

Complex Effects of a Land-Use Gradient on Pollinators and Natural Enemies: Natural Habitats Mitigate the Effects of Aphid Infestation on Pollination Services

Abstract
:Simple Summary
Abstract
1. Introduction
- (1)
- What are the combined and interactive effects of pollinators, pests, and their natural enemies on fruit and seed set in an agro-ecosystem?
- (2)
- How do these interactions vary with local and landscape habitat characteristics?
2. Methods
2.1. Study Area
2.2. Experimental Design
2.3. Phytometer Species and Aphid Infestation
2.4. Data Collection
- (a)
- Landscape and local habitat characteristics
- (b)
- Pollination services
- (c)
- Biological control services
- (1)
- Aphid predation rate—calculated per phytometer plant by the ratio where and are the numbers of aphids in the caged and uncaged treatments respectively, on day 5 in the field. This ratio serves as an index ranging from 0 to 1, with higher values corresponding to higher predation rates. On the fifth day in the field, all aphids and large nymphs (stages 3 and 4) were systematically and thoroughly counted on plant leaves and stems in both treatments. Subsequently, each plant was enveloped with a dense organza bag and transported to the laboratory for aphid parasitism determination.
- (2)
- Aphid parasitism rate—at the end of the fifth day in the field, each infested phytometer was enclosed within an organza bag and transferred to the laboratory for the monitoring of aphid mummy development (i.e., the formation of parasitoid pupae within its host aphid) using the following method: two aphid-infested leaves per plant (50–80 adult aphids per leaf) were detached, and the petiole was inserted in water-filled tubes and placed in a monitoring cage (Figure S2). Adequate fresh leaves and water were supplied as needed for maintaining the aphids. On days 7 and 10, the number of formed mummies was recorded. The same procedure was carried out for control plants in the caged infested treatment. All emerging parasitoids were identified to the genus or species level.
2.5. Statistical Analyses
3. Results
3.1. Effects of Land Use on Pollination Services
3.2. Effects of Land Use on Aphid Parasitism and Predation Rates
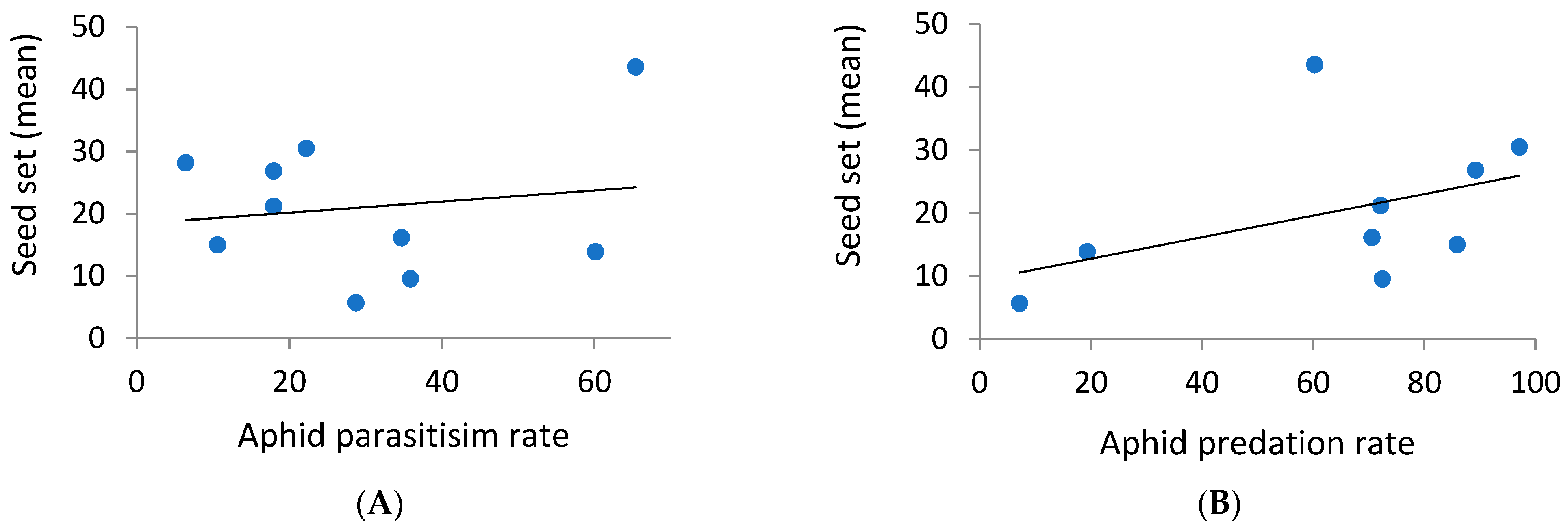
3.3. Combined Effects of Natural Enemies and Pollinators on the Seed Set of Aphid-Infested Phytometers
4. Discussion
4.1. Pollinator and Natural Enemy Responses to Landscape and Habitat Characteristics
4.2. Interactions among Pollinators, Herbivores, and the Pollination and Pest Control Services They Deliver across Landscape Gradients
4.3. Combined Effects of Natural Enemies and Pollinators on Seed Set by Aphid-Infested Phytometers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Power, A.G. Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2959–2971. [Google Scholar] [CrossRef]
- Dainese, M.; Martin, E.A.; Aizen, M.A.; Albrecht, M.; Bartomeus, I.; Bommarco, R.; Carvalheiro, L.G.; Chaplin-Kramer, R.; Gagic, V.; Garibaldi, L.A.; et al. A global synthesis reveals biodiversity-mediated benefits for crop production. Sci. Adv. 2019, 5, eaax0121. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. BioScience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Jeanneret, P.; Aviron, S.; Alignier, A.; Lavigne, C.; Helfenstein, J.; Herzog, F.; Kay, S.; Petit, S. Agroecology landscapes. Landsc. Ecol. 2021, 36, 2235–2257. [Google Scholar] [CrossRef]
- Lundin, O.; Rundlöf, M.; Jonsson, M.; Bommarco, R.; Williams, N.M. Integrated pest and pollinator management—Expanding the concept. Front. Ecol. Environ. 2021, 19, 283–291. [Google Scholar] [CrossRef]
- Jeavons, E.; Le Lann, C.; van Baaren, J. Interactions between natural enemies and pollinators: Combining ecological theory with agroecological management. Entomol. Gen. 2023, 43, 243–259. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Andersson, G.K.S.; Requier, F.; Fijen, T.P.M.; Hipólito, J.; Kleijn, D.; Pérez-Méndez, N.; Rollin, O. Complementarity and synergisms among ecosystem services supporting crop yield. Glob. Food Secur. 2018, 17, 38–47. [Google Scholar] [CrossRef]
- Lundin, O.; Smith, H.G.; Rundlof, M.; Bommarco, R. When ecosystem services interact: Crop pollination benefits depend on the level of pest control. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122243. [Google Scholar] [CrossRef]
- Sutter, L.; Albrecht, M. Synergistic interactions of ecosystem services: Florivorous pest control boosts crop yield increase through insect pollination. Proceedings. Biol. Sci. R. Soc. 2016, 283, 20152529. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Schmid, B.; Hautier, Y.; Mueller, C.B. Diverse pollinator communities enhance plant reproductive success. Proc. R. Soc. B Biol. Sci. 2012, 279, 4845–4852. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, S.A.M.; Herbertsson, L.; Rundlof, M.; Bommarco, R.; Smith, H.G. Experimental evidence that honeybees depress wild insect densities in a flowering crop. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161641. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.J.; Biesmeijer, J.C.; Varma, V.; Wäckers, F.L. Realising multiple ecosystem services based on the response of three beneficial insect groups to floral traits and trait diversity. Basic Appl. Ecol. 2012, 13, 363–370. [Google Scholar] [CrossRef]
- Jeavons, E.; van Baaren, J.; Le Lann, C. Resource partitioning among a pollinator guild: A case study of monospecific flower crops under high honeybee pressure. Acta Oecologica 2020, 104, 103527. [Google Scholar] [CrossRef]
- Greco, C.F.; Kevan, P.G. Patch choice in the anthophilous ambush predator Phymata americana: Improvement by switching hunting sites as part of the initial choice. Can. J. Zool. Rev. Can. Zool. 1995, 73, 1912–1917. [Google Scholar] [CrossRef]
- Sinu, P.A.; Sibisha, V.C.; Reshmi, M.V.N.; Reshmi, K.S.; Jasna, T.V.; Aswathi, K.; Megha, P.P. Invasive ant (Anoplolepis gracilipes) disrupts pollination in pumpkin. Biol. Invasions 2017, 19, 2599–2607. [Google Scholar] [CrossRef]
- Gagic, V.; Marcora, A.; Howie, L. Additive and interactive effects of pollination and biological pest control on crop yield. J. Appl. Ecol. 2019, 56, 2528–2535. [Google Scholar] [CrossRef]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of pollinators and natural enemies: A meta-analysis of landscape and local effects on abundance and richness in crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does landscape composition affect pest abundance and their control by natural enemies? A review. Agric. Ecosyst. Environ. 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Holland, J.M.; Douma, J.C.; Crowley, L.; James, L.; Kor, L.; Stevenson, D.R.W.; Smith, B.M. Semi-natural habitats support biological control, pollination and soil conservation in Europe. A review. Agron. Sustain. Dev. 2017, 37, 31. [Google Scholar] [CrossRef]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martinez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [PubMed]
- Bartual, A.M.; Sutter, L.; Bocci, G.; Moonen, A.C.; Cresswell, J.; Entling, M.; Giffard, B.; Jacot, K.; Jeanneret, P.; Holland, J.; et al. The potential of different semi-natural habitats to sustain pollinators and natural enemies in European agricultural landscapes. Agric. Ecosyst. Environ. 2019, 279, 43–52. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Martin, E.A.; Dainese, M.; Clough, Y.; Baldi, A.; Bommarco, R.; Gagic, V.; Garratt, M.P.D.; Holzschuh, A.; Kleijn, D.; Kovacs-Hostyanszki, A.; et al. The interplay of landscape composition and configuration: New pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol. Lett. 2019, 22, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Egan, P.A.; Dicks, L.V.; Hokkanen, H.M.T.; Stenberg, J.A. Delivering Integrated Pest and Pollinator Management (IPPM). Trends Plant Sci. 2020, 25, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Pisanty, G.; Mandelik, Y. Profiling crop pollinators: Life history traits predict habitat use and crop visitation by Mediterranean wild bees. Ecol. Appl. 2015, 25, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Weizel, Y.; Polak, G.; Cohen, Y. Ecology of the Vegetation in Israel; Tel Aviv University: Tel Aviv-Yafo, Israel, 1978. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2000. [Google Scholar]
- Roth, T.; Coll, M.; Mandelik, Y. The role of uncultivated habitats in supporting wild bee communities in Mediterranean agricultural landscapes. Diversity 2023, 15, 294. [Google Scholar] [CrossRef]
- Johnson, J.B.; Omland, K.S. Model selection in ecology and evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. In R Package Version 0.4.6; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bartoń, K. MuMIn:Multi-Model Inference. In R Package Version 1.47.5; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Lüdecke, D. sjPlot:Data Visualization for Statistics in Social. In R Package Version 2.8.14; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Macfadyen, S.; Muller, W. Edges in agricultural landscapes: Species interactions and movement of natural enemies. PLoS ONE 2013, 8, e59659. [Google Scholar] [CrossRef] [PubMed]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M.S. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Munzenberg, U.; Burger, C.; Thies, C.; Tscharntke, T. Scale-dependent effects of landscape context on three pollinator guilds. Ecology 2002, 83, 1421–1432. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Jervis, M.A. Does floral nectar improve biological control by parasitoids. In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Cambridge University Press: Cambridge, UK, 2005; pp. 267–304. [Google Scholar]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batary, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef]
- Duflot, R.; Aviron, S.; Ernoult, A.; Fahrig, L.; Burel, F. Reconsidering the role of ‘semi-natural habitat’ in agricultural landscape biodiversity: A case study. Ecol. Res. 2015, 30, 75–83. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Banos-Picon, L.; Tormos, J.; Asis, J.D. Natural enemies and pollinators in traditional cherry orchards: Functionally important taxa respond differently to farming system. Agric. Ecosyst. Environ. 2020, 295, 106920. [Google Scholar] [CrossRef]
- Nottebrock, H.; Schmid, B.; Mayer, K.; Devaux, C.; Esler, K.J.; Bohning-Gaese, K.; Schleuning, M.; Pagel, J.; Schurr, F.M. Sugar landscapes and pollinator-mediated interactions in plant communities. Ecography 2017, 40, 1129–1138. [Google Scholar] [CrossRef]
- Heimpel, G.E. Linking parasitoid nectar feeding and dispersal in conservation biological control. Biol. Control 2019, 132, 36–41. [Google Scholar] [CrossRef]
- Holzschuh, A.; Dainese, M.; Gonzalez-Varo, J.P.; Mudri-Stojnic, S.; Riedinger, V.; Rundlof, M.; Scheper, J.; Wickens, J.B.; Wickens, V.J.; Bommarco, R.; et al. Mass-flowering crops dilute pollinator abundance in agricultural landscapes across Europe. Ecol. Lett. 2016, 19, 1228–1236. [Google Scholar] [CrossRef]
- Evans, T.M.; Cavers, S.; Ennos, R.; Vanbergen, A.J.; Heard, M.S. Florally rich habitats reduce insect pollination and the reproductive success of isolated plants. Ecol. Evol. 2017, 7, 6507–6518. [Google Scholar] [CrossRef] [PubMed]
- Grab, H.; Blitzer, E.J.; Danforth, B.; Loeb, G.; Poveda, K. Temporally dependent pollinator competition and facilitation with mass flowering crops affects yield in co-blooming crops. Sci. Rep. 2017, 7, 45296. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.H.; Wang, J.M.; Pan, D.D.; An, J.D. Landscape-modified concentration effect and waylaying effect of bees and their consequences on pollination of mass-flowering plants in agricultural ecosystems. Agric. Ecosyst. Environ. 2019, 280, 24–34. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Rusman, Q.; Poelman, E.H.; Nowrin, F.; Polder, G.; Lucas-Barbosa, D. Floral plasticity: Herbivore-species-specific-induced changes in flower traits with contrasting effects on pollinator visitation. Plant Cell Environ. 2019, 42, 1882–1896. [Google Scholar] [CrossRef]
- Strauss, S.Y.; Conner, J.K.; Rush, S.L. Foliar herbivory affects floral characters and plant attractiveness to pollinators: Implications for male and female plant fitness. Am. Nat. 1996, 147, 1098–1107. [Google Scholar] [CrossRef]
- Lehtila, K.; Strauss, S.Y. Leaf damage by herbivores affects attractiveness to pollinators in wild radish, Raphanus raphanistrum. Oecologia 1997, 111, 396–403. [Google Scholar] [CrossRef]
- Krupnick, G.A.; Weis, A.E.; Campbell, D.R. The consequences of floral herbivory for pollinator service to Isomeris arborea. Ecology 1999, 80, 125–134. [Google Scholar] [CrossRef]
- Mothershead, K.; Marquis, R.J. Fitness impacts of herbivory through indirect effects on plant-pollinator interactions in Oenothera macrocarpa. Ecology 2000, 81, 30–40. [Google Scholar] [CrossRef]
- Barber, N.A.; Adler, L.S.; Theis, N.; Hazzard, R.V.; Kiers, E.T. Herbivory reduces plant interactions with above- and belowground antagonists and mutualists. Ecology 2012, 93, 1560–1570. [Google Scholar] [CrossRef]
- Moreira, X.; Castagneyrol, B.; Abdala-Roberts, L.; Traveset, A. A meta-analysis of herbivore effects on plant attractiveness to pollinators. Ecology 2019, 100, e02707. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Heimpel, G. Nectar availability and parasitoid sugar feeding. In Proceedings of the 1st International Symposium on Biological Control of Arthropods, Honolulu, HI, USA, 14–18 January 2002; USDA Forest Service: Morgantown, WV, USA, 2003. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable (& Model Type) | Fixed Effects Predictors | Random Effects |
|---|---|---|
| Number of visits in flowers - Infested & un-infested phytometers: Overall pollinators Honeybees Wild bees Flies (Glmer, Poisson/ glmer, Negative binomial) | Proportion of natural/semi-natural area within: 250–2000 m radius (1) Aphid treatment Aphid treatment * % natural/semi-natural Local flower abundance Local flower species richness Number of flowers [offset variable] | Date/plot (2) Sub-region |
| Number of visits in flowers - Infested phytometers only: Overall pollinators Honeybees Wild bees Flies (Glmer, Poisson/ glmer, negative binomial) | Proportion of natural/semi-natural area within: 250–2000 m radius (1) Aphid treatment Aphid treatment * % natural/semi-natural Local flower abundance Local flower species richness Aphid predation rate Aphid parasitism rate Number of flowers [offset variable] | Date Sub-region |
| Aphid predation rate (lmer) Aphid parasitism rate (glmer, binomial) | Proportion of natural/semi-natural area within: 250–2000 m radius (1) Local flower abundance Local flower species richness Pollinator visit frequency | Date Sub-region |
| Seed set in infested phytometers (Glmer, Poisson/ glmer, Negative binomial) | Pollinator visit frequency * Aphid predation rate Pollinator visit frequency * Aphid parasitism rate Aphid predation rate * Aphid parasitism rate Number of pods [offset variable] | Date Sub-region |
| (1) A separated model for each radius | ||
| (2) Date nested in plot * for interaction |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shapira, T.; Roth, T.; Bar, A.; Coll, M.; Mandelik, Y. Complex Effects of a Land-Use Gradient on Pollinators and Natural Enemies: Natural Habitats Mitigate the Effects of Aphid Infestation on Pollination Services. Insects 2023, 14, 872. https://doi.org/10.3390/insects14110872
Shapira T, Roth T, Bar A, Coll M, Mandelik Y. Complex Effects of a Land-Use Gradient on Pollinators and Natural Enemies: Natural Habitats Mitigate the Effects of Aphid Infestation on Pollination Services. Insects. 2023; 14(11):872. https://doi.org/10.3390/insects14110872
Chicago/Turabian StyleShapira, Tal, Tohar Roth, Adi Bar, Moshe Coll, and Yael Mandelik. 2023. "Complex Effects of a Land-Use Gradient on Pollinators and Natural Enemies: Natural Habitats Mitigate the Effects of Aphid Infestation on Pollination Services" Insects 14, no. 11: 872. https://doi.org/10.3390/insects14110872