103]. Probing further into the metabolic complexities of holometabolous insect development, a subsequent study suggests a synchronized interplay between glycolysis and autophagy. This balance is orchestrated via the intricate interrelation of the 20E and insulin signaling pathways. Specifically, insulin accelerates glycolysis and cell proliferation by phosphorylating PGK1. In contrast, 20E employs PTEN to dephosphorylate PGK1, effectively inhibiting glycolysis. This discovery sheds additional light on the subtle role of 20E in regulating autophagy [80]. In the H. armigera epidermal cell line, STIM1 (stromal interacting molecule 1) regulates store-operated calcium entry, with 20E-induced phosphorylation at Ser-485 via PKC (protein kinase C) playing a crucial role in calcium ion influx, STIM1 clustering, and its interaction with Orai1 and subsequent induction of apoptosis, further confirming the role of 20E in regulating apoptosis [104]. In contrast, the activation of the EcRB1/USP1 transcription complex by 20E led to an upregulation of PKCδ expression in various tissues during metamorphosis, while silencing PKCδ hindered the larval–pupal transition, inhibited tissues from undergoing autophay and apoptosis, and resulted in a decreased expression of the transcription factor Brz-7, as well as the apoptotic executors caspase-3 and caspase-6 [105]. This mechanism strongly suggests that the phosphorylation of PKCδ at the threonine residue located at position 1343 was critical for its proapoptotic activity. While 20E plays different roles in different species, leading to diverse impacts, the results of its actions are closely interlinked. Particularly noteworthy is the upregulation of prodeath serine/threonine protein kinase expression for PCD by the steroid hormone 20E, with studies illuminating that in the lepidopteran insect H. armigera, the knockdown of prodeath S/TK—achieved by injecting dsRNA into the larval hemocoel—not only prevents 20E-induced metamorphosis and PCD, but also attenuates the expression of a number of genes integral to the PCD and 20E signaling pathway, and intriguingly, 20E amplifies prodeath S/TK expression via its nuclear receptors EcR-B1 and USP1, further solidifying the pivotal role that 20E plays in autophagy and apoptosis [106].
4.4. Regulation in the Others
While D. melanogaster exhibits a notably specialized developmental trajectory, suggesting that its apoptotic regulatory mechanisms may not be emblematic of the broader insect class, insights from B. germanica (Blattella germanica) offer a contrasting perspective. This species is known to possess two inhibitors of apoptosis proteins, with BgIAP1 being indispensable for maintaining tissue vitality, particularly in the prothoracic gland, throughout nymphal maturation. A closer look reveals that the orchestrated degeneration of the prothoracic gland is governed by an intricate 20E-initiated cascade of nuclear receptors, culminating in the potent activation of the death-inducing Fushi tarazu factor 1 as the nymph transitions to adulthood. Complementing this understanding, it is noteworthy that the JH acts as a formidable safeguard, effectively preventing the degeneration of the prothoracic gland [107].
In the honeybee, Apis mellifera Ligustica, a study has shown that 20E triggers apoptosis in the larval fat body during pupation, a process that is mitigated via the RNAi-mediated knockdown of ECR genes. The peak expression of the ECR gene occurs in seven-day-old larvae, followed by a gradual decline during the pupal stage, highlighting 20E’s decisive role in the regulation of pupation via apoptosis [108]. In Heortia vitessoides Moore, research has uncovered expression patterns of HvATG8, with a notable upregulation in the prepupal, pupal, and adult stages, as well as in the larval midgut and adult abdomen. Crucially, this suppression of HvATG8 led to a simultaneous decrease in HvATG3, revealing a functional interplay between these genes. Additionally, HvATG8 expression demonstrated responsiveness to 20E, as well as environmental stressors like starvation and extreme temperatures, indicating its pivotal role in stress response and developmental regulation [109]. During the pupal stage, the orchestrated degeneration of larval cells occurs via autophagic cell death, a process marked by caspase-3 activity and regulated by 20E via the ecdysone receptor EcR-B1 pathway. This is further substantiated via the increased activity of acid phosphatase and the simultaneous upregulation of ATG6 and ATG8, which align with autophagosome formation in the fat body of Galleria, underscoring autophagy’s pivotal role in developmental remodeling [110].
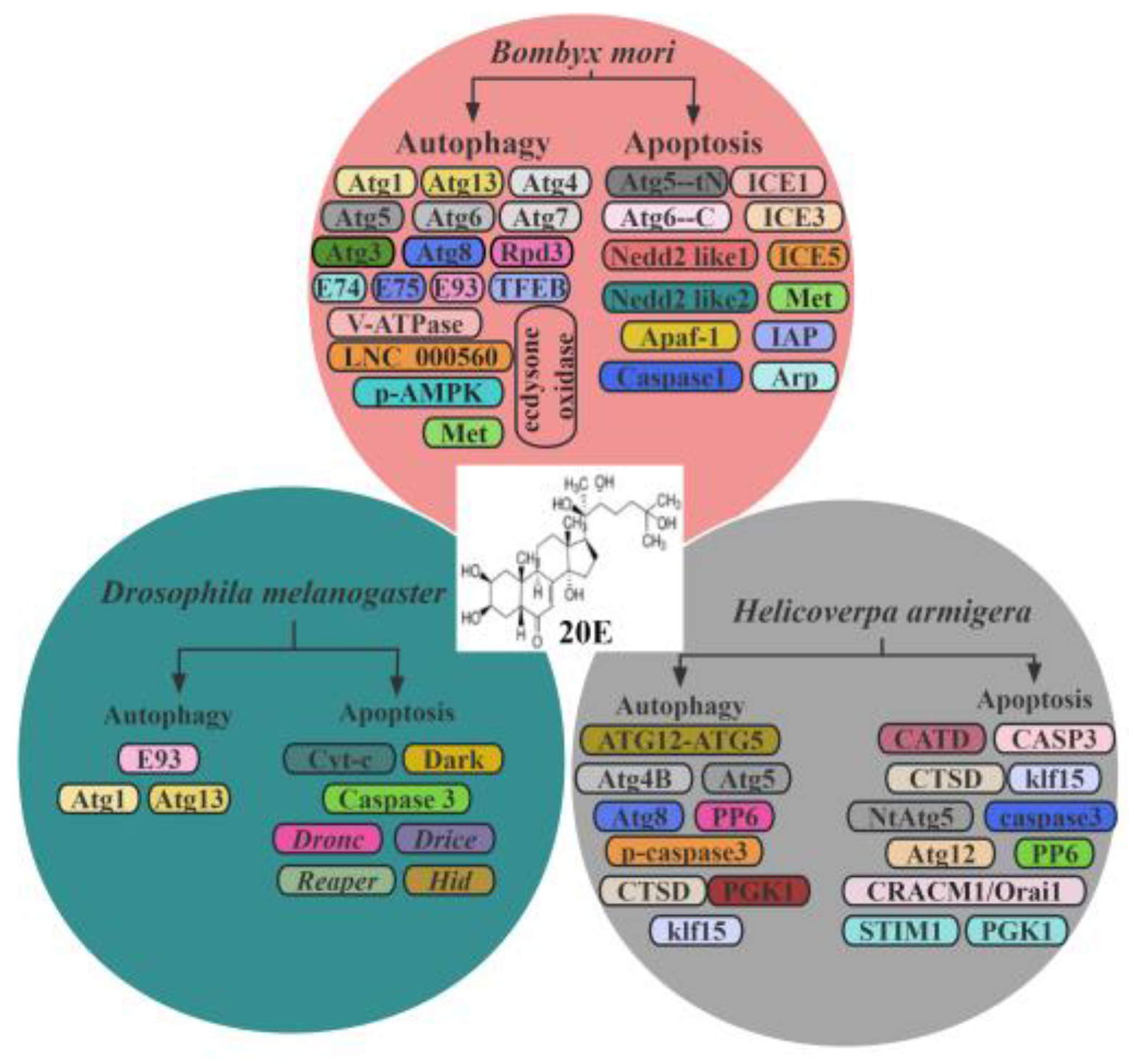
Taken together, the regulation of autophagy and apoptosis by 20E represents a critical point in the cellular physiology of the insects (Table 2). These processes, fundamental to cellular homeostasis and development, are intricately modulated by 20E to ensure that cells are either rejuvenated via autophagy or undergo programmed death via apoptosis at the appropriate developmental stages.
Table 2.
20E regulated autophagy and apoptosis in insects.
| PCD-Related Enzymes | Species | Autophagy/Apoptosis | Up/Downregulation | Pathway | Associated Protein/Factors |
|---|---|---|---|---|---|
| Caspase | D. melanogaster | Apoptosis | Up [54] | PI3K-TORC1 | Caspase-9, Caspase 3, Caspase-6 |
| AMPK-PP2A axis | D. melanogaster | Autophagy | Down [15] | Insulin/IGF | PP2A, AKT |
| ATG1/ATG13 complex | D. melanogaster | Autophagy | Down [90] | PI3K/mTOR | PTEN |
| BmADK | B. mori | Autophagy and apoptosis | Down [61] | \ | ATG-8, Caspase-9, Ec-R, E74A, Br-C |
| BmV-ATPase | B. mori | Autophagy and apoptosis | Up [92] | mTOR | TFEB, BmATG8, BmATG5, BmATG6 |
| 20E-EcR-USP complex | B. mori | Autophagy and apoptosis | Down [94] | BECN1/ATG6-PIK3C3/Vps34 (catalytic subunit of class III PtdIns3K) | Br-C, E74, E75, E93 |
| BIGFLP | B. mori | Autophagy | Up [60] | Insulin/IGF | Insulin-like peptides |
| HDACs | B. mori | Autophagy | Up [98] | BmRpd3/HsHDAC1 | mTORC1 |
| HaP60 | H. armigera | Apoptosis | Up [62] | Insulin/IGF | ILPs, HaFOXO |
| PDK1 and FoxO | H. armigera | Apoptosis | Up [103] | \ | Insulin, calcium channel modulator 1 |
| CTSD/CathD/CATD | H. armigera | Apoptosis | Up [101] | Activation of MAPK1/ERK2 -MAPK3/ERK1 and AKT/protein kinase B | Ccaspase 3 and Caspase 7 |
| PGK1 | H. armigera | Autophagy | Up [80] | \ | ARD1, PTEN |
| PKCδ | H. armigera | Apoptosis | Up [105] | EcRB1/USP1 | Brz-7, Caspase-3, Caspase-6 |
5. Discussion
In the realm of insect development and metamorphosis, the intricate interplay between 20E and PCD presents a maze of unresolved questions. The role of 20E, as a primary orchestrator of insect growth, raises fundamental questions about its ability to modulate PCD in different tissues and developmental stages. Initially, insect metamorphosis is a remarkable process characterized by extensive PCD, a phenomenon which might not be as pervasive in other organisms. The intricacy of insect metamorphosis necessitates this cell death, ensuring redundant tissues are eliminated to facilitate the emergence of a mature individual. Moreover, the hormone 20E in insects serves as a vital regulator of this PCD. It acts as a guide, directing cells towards their demise by modulating gene expression and internal biochemical processes. Such a regulatory mechanism, and the prominence of this hormone, might not be mirrored or hold equivalent significance in other organisms. Subsequently, the occurrence of PCD in insects is intricately timed, coinciding with specific developmental stages and tissues. This is particularly evident during phases like metamorphosis and tissue remodeling. Contrastingly, the patterns of cell death in other organisms may diverge, given that different species have their own unique developmental trajectories and life cycle demands. Collectively, while there might be overarching similarities in the regulation of PCD across various organisms, insects exhibit unique characteristics and mechanisms. These are intrinsically tied to their biological attributes and developmental paradigms. One wonders whether the cellular response to 20E is a direct, immediate one, or whether it is a more complicated process that unfolds in a series of nuanced steps, revealing a complex choreography of cellular events. Beyond the well-established EnR pathway, there is speculation about potential alternative pathways—hidden alleys and byways, through which 20E might exert its influence on PCD.
The emergence of pesticide-resistant insect strains raises another pressing question: To what extent are alterations in 20E signaling or perturbations in PCD pathways responsible for this burgeoning resistance? Such resistance not only poses challenges for agriculture and pest management, but also underscores the need to understand the molecular and physiological intricacies that underlie these changes.
From an ecological perspective, the implications of 20E-induced PCD go beyond the individual insect. What are the cascading effects on ecosystems when 20E pathways are altered, either naturally or via human intervention? The potential impacts on food webs, species interactions, and ecosystem stability warrant thorough investigation. Finally, the evolutionary perspective offers a rich tapestry of insights. The 20E-PCD relationship, which has persisted across evolutionary timescales, prompts reflection on its significance in the insect evolutionary narrative. Does this relationship represent a conserved developmental mechanism that has been essential for insect success? Or does it suggest a dynamic interplay, constantly reshaped by ecological pressures and evolutionary forces? Beyond its established roles in autophagy and apoptosis, how might 20E influence other forms of PCD, such as necroptosis or pyroptosis? How does the regulation of PCD in insects compare to that in other organisms? Could 20E also play a pivotal role in the insect reproductive processes? If so, what are the underlying mechanisms through which these hormones operate? Are there specific insect species or developmental stags where 20E’s influence on PCD mechanisms other than autophagy and apoptosis is more pronounced? Combining these questions, it becomes evident that the 20E-PCD axis in insects transcends being a mere biological mechanism; rather, it represents a multifaceted subject intertwining ecology, evolution, and applied science. Addressing these questions is not only pivotal for advancing our understanding of insect biology, but also for addressing the challenges and opportunities they present in a rapidly changing world.
In summary, a comprehensive understanding of the 20E signaling landscape, highlighting its important role in mediating autophagy and apoptosis in insects and clearly revealing the mechanisms behind this action, will provide new avenues in our research (Figure 2) [111]. By delving into the 20E-autophagy/apoptosis axis, we gain profound insights not only into the complex interconnections within endocrine networks, but also into the innovative adaptive strategies insects have evolved in response to environmental challenges.
Author Contributions
W.W., L.L., H.L. and Q.Z. conceptualized the manuscript; L.L. and Q.Z. wrote the manuscript text; L.L., H.L. and W.W. reviewed the text; L.L., Q.Z. and J.H. designed the figures and tables; W.W. and H.L. obtained funding and supervised. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Science and Technology Projects in Guangzhou (Grant #2023A04J0862), funding by high-level talent introduction into Guangdong Pharmaceuical University (Grant #51304043013), and also funding by Guangdong Province College Students Innovation and Entrepreneurship Training Program (Grant #202210573047).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 20E | 20-Hydroxyecdysone |
| PCD | Programmed cell death |
| E | Ecdysone |
| D. melanogaster | Drosophila melanogaster |
| B. mori | Bombyx mori |
| H. Armigera | Helicoverpa armigera |
| B. germanica | Blattella germanica |
| EnR | ecdysone receptor |
| JH | juvenile hormone |
| Hemimetabola | Dictyoptera to Hemiptera |
| USP | ultraspiracle protein |
| EcR-B1 | ecdysone receptor B1 isoform |
| USP1 | ubiquitin specific peptidase 1 |
| PTTH | Prothoracicotropic Hormone |
| Insulin/IIS | Insulin-like growth factor receptor signaling |
| IAP | inhibitor of apoptosis |
| AMPK | AMP-activated protein kinase |
| PP2A | protein phosphatase 2A |
| AKT | protein kinase B |
| Br-C | Broad-Complex |
| E74 | Ecdysone-induced protein 74 |
| E75 | Ecdysone-induced protein 75 |
| E93 | Ecdysone-induced protein 93 |
| BIGFLP | Bombyx imp-L2 gene-enhancer flippase protein |
| HaP60 | H. armigera P60 |
| Klf15 | Krüppel-like factor |
| PTEN | phosphatase and tensin homolog |
| Cyt-c | cytochrome C |
| lncRNAs | Long non-coding RNAs |
| Hid | head involution defective |
| Fkh | Fork head |
| CTSD | cathepsin D |
| FoxO | Forkhead Box O |
| Atgs | autophagy related genes |
| PE | phosphatidylethanolamine |
| α-KG | alpha-ketoglutarate |
| Glu | glutamate |
| STIM1 | stromal interacting molecule 1 |
| PKC | protein kinase C |
| GDH | glutamate dehydrogenase |
| PDK1 | phosphoinositide-dependent kinase-1 |
References
- Kaplanis, J.N.; Thompson, M.J.; Robbins, W.E.; Bryce, B.M. Insect hormones: Alpha ecdysone and 20-hydroxyecdysone in bracken fern. Science 1967, 157, 1436–1438. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Tian, L.; Guo, X.; Li, K.; Li, J.; Deng, X.; Li, Q.; Xia, Q.; Zhong, Y.; Huang, Z.; et al. BmATG5 and BmATG6 mediate apoptosis following autophagy induced by 20-hydroxyecdysone or starvation. Autophagy 2016, 12, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Werling, K.; Shaw, W.R.; Itoe, M.A.; Westervelt, K.A.; Marcenac, P.; Paton, D.G.; Peng, D.; Singh, N.; Smidler, A.L.; South, A.; et al. Steroid Hormone Function Controls Non-competitive Plasmodium Development in Anopheles. Cell 2019, 177, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Chen, Z.; Wen, J.; Jia, Q. Sterol Regulation of Development and 20-Hydroxyecdysone Biosynthetic and Signaling Genes in Drosophila melanogaster. Cells 2023, 12, 1739. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, H.; Yamada, R. Ecdysteroids during early embryonic development in silkworm Bombyx mori: Metabolism and functions. Zool. Sci. 2004, 21, 503–516. [Google Scholar] [CrossRef]
- Wang, X.P.; Sun, S.P.; Li, Y.X.; Wang, L.; Dong, D.J.; Wang, J.X.; Zhao, X.F. 20-hydroxyecdysone reprograms amino acid metabolism to support the metamorphic development of Helicoverpa armigera. Cell Rep. 2023, 42, 112644. [Google Scholar] [CrossRef]
- Denton, D.; Aung-Htut, M.T.; Kumar, S. Developmentally programmed cell death in Drosophila. Biochim. Biophys. Acta 2013, 1833, 3499–3506. [Google Scholar] [CrossRef]
- He, H.J.; Hou, L.; Wang, J.X.; Zhao, X.F. The apoptosis inhibitor survivin prevents insect midgut from cell death during postembryonic development. Mol. Biol. Rep. 2012, 39, 1691–1699. [Google Scholar] [CrossRef] [PubMed]
- Tower, J. Programmed cell death in aging. Ageing Res. Rev. 2015, 23, 90–100. [Google Scholar] [CrossRef]
- Yamamoto, H.; Matsui, T. Molecular mechanisms of macroautophagy, microautophagy, and chaperone-mediated autophagy. J. Nippon Med. Sch. 2023. [Google Scholar] [CrossRef] [PubMed]
- Sorice, M. Crosstalk of Autophagy and Apoptosis. Cells 2022, 11, 1479. [Google Scholar] [CrossRef]
- Hsu, H.W.; Chiu, M.C.; Shih, C.J.; Matsuura, K.; Yang, C.S. Apoptosis as a primary defense mechanism in response to viral infection in invasive fire ant Solenopsis invicta. Virology 2019, 531, 255–259. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, Q.; Zhou, X.L.; Zhu, Y.; Dong, Z.Q.; Chen, P.; Pan, M.H.; Lu, C. Bombyx mori Nuclear Polyhedrosis Virus (BmNPV) Induces Host Cell Autophagy to Benefit Infection. Viruses 2017, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Rost-Roszkowska, M.M.; Poprawa, I.; Chachulska-Zymełka, A. Apoptosis and autophagy in the midgut epithelium of Acheta domesticus (Insecta, Orthoptera, Gryllidae). Zool. Sci. 2010, 27, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Zhou, S.; Liu, S.; Li, K.; Zhao, H.; Long, S.; Liu, H.; Xie, Y.; Su, Y.; Yu, F.; et al. The AMPK-PP2A axis in insect fat body is activated by 20-hydroxyecdysone to antagonize insulin/IGF signaling and restrict growth rate. Proc. Natl. Acad. Sci. USA 2020, 117, 9292–9301. [Google Scholar] [CrossRef]
- Taylor, D.M.; Olds, C.L.; Haney, R.S.; Torrevillas, B.K.; Luckhart, S. Comprehensive and Durable Modulation of Growth, Development, Lifespan and Fecundity in Anopheles stephensi Following Larval Treatment with the Stress Signaling Molecule and Novel Antimalarial Abscisic Acid. Front. Microbiol. 2019, 10, 3024. [Google Scholar] [CrossRef]
- Li, Y.B.; Li, X.R.; Yang, T.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone promotes switching from autophagy to apoptosis by increasing intracellular calcium levels. Insect Biochem. Mol. Biol. 2016, 79, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wu, X.; Zhou, L.; He, T.; Yin, Q.; Liu, S. 20-Hydroxyecdysone-responsive microRNAs of insects. RNA Biol. 2020, 17, 1454–1471. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory Pathways Controlling Female Insect Reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Fu, B.; Wang, C.; Liu, J.; Tang, Y.; Zhang, W.; Zhu, J.; Li, Y.; Pan, Q.; Liu, F. The role of 20E biosynthesis relative gene Shadow in the reproduction of the predatory mirid bug, Cyrtorhinus lividipennis (Hemiptera: Miridae). Arch. Insect Biochem. Physiol. 2022, 109, e21854. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.; Kim, H.Y.; Chen, X.; Shen, W.; Jang, J.S.; Stein, S.N.; Cormier, O.; Pereira, L.; Shih, C.; Krieger, C.; et al. 20-hydroxyecdysone (20E) signaling regulates amnioserosa morphogenesis during Drosophila dorsal closure: EcR modulates gene expression in a complex with the AP-1 subunit, Jun. Biol. Open 2021, 10, bio058605. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Sun, X.; Jin, X.Y. 20E-regulated USP expression and phosphorylation in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2003, 33, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Shimizu, S.; Fujiwara, H.; Sakurai, S.; Iwami, M. EcR expression in the prothoracicotropic hormone-producing neurosecretory cells of the Bombyx mori brain. FEBS J. 2006, 273, 3861–3868. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Hiruma, K.; Hu, X.; Jindra, M.; Riddiford, L.M. Activation of a delayed-early gene encoding MHR3 by the ecdysone receptor heterodimer EcR-B1-USP-1 but not by EcR-B1-USP-2. Mol. Cell. Biol. 1999, 19, 4897–4906. [Google Scholar] [CrossRef] [PubMed]
- He, H.J.; Wang, Q.; Zheng, W.W.; Wang, J.X.; Song, Q.S.; Zhao, X.F. Function of nuclear transport factor 2 and Ran in the 20E signal transduction pathway in the cotton bollworm, Helicoverpa armigera. BMC Mol. Cell Biol. 2010, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Hiruma, K.; Jindra, M.; Shinoda, T.; Segraves, W.A.; Malone, F.; Riddiford, L.M. Regulation of the transcription factor E75 by 20-hydroxyecdysone and juvenile hormone in the epidermis of the tobacco hornworm, Manduca sexta, during larval molting and metamorphosis. Dev. Biol. 1998, 193, 127–138. [Google Scholar] [CrossRef]
- Dong, D.J.; Wang, J.X.; Zhao, X.F. A eukaryotic initiation factor 5C is upregulated during metamorphosis in the cotton bollworm, Helicoverpa armigera. BMC Dev. Biol. 2009, 9, 19. [Google Scholar] [CrossRef]
- Kefi, M.; Balabanidou, V.; Douris, V.; Lycett, G.; Feyereisen, R.; Vontas, J. Two functionally distinct CYP4G genes of Anopheles gambiae contribute to cuticular hydrocarbon biosynthesis. Insect Biochem. Mol. Biol. 2019, 110, 52–59. [Google Scholar] [CrossRef]
- Cruz, J.; Martín, D.; Franch-Marro, X. Egfr Signaling Is a Major Regulator of Ecdysone Biosynthesis in the Drosophila Prothoracic Gland. Curr. Biol. 2020, 30, 1547–1554. [Google Scholar] [CrossRef]
- Engelmann, F. 20-hydroxyecdysone, what it can do. Science 1971, 174, 1041. [Google Scholar] [CrossRef]
- Temirgaziyev, B.S.; Kučáková, K.; Baizhigit, Y.A.; Jurášek, M.; Džubák, P.; Hajdúch, M.; Dolenský, B.; Drašar, P.B.; Tuleuov, B.I.; Adekenov, S.M. Bioavailability and structural study of 20-hydroxyecdysone complexes with cyclodextrins. Steroids 2019, 147, 37–41. [Google Scholar] [CrossRef]
- Horike, N.; Sonobe, H. Ecdysone 20-monooxygenase in eggs of the silkworm, Bombyx mori: Enzymatic properties and developmental changes. Arch. Insect Biochem. Physiol. 1999, 41, 9–17. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Yamanaka, N.; Gilbert, L.I.; O’Connor, M.B. The insect neuropeptide PTTH activates receptor tyrosine kinase torso to initiate metamorphosis. Science 2009, 326, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Ou, Q.; King-Jones, K. What goes up must come down: Transcription factors have their say in making ecdysone pulses. Curr. Top. Dev. Biol. 2013, 103, 35–71. [Google Scholar]
- Erion, R.; Sehgal, A. Regulation of insect behavior via the insulin-signaling pathway. Front. Physiol. 2013, 4, 353. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, G.; Yu, L.; Du, S.; Jiang, D.; Chu, X.; Wang, K.; Wu, S.; Wang, R.; Zhang, F.; et al. Temperature and metal ions regulate larval diapause termination via the 20-hydroxyecdysone and juvenile hormone pathways in Monochamus alternatus. Pest Manag. Sci. 2023, 79, 437–446. [Google Scholar] [CrossRef]
- Guo, S.; Tian, Z.; Wu, Q.W.; King-Jones, K.; Liu, W.; Zhu, F.; Wang, X.P. Steroid hormone ecdysone deficiency stimulates preparation for photoperiodic reproductive diapause. PLoS Genet. 2021, 17, e1009352. [Google Scholar] [CrossRef]
- Rauschenbach, I.Y.; Chentsova, N.A.; Alekseev, A.A.; Gruntenko, N.E.; Adonyeva, N.V.; Karpova, E.K.; Komarova, T.N.; Vasiliev, V.G.; Bownes, M. Dopamine and octopamine regulate 20-hydroxyecdysone level in vivo in Drosophila. Arch. Insect Biochem. Physiol. 2007, 65, 95–102. [Google Scholar] [CrossRef]
- Satake, S.; Masumura, M.; Ishizaki, H.; Nagata, K.; Kataoka, H.; Suzuki, A.; Mizoguchi, A. Bombyxin, an insulin-related peptide of insects, reduces the major storage carbohydrates in the silkworm Bombyx mori. Comp. Biochem. Phys. B 1997, 118, 349–357. [Google Scholar] [CrossRef]
- Iga, M.; Kataoka, H. Recent studies on insect hormone metabolic pathways mediated by cytochrome P450 enzymes. Biol. Pharm. Bull. 2012, 35, 838–843. [Google Scholar] [CrossRef]
- Guittard, E.; Blais, C.; Maria, A.; Parvy, J.P.; Pasricha, S.; Lumb, C.; Lafont, R.; Daborn, P.J.; Dauphin-Villemant, C. CYP18A1, a key enzyme of Drosophila steroid hormone inactivation, is essential for metamorphosis. Dev. Biol. 2011, 349, 35–45. [Google Scholar] [CrossRef]
- Shimell, M.; O’Connor, M.B. The cytochrome P450 Cyp6t3 is not required for ecdysone biosynthesis in Drosophila melanogaster. MicroPubl. Biol. 2022, 2022. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Gilbert, L.I. Daphnia Halloween genes that encode cytochrome P450s mediating the synthesis of the arthropod molting hormone: Evolutionary implications. BMC Evol. Biol. 2008, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, S.; Jia, Q.; Wu, L.; Yuan, D.; Li, E.Y.; Feng, Q.; Wang, G.; Palli, S.R.; Wang, J.; et al. Juvenile hormone membrane signaling phosphorylates USP and thus potentiates 20-hydroxyecdysone action in Drosophila. Sci. Bull. 2022, 67, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Michalik, J.; Chojnicka, B.; Cymborowski, B. Vitellogenesis in virgin and mated females of the mealworm beetle, Tenebrio molitor. Acta Biochim. Pol. 1996, 43, 623–631. [Google Scholar] [CrossRef]
- Panaitof, S.C.; Scott, M.P. Effect of juvenile hormone on vitellogenin gene expression in the fat body of burying beetles, Nicrophorus orbicollis. Arch. Insect Biochem. Physiol. 2006, 63, 82–91. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory Mechanisms of Vitellogenesis in Insects. Front. Cell Dev. Biol. 2020, 8, 593613. [Google Scholar] [CrossRef]
- Zhu, L.; Yin, T.Y.; Sun, D.; Liu, W.; Zhu, F.; Lei, C.L.; Wang, X.P. Juvenile hormone regulates the differential expression of putative juvenile hormone esterases via methoprene-tolerant in non-diapause-destined and diapause-destined adult female beetle. Gene 2017, 627, 373–378. [Google Scholar] [CrossRef]
- Wyatt, G.R.; Davey, K.G. Cellular and Molecular Actions of Juvenile Hormone. II. Roles of Juvenile Hormone in Adult Insects. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 1996; Volume 26, pp. 1–155. [Google Scholar]
- Mpakou, V.E.; Velentzas, A.D.; Velentzas, P.D.; Margaritis, L.H.; Stravopodis, D.J.; Papassideri, I.S. Programmed cell death of the ovarian nurse cells during oogenesis of the ladybird beetle Adalia bipunctata (Coleoptera: Coccinellidae). Dev. Growth Differ. 2011, 53, 804–815. [Google Scholar] [CrossRef]
- Liu, S.; Li, K.; Gao, Y.; Liu, X.; Chen, W.; Ge, W.; Feng, Q.; Palli, S.R.; Li, S. Antagonistic actions of juvenile hormone and 20-hydroxyecdysone within the ring gland determine developmental transitions in Drosophila. Proc. Natl. Acad. Sci. USA 2018, 115, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Petryk, A.; Warren, J.T.; Marqués, G.; Jarcho, M.P.; Gilbert, L.I.; Kahler, J.; Parvy, J.P.; Li, Y.; Dauphin-Villemant, C.; O’Connor, M.B. Shade is the Drosophila P450 enzyme that mediates the hydroxylation of ecdysone to the steroid insect molting hormone 20-hydroxyecdysone. Proc. Natl. Acad. Sci. USA 2003, 100, 13773–13778. [Google Scholar] [CrossRef]
- Zheng, J.; Tian, K.; Yuan, Y.; Li, M.; Qiu, X. Identification and expression patterns of Halloween genes encoding cytochrome P450s involved in ecdysteroid biosynthesis in the cotton bollworm Helicoverpa armigera. B Entomol. Res. 2017, 107, 85–95. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Li, S. E93 predominantly transduces 20-hydroxyecdysone signaling to induce autophagy and caspase activity in Drosophila fat body. Insect Biochem. Mol. Biol. 2014, 45, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S.H.; Palli, S.R.; Cao, Y.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E75 Isoforms Mediate Steroidogenesis Autoregulation and Regulate Developmental Timing in Bombyx. J. Biol. Chem. 2016, 291, 18163–18175. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S.R.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. J Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [PubMed]
- Tohidi-Esfahani, D.; Graham, L.D.; Hannan, G.N.; Simpson, A.M.; Hill, R.J. An ecdysone receptor from the pentatomomorphan, Nezara viridula, shows similar affinities for moulting hormones makisterone A and 20-hydroxyecdysone. Insect Biochem. Mol. Biol. 2011, 41, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jia, Q.; Tettamanti, G.; Li, S. Balancing crosstalk between 20-hydroxyecdysone-induced autophagy and caspase activity in the fat body during Drosophila larval-prepupal transition. Insect Biochem. Mol. Biol. 2013, 43, 1068–1078. [Google Scholar] [CrossRef]
- Tian, L.; Guo, E.; Wang, S.; Liu, S.; Jiang, R.J.; Cao, Y.; Ling, E.; Li, S. Developmental regulation of glycolysis by 20-hydroxyecdysone and juvenile hormone in fat body tissues of the silkworm, Bombyx mori. J. Mol. Cell Biol. 2010, 2, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Fujinaga, D.; Kohmura, Y.; Okamoto, N.; Kataoka, H.; Mizoguchi, A. Insulin-like growth factor (IGF)-like peptide and 20-hydroxyecdysone regulate the growth and development of the male genital disk through different mechanisms in the silkmoth, Bombyx mori. Insect Biochem. Mol. Biol. 2017, 87, 35–44. [Google Scholar] [CrossRef]
- Song, K.; Li, Y.; He, H.; Liu, L.; Zhao, P.; Xia, Q.; Wang, Y. A Novel Adenosine Kinase from Bombyx mori: Enzymatic Activity, Structure, and Biological Function. Int. J. Mol. Sci. 2019, 20, 3732. [Google Scholar] [CrossRef]
- Di, Y.Q.; Zhao, Y.M.; Jin, K.Y.; Zhao, X.F. Subunit P60 of phosphatidylinositol 3-kinase promotes cell proliferation or apoptosis depending on its phosphorylation status. PLoS Genet. 2021, 17, e1009514. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Xu, Q.Y.; Fu, K.Y.; Guo, W.C.; Li, G.Q. RNA interference against the putative insulin receptor substrate gene chico affects metamorphosis in Leptinotarsa decemlineata. Insect Biochem. Mol. Biol. 2018, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bhaskara, S.; Chandrasekharan, M.B.; Ganguly, R. Caffeine induction of Cyp6a2 and Cyp6a8 genes of Drosophila melanogaster is modulated by cAMP and D-JUN protein levels. Gene 2008, 415, 49–59. [Google Scholar] [CrossRef]
- Jones, R.T.; Bakker, S.E.; Stone, D.; Shuttleworth, S.N.; Boundy, S.; McCart, C.; Daborn, P.J.; Ffrench-Constant, R.H.; van den Elsen, J.M. Homology modelling of Drosophila cytochrome P450 enzymes associated with insecticide resistance. Pest Manag. Sci. 2010, 66, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.I. Halloween genes encode P450 enzymes that mediate steroid hormone biosynthesis in Drosophila melanogaster. Mol. Cell. Endocrinol. 2004, 215, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Segraves, W.A.; Hogness, D.S. The E75 ecdysone-inducible gene responsible for the 75B early puff in Drosophila encodes two new members of the steroid receptor superfamily. Gene Dev. 1990, 4, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, T.J.; Dubrovskaya, V.A.; Jannat, H.; Maughan, B.; Dubrovsky, E.B. Hormonal regulation of the E75 gene in Drosophila: Identifying functional regulatory elements through computational and biological analysis. J. Mol. Biol. 2009, 387, 794–808. [Google Scholar] [CrossRef]
- Rewitz, K.F.; Yamanaka, N.; O’Connor, M.B. Steroid hormone inactivation is required during the juvenile-adult transition in Drosophila. Dev. Cell 2010, 19, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mao, T.; Wang, H.; Lu, Z.; Qu, J.; Fang, Y.; Chen, J.; Li, M.; Cheng, X.; Hu, J.; et al. The CncC/keap1 pathway is activated in high temperature-induced metamorphosis and mediates the expression of Cyp450 genes in silkworm, Bombyx mori. Biochem. Biophys. Res. Commun. 2019, 514, 1045–1050. [Google Scholar] [CrossRef]
- Li, B.; Zhang, H.; Ni, M.; Wang, B.B.; Li, F.C.; Xu, K.Z.; Shen, W.D.; Xia, Q.Y.; Zhao, P. Identification and characterization of six cytochrome P450 genes belonging to CYP4 and CYP6 gene families in the silkworm, Bombyx mori. Mol. Biol. Rep. 2014, 41, 5135–5146. [Google Scholar] [CrossRef]
- Li, Z.; Ge, X.; Ling, L.; Zeng, B.; Xu, J.; Aslam, A.F.; You, L.; Palli, S.R.; Huang, Y.; Tan, A. CYP18A1 regulates tissue-specific steroid hormone inactivation in Bombyx mori. Insect Biochem. Mol. Biol. 2014, 54, 33–41. [Google Scholar] [CrossRef]
- Li, K.; Guo, E.; Hossain, M.S.; Li, Q.; Cao, Y.; Tian, L.; Deng, X.; Li, S. Bombyx E75 isoforms display stage- and tissue-specific responses to 20-hydroxyecdysone. Sci. Rep. 2015, 5, 12114. [Google Scholar] [CrossRef] [PubMed]
- Reza, A.M.; Kanamori, Y.; Shinoda, T.; Shimura, S.; Mita, K.; Nakahara, Y.; Kiuchi, M.; Kamimura, M. Hormonal control of a metamorphosis-specific transcriptional factor Broad-Complex in silkworm. Comp. Biochem. Phys. B 2004, 139, 753–761. [Google Scholar] [CrossRef]
- Sekimoto, T.; Iwami, M.; Sakurai, S. Coordinate responses of transcription factors to ecdysone during programmed cell death in the anterior silk gland of the silkworm, Bombyx mori. Insect Mol. Biol. 2006, 15, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Yang, X.; Bu, Z.; Li, X.; Zhang, Z.; Sun, W. Identification and Characterization of Genes Involved in Ecdysteroid Esterification Pathway Contributing to the High 20-Hydroxyecdysone Resistance of Helicoverpa armigera. Front. Physiol. 2020, 11, 508. [Google Scholar] [CrossRef] [PubMed]
- Winbush, A.; Weeks, J.C. Steroid-triggered, cell-autonomous death of a Drosophila motoneuron during metamorphosis. Neural Dev. 2011, 6, 15. [Google Scholar] [CrossRef]
- Li, Y.B.; Yang, T.; Wang, J.X.; Zhao, X.F. The Steroid Hormone 20-Hydroxyecdysone Regulates the Conjugation of Autophagy-Related Proteins 12 and 5 in a Concentration and Time-Dependent Manner to Promote Insect Midgut Programmed Cell Death. Front. Endocrinol. 2018, 9, 28. [Google Scholar] [CrossRef]
- Wang, X.P.; Huang, Z.; Li, Y.L.; Jin, K.Y.; Dong, D.J.; Wang, J.X.; Zhao, X.F. Krüppel-like factor 15 integrated autophagy and gluconeogenesis to maintain glucose homeostasis under 20-hydroxyecdysone regulation. PLoS Genet. 2022, 18, e1010229. [Google Scholar] [CrossRef]
- Kang, X.L.; Li, Y.X.; Dong, D.J.; Wang, J.X.; Zhao, X.F. 20-Hydroxyecdysone counteracts insulin to promote programmed cell death by modifying phosphoglycerate kinase 1. BMC Biol. 2023, 21, 119. [Google Scholar] [CrossRef]
- Tennessen, J.M. Ecdysone and 20-hydroxyecdysone are not required to activate glycolytic gene expression in Drosophila melanogaster embryos. MicroPubl. Biol. 2021, 2021. [Google Scholar] [CrossRef]
- Tettamanti, G.; Casartelli, M. Cell death during complete metamorphosis. Philos. Trans. R. Soc. B 2019, 374, 20190065. [Google Scholar] [CrossRef] [PubMed]
- Rost-Roszkowska, M.M. Degeneration of the midgut epithelium in Allacma fusca L. (Insecta, Collembola, Symphypleona): Apoptosis and necrosis. Zool. Sci. 2008, 25, 753–759. [Google Scholar] [CrossRef]
- Song, X.F.; Tian, H.; Zhang, P.; Zhang, Z.X. Expression of Cyt-c-Mediated Mitochondrial Apoptosis-Related Proteins in Rat Renal Proximal Tubules during Development. Nephron 2017, 135, 77–86. [Google Scholar] [CrossRef]
- Nicolson, S.; Denton, D.; Kumar, S. Ecdysone-mediated programmed cell death in Drosophila. Int. J. Dev. Biol. 2015, 59, 23–32. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Li, S. Sumoylation modulates 20-hydroxyecdysone signaling by maintaining USP protein levels in Drosophila. Insect Biochem. Mol. Biol. 2014, 54, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Cao, W.; Qiu, Y.; Deng, R.; Liu, J.; Zhang, L.; Dong, R.; Liu, F.; Li, S.; Zhao, H.; et al. The appearance of cytoplasmic cytochrome C precedes apoptosis during Drosophila salivary gland degradation. Insect Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Draizen, T.A.; Ewer, J.; Robinow, S. Genetic and hormonal regulation of the death of peptidergic neurons in the Drosophila central nervous system. J. Neurobiol. 1999, 38, 455–465. [Google Scholar] [CrossRef]
- Cao, C.; Liu, Y.; Lehmann, M. Fork head controls the timing and tissue selectivity of steroid-induced developmental cell death. J. Cell Biol. 2007, 176, 843–852. [Google Scholar] [CrossRef]
- Rusten, T.E.; Lindmo, K.; Juhász, G.; Sass, M.; Seglen, P.O.; Brech, A.; Stenmark, H. Programmed autophagy in the Drosophila fat body is induced by ecdysone through regulation of the PI3K pathway. Dev. Cell 2004, 7, 179–192. [Google Scholar] [CrossRef]
- Yin, V.P.; Thummel, C.S. Mechanisms of steroid-triggered programmed cell death in Drosophila. Semin. Cell Dev. Biol. 2005, 16, 237–243. [Google Scholar] [CrossRef]
- Dai, Y.; Li, K.; Wu, W.; Wu, K.; Yi, H.; Li, W.; Xiao, Y.; Zhong, Y.; Cao, Y.; Tian, L. Steroid hormone 20-hydroxyecdysone induces the transcription and complex assembly of V-ATPases to facilitate autophagy in Bombyx mori. Insect Biochem. Mol. Biol. 2020, 116, 103255. [Google Scholar] [CrossRef]
- Tian, L.; Liu, S.; Liu, H.; Li, S. 20-hydroxyecdysone upregulates apoptotic genes and induces apoptosis in the Bombyx fat body. Arch. Insect Biochem. Physiol. 2012, 79, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Ma, L.; Guo, E.; Deng, X.; Ma, S.; Xia, Q.; Cao, Y.; Li, S. 20-Hydroxyecdysone upregulates Atg genes to induce autophagy in the Bombyx fat body. Autophagy 2013, 9, 1172–1187. [Google Scholar] [CrossRef]
- Wang, X.; Ding, Y.; Lu, X.; Geng, D.; Li, S.; Raikhel, A.S.; Zou, Z. The ecdysone-induced protein 93 is a key factor regulating gonadotrophic cycles in the adult female mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2021, 118, e2021910118. [Google Scholar] [CrossRef] [PubMed]
- Romanelli, D.; Casartelli, M.; Cappellozza, S.; de Eguileor, M.; Tettamanti, G. Roles and regulation of autophagy and apoptosis in the remodelling of the lepidopteran midgut epithelium during metamorphosis. Sci. Rep. 2016, 6, 32939. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, J.; Xu, X.; Ma, W.; Tian, L.; Li, K. Functional identification of Bombyx mori Atg13 in autophagy. Arch. Insect Biochem. Physiol. 2020, 105, e21718. [Google Scholar] [CrossRef]
- Wu, W.; Luo, M.; Li, K.; Dai, Y.; Yi, H.; Zhong, Y.; Cao, Y.; Tettamanti, G.; Tian, L. Cholesterol derivatives induce dephosphorylation of the histone deacetylases Rpd3/HDAC1 to upregulate autophagy. Autophagy 2021, 17, 512–528. [Google Scholar] [CrossRef]
- Xu, X.; Cui, L.; Zhong, W.; Cai, Y. Autophagy-Associated lncRNAs: Promising Targets for Neurological Disease Diagnosis and Therapy. Neural Plast. 2020, 2020, 8881687. [Google Scholar] [CrossRef]
- Qiao, H.; Wang, J.; Wang, Y.; Yang, J.; Wei, B.; Li, M.; Wang, B.; Li, X.; Cao, Y.; Tian, L.; et al. Transcriptome analysis reveals potential function of long non-coding RNAs in 20-hydroxyecdysone regulated autophagy in Bombyx mori. BMC Genom. 2021, 22, 374. [Google Scholar] [CrossRef]
- Di, Y.Q.; Han, X.L.; Kang, X.L.; Wang, D.; Chen, C.H.; Wang, J.X.; Zhao, X.F. Autophagy triggers CTSD (cathepsin D) maturation and localization inside cells to promote apoptosis. Autophagy 2021, 17, 1170–1192. [Google Scholar] [CrossRef]
- Pan, J.; Di, Y.Q.; Li, Y.B.; Chen, C.H.; Wang, J.X.; Zhao, X.F. Insulin and 20-hydroxyecdysone oppose each other in the regulation of phosphoinositide-dependent kinase-1 expression during insect pupation. J. Biol. Chem. 2018, 293, 18613–18623. [Google Scholar] [CrossRef]
- Li, Y.B.; Pei, X.Y.; Wang, D.; Chen, C.H.; Cai, M.J.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone upregulates calcium release-activated calcium channel modulator 1 expression to induce apoptosis in the midgut of Helicoverpa armigera. Cell Calcium 2017, 68, 24–33. [Google Scholar] [CrossRef]
- Chen, C.H.; Di, Y.Q.; Shen, Q.Y.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone induces phosphorylation and aggregation of stromal interacting molecule 1 for store-operated calcium entry. J. Biol. Chem. 2019, 294, 14922–14936. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Pan, J.; Di, Y.Q.; Liu, W.; Hou, L.; Wang, J.X.; Zhao, X.F. Protein kinase C delta phosphorylates ecdysone receptor B1 to promote gene expression and apoptosis under 20-hydroxyecdysone regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E7121–E7130. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Liu, W.; Zhao, W.L.; Wang, J.X.; Zhao, X.F. Upregulation of the expression of prodeath serine/threonine protein kinase for programmed cell death by steroid hormone 20-hydroxyecdysone. Apoptosis 2013, 18, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Mané-Padrós, D.; Cruz, J.; Vilaplana, L.; Nieva, C.; Ureña, E.; Bellés, X.; Martín, D. The hormonal pathway controlling cell death during metamorphosis in a hemimetabolous insect. Dev. Biol. 2010, 346, 150–160. [Google Scholar] [CrossRef]
- Yu, J.; Song, H.; Wang, Y.; Liu, Z.; Wang, H.; Xu, B. 20-hydroxyecdysone Upregulates Ecdysone Receptor (ECR) Gene to Promote Pupation in the Honeybee, Apis mellifera Ligustica. Integr. Comp. Biol. 2023, 63, 288–303. [Google Scholar] [CrossRef]
- Li, Z.; Lyu, Z.; Ye, Q.; Cheng, J.; Wang, C.; Lin, T. Cloning, Expression Analysis, 20-Hydroxyecdysone Induction, and RNA Interference Study of Autophagy-Related Gene 8 from Heortia vitessoides Moore. Insects 2020, 11, 245. [Google Scholar] [CrossRef]
- Poyraz, T.E.; Goncu, E.; Koc, K. Apoptotic and autophagic characteristics of perivisceral fat body remodeling of the greater wax moth Galleria mellonella and effects of juvenile hormone analog, fenoxycarb, on these processes. Arch. Insect Biochem. Physiol. 2021, 107, e21780. [Google Scholar]
- Ekoka, E.; Maharaj, S.; Nardini, L.; Dahan-Moss, Y.; Koekemoer, L.L. 20-Hydroxyecdysone (20E) signaling as a promising target for the chemical control of malaria vectors. Parasite Vector 2021, 14, 86. [Google Scholar] [CrossRef]
Figure 1.
EcR-USP swiftly and potently induces the expression of primary-response genes upon 20E stimulation.
Figure 1.
EcR-USP swiftly and potently induces the expression of primary-response genes upon 20E stimulation.

Figure 2.
20E regulated autophagy and apoptosis in insects.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, L.; Li, H.; Zheng, Q.; Hu, J.; Wu, W. Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone. Insects 2023, 14, 871. https://doi.org/10.3390/insects14110871
AMA Style
Lin L, Li H, Zheng Q, Hu J, Wu W. Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone. Insects. 2023; 14(11):871. https://doi.org/10.3390/insects14110871
Chicago/Turabian StyleLin, Luobin, Huaqin Li, Qinzhou Zheng, Jiaxuan Hu, and Wenmei Wu. 2023. "Research Progress on the Regulation of Autophagy and Apoptosis in Insects by Sterol Hormone 20-Hydroxyecdysone" Insects 14, no. 11: 871. https://doi.org/10.3390/insects14110871
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.