
Estimating the Potential of Insects from Warmer Regions to Overwinter in Colder Regions under a Warming Winter Scenario Using Simulation Experiments: A Case Study in Sesamia nonagrioides

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Treatments
2.3. Analysis of Survival
2.4. Statistical Analysis
3. Results
3.1. Survival in the Laboratory Scenarios and in the Field
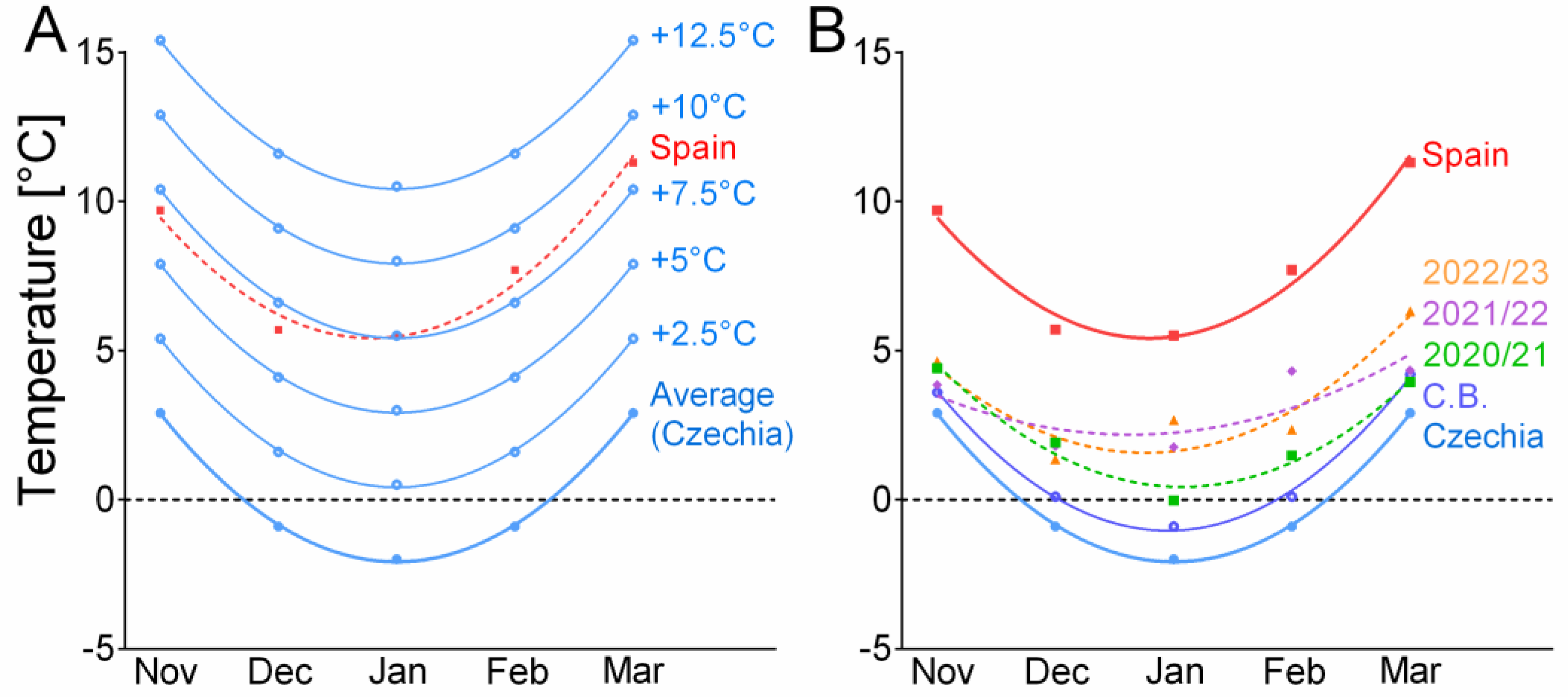
3.2. Temperature Conditions in the Field
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harrison, J.F.; Woods, H.A.; Roberts, S.P. Ecological and Environmental Physiology of Insects; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Stevens, P.W.; Lamont, M.M.; Brusca, R.C.; Hart, K.M.; Waddle, J.H.; Langtimm, C.A.; Williams, C.M.; Keim, B.D.; Terando, A.J.; et al. Tropicalization of temperate ecosystems in North America: The northward range expansion of tropical organisms in response to warming winter temperatures. Glob. Chang. Biol. 2021, 27, 3009–3034. [Google Scholar] [CrossRef] [PubMed]
- Outhwaite, C.L.; McCann, P.; Newbold, T. Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 2022, 605, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Blumenthal, D.M.; Runion, G.B.; Hunt, R.E., Jr.; Diaz-Soltero, H. Invasive species and climate change: An agronomic perspective. Clim. Chang. 2011, 105, 13–42. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Sunday, J.; Bates, A.; Dulvy, N. Thermal tolerance and the global redistribution of animals. Nat. Clim. Chang. 2012, 2, 686–690. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Climate change and biological invasions: Evidence, expectations, and response options. Biol. Rev. 2017, 92, 1297–1313. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Screen, J.A.; Furtado, J.C.; Barlow, M.; Whittleston, D.; Coumou, D.; Francis, J.; Dethloff, K.; Entekhabi, D.; Overland, J.; et al. Recent Arctic amplification and extreme mid-latitude weather. Nat. Geosci. 2014, 7, 627–637. [Google Scholar] [CrossRef]
- Hill, M.P.; Thomson, L.J. Species distribution modelling in predicting response to climate change. In Climate Change and Insect Pests; Björkman, C., Niemelä, P., Eds.; CABI: Wallingford, UK, 2015; pp. 16–37. [Google Scholar]
- Moyal, P.; Tokro, P.; Bayram, A.; Savopoulou-Soultani, M.; Conti, E.; Eizaguirre, M.; Le Rue, B.; Avand-Faghih, A.; Frerot, B.; Andreadis, S. Origin and taxonomic status of the Palearctic population of the stem borer Sesamia nonagrioides (Lefèbvre) (Lepidoptera: Noctuidae). Biol. J. Linn 2011, 103, 904–922. [Google Scholar] [CrossRef]
- Eizaguirre, M.; Fantinou, A.A. Abundance of Sesamia nonagrioides (Lef.) (Lepidoptera:Noctuidae) on the Edges of the Mediterranean Basin. Psyche A J. Entomol. 2012, 2012, 854045. [Google Scholar] [CrossRef]
- Eizaguirre, M.; Albajes, R. Diapause induction in the Stem Corn Borer, Sesamia nonagrioides (Lepidoptera: Noctuidae). Entomol. Gener. 1992, 17, 277–283. [Google Scholar] [CrossRef]
- Gillyboeuf, N.; Anglade, P.; Lavenseau, L.; Peypelut, L. Cold hardiness and overwintering strategy of the pink maize stalk borer, Sesamia nonagrioides Lef (lepidoptera, noctuidae). Oecologia 1994, 99, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, S.S.; Vryzas, Z.; Papadopoulou-Mourkidou, E.; Savopoulou-Soultani, M. Cold tolerance of field-collected and laboratory reared larvae of Sesamia nonagrioides (Lepidoptera: Noctuidae). CryoLetters 2011, 32, 297–307. [Google Scholar] [PubMed]
- Fantinou, A.A.; Karandinos, M.G.; Tsitsipis, A.A. Diapause Induction in the Sesamia nonargioides (Lepidoptera: Noctuidae) Effect of Photoperiod and Temperature. Environ. Entomol. 1995, 24, 1458–1466. [Google Scholar] [CrossRef]
- Štěpánek, P.; Trnka, M.; Meitner, J.; Dubrovský, M.; Zahradníček, P.; Lhotka, O.; Skalák, P.; Kyselý, J.; Farda, A.; Semerádová, D. Očekávané Klimatické Podmínky v České Republice Část I. Změna Základních Parametrů (Expected Climatic Conditions in the Czech Republic Part I. Change in Basic Parameters; in Czech); CzechGlobe: Brno, Czech Republic, 2019. [Google Scholar]
- Maiorano, A.; Donatelli, M.; Fumagalli, D. Potential distribution and phenological development of the Mediterranean Corn Borer (Sesamia nonagrioides) under warming climate in Europe. In Proceedings of the 6th International Congress on Environmental Modelling and Software, Leipzig, Germany, 1–5 July 2012; 324. [Google Scholar]
- Williams, C.M.; Henry, H.A.L.; Sinclair, B.J. Cold truths: How winter drives responses of terrestrial organisms to climate change. Biol. Rev. 2015, 90, 214–235. [Google Scholar] [CrossRef] [PubMed]
- Studd, E.K.; Bates, A.E.; Bramburger, A.J.; Fernandes, T.; Hayden, B.; Henry, H.A.; Humphries, M.M.; Martin, R.; McMeans, B.C.; Moise, E.R.; et al. Nine maxims for the ecology of cold-climate winters. BioScience 2021, 71, 820–830. [Google Scholar] [CrossRef]
- Forrest, J.R. Complex responses of insect phenology to climate change. Curr. Opin. Insect. Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozsypal, J. Estimating the Potential of Insects from Warmer Regions to Overwinter in Colder Regions under a Warming Winter Scenario Using Simulation Experiments: A Case Study in Sesamia nonagrioides. Insects 2023, 14, 957. https://doi.org/10.3390/insects14120957
Rozsypal J. Estimating the Potential of Insects from Warmer Regions to Overwinter in Colder Regions under a Warming Winter Scenario Using Simulation Experiments: A Case Study in Sesamia nonagrioides. Insects. 2023; 14(12):957. https://doi.org/10.3390/insects14120957
Chicago/Turabian StyleRozsypal, Jan. 2023. "Estimating the Potential of Insects from Warmer Regions to Overwinter in Colder Regions under a Warming Winter Scenario Using Simulation Experiments: A Case Study in Sesamia nonagrioides" Insects 14, no. 12: 957. https://doi.org/10.3390/insects14120957