
High and Low Temperatures Differentially Affect Survival, Reproduction, and Gene Transcription in Male and Female Moths of Spodoptera frugiperda
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
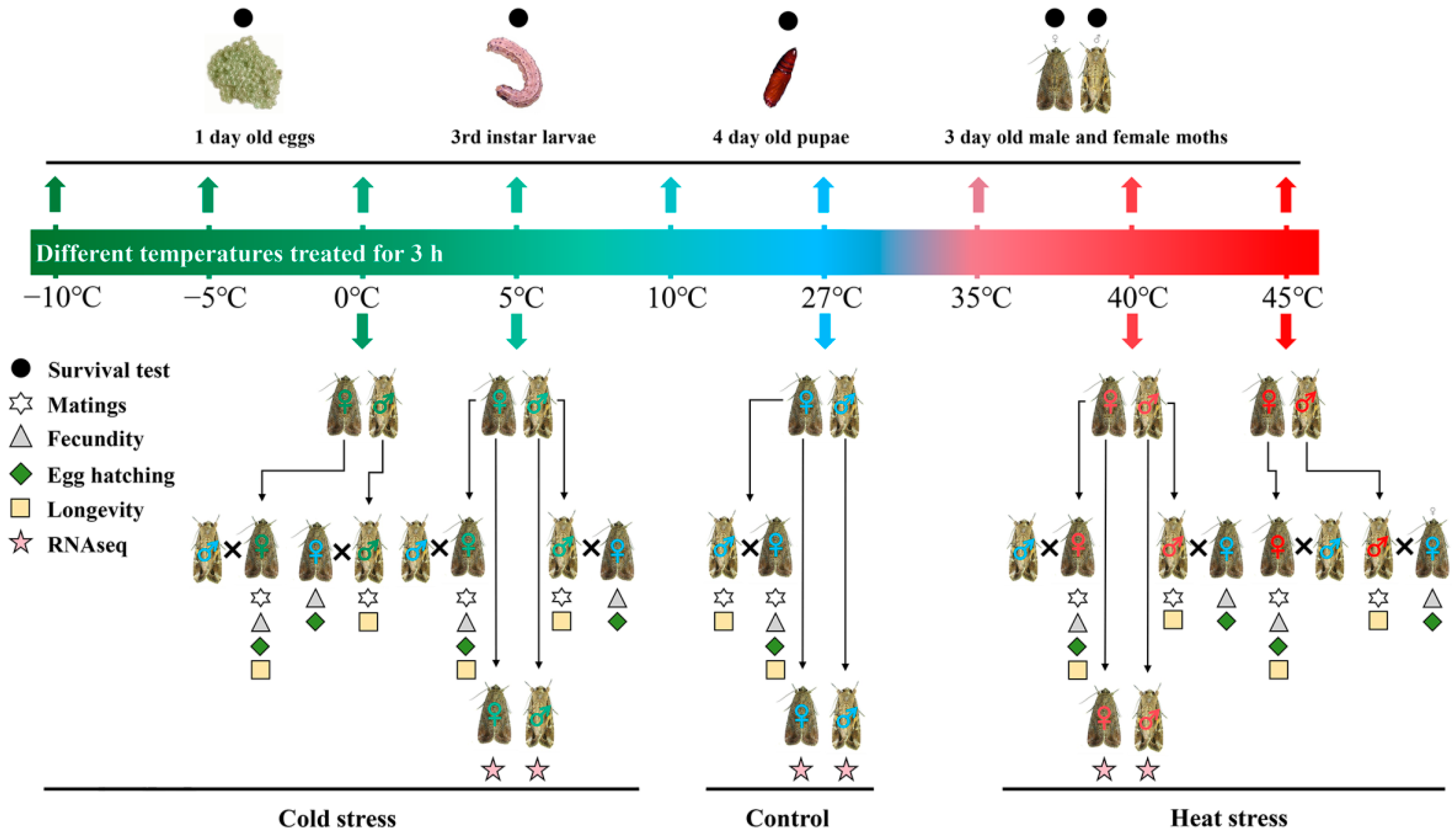
2. Materials and Methods
2.1. Insects
2.2. Effect of Temperature Stress on Survival and Reproduction
2.3. Effect of Temperature Stress on Gene Transcription
2.3.1. Treatments and Sampling
2.3.2. Library Preparation and Sequencing
2.3.3. Quality Control and Assembly
2.3.4. Differential Expression Analysis and Enrichment Analysis of Differentially Expressed Genes
2.3.5. Validation of RNA-Seq Sequencing Data
3. Results
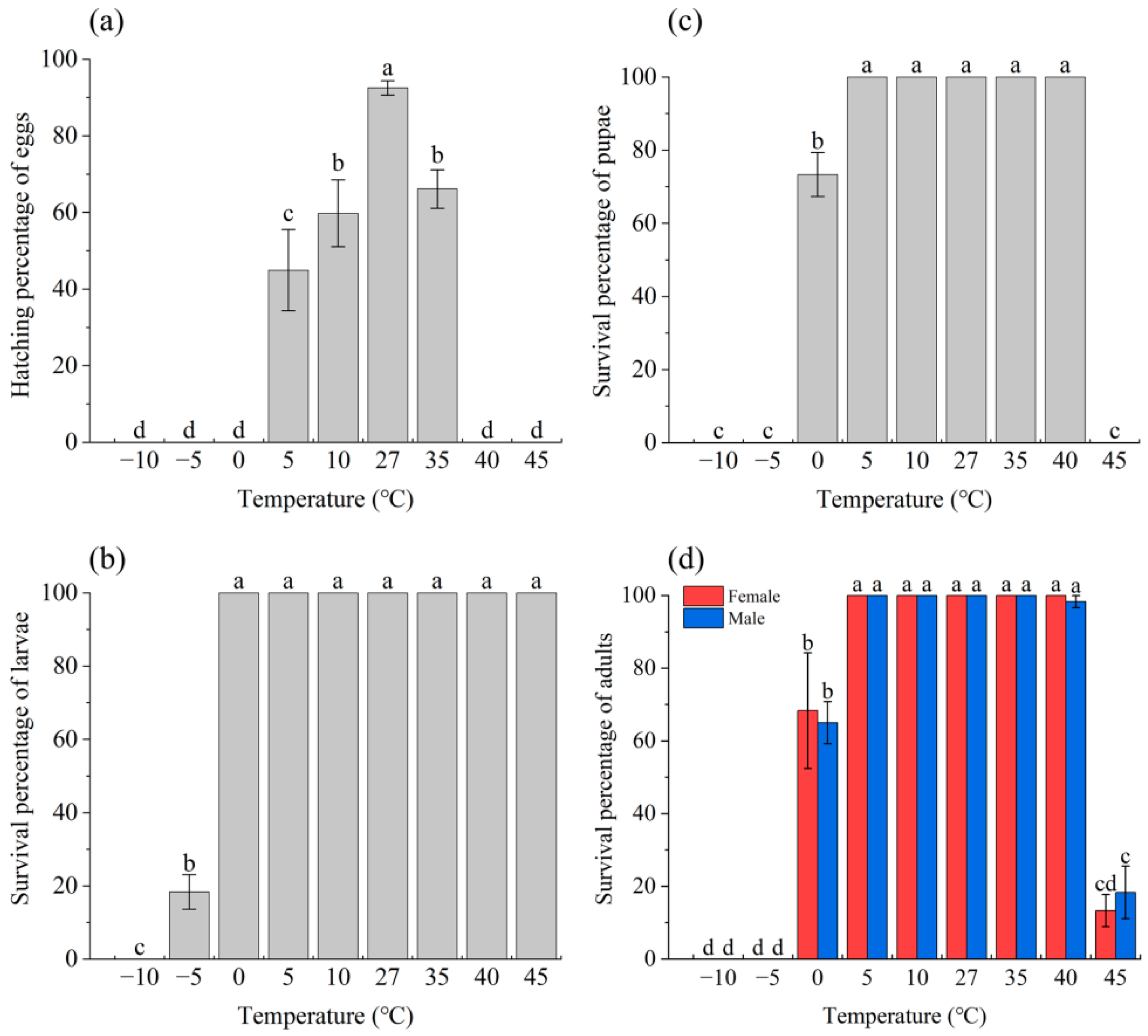
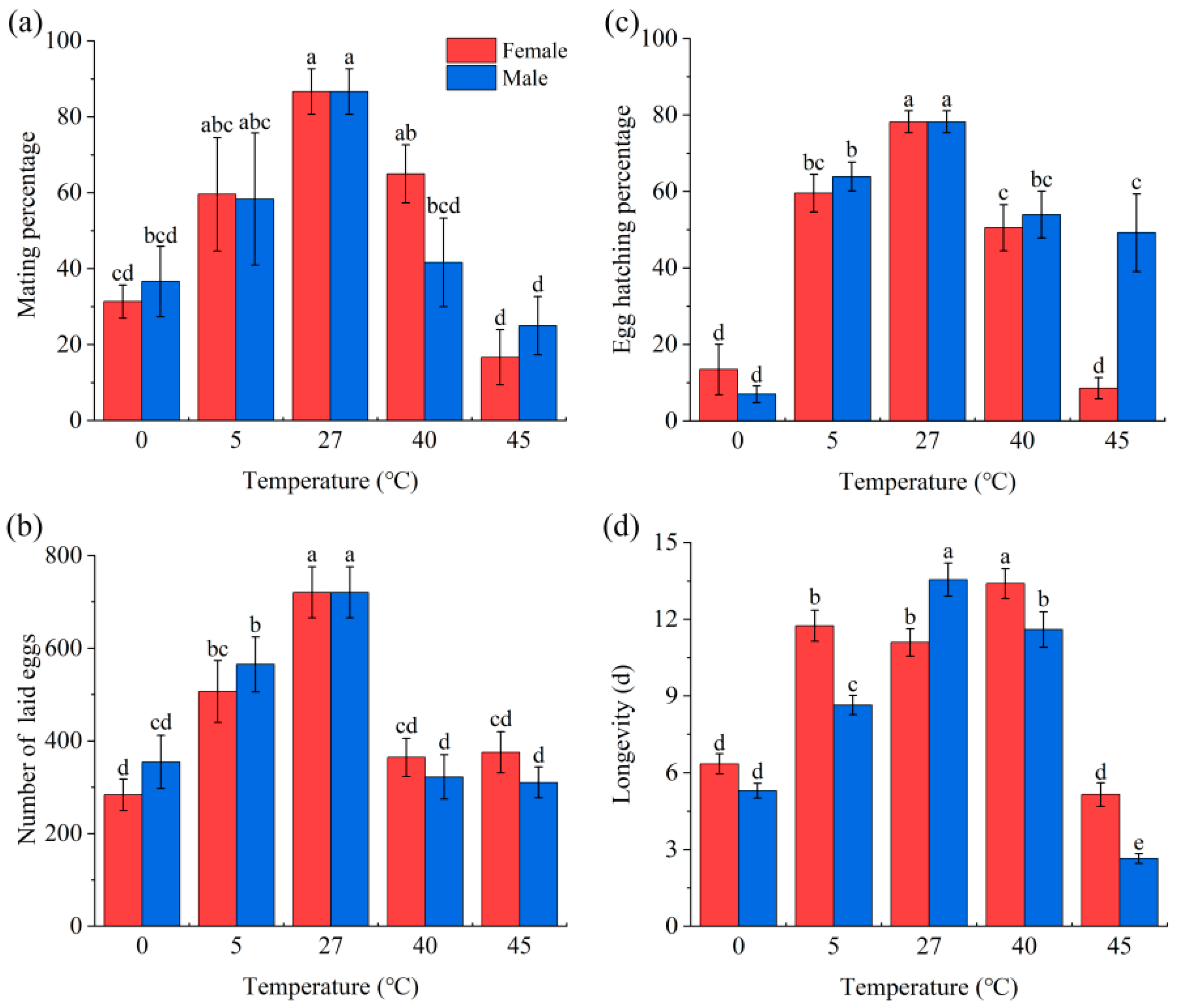
3.1. Effect of Temperature Stress on Survival and Reproduction
3.2. RNA Sequencing and Assembly
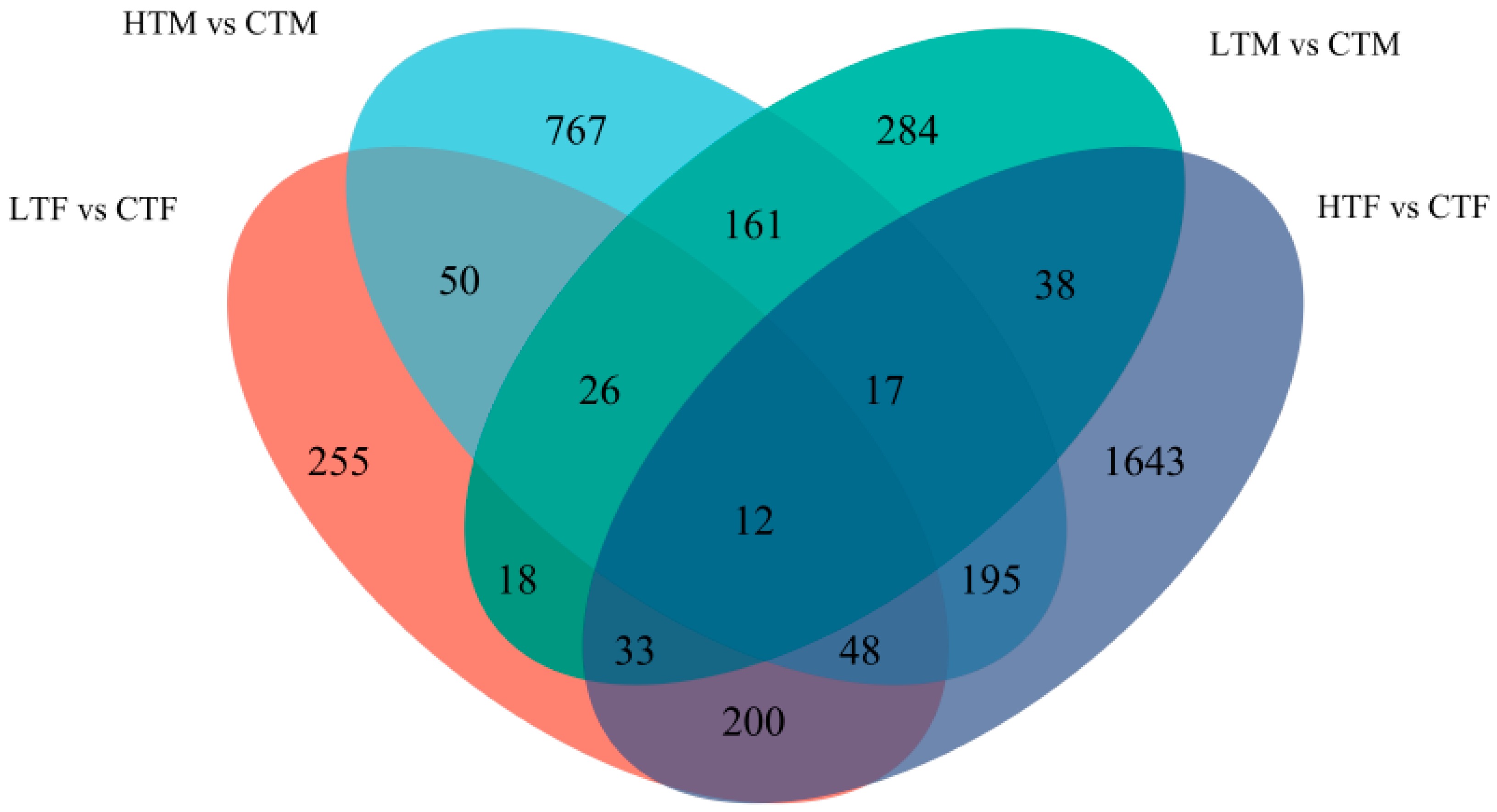
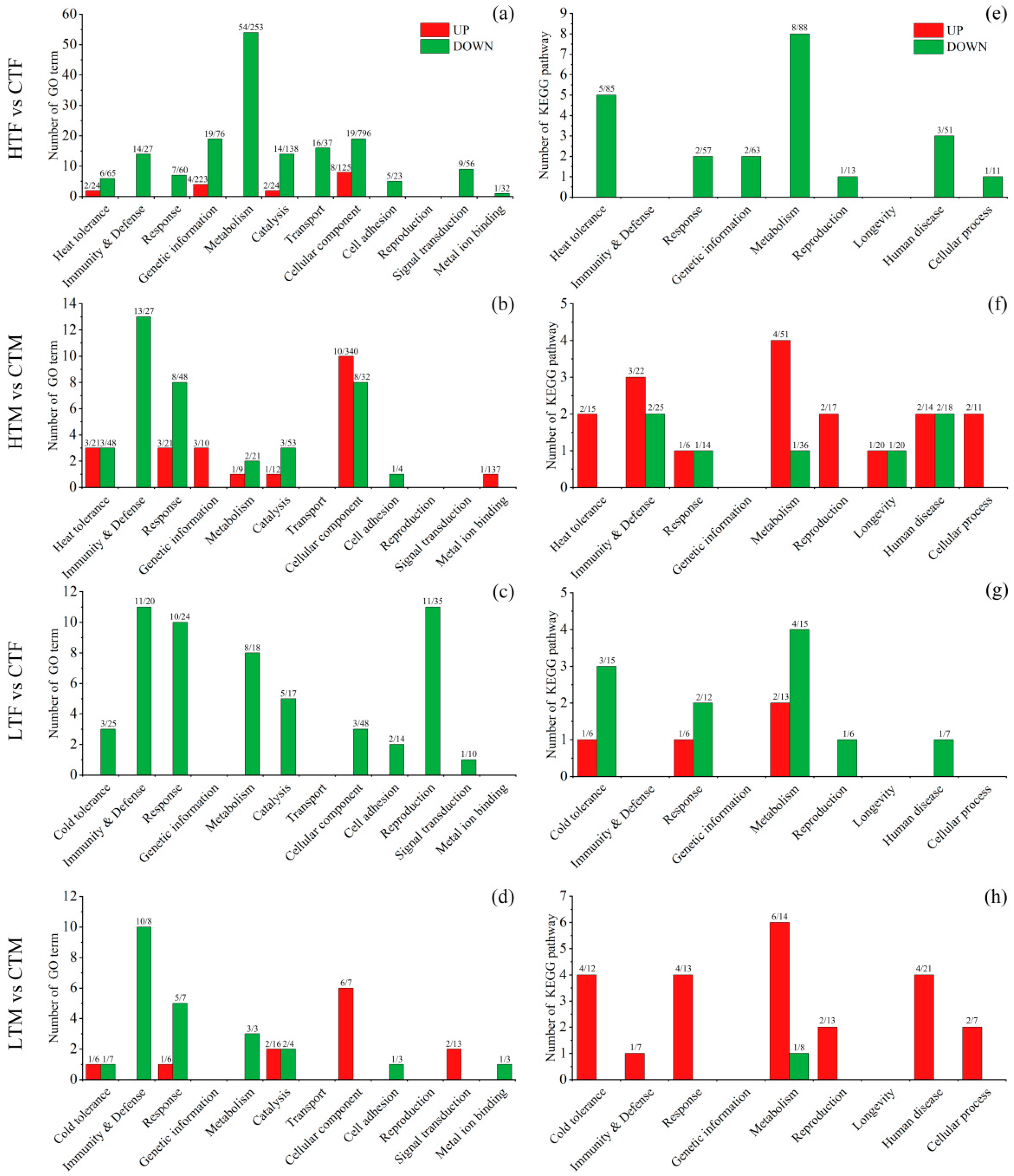
3.3. Overview of Transcriptional Changes and Enrichment Analysis
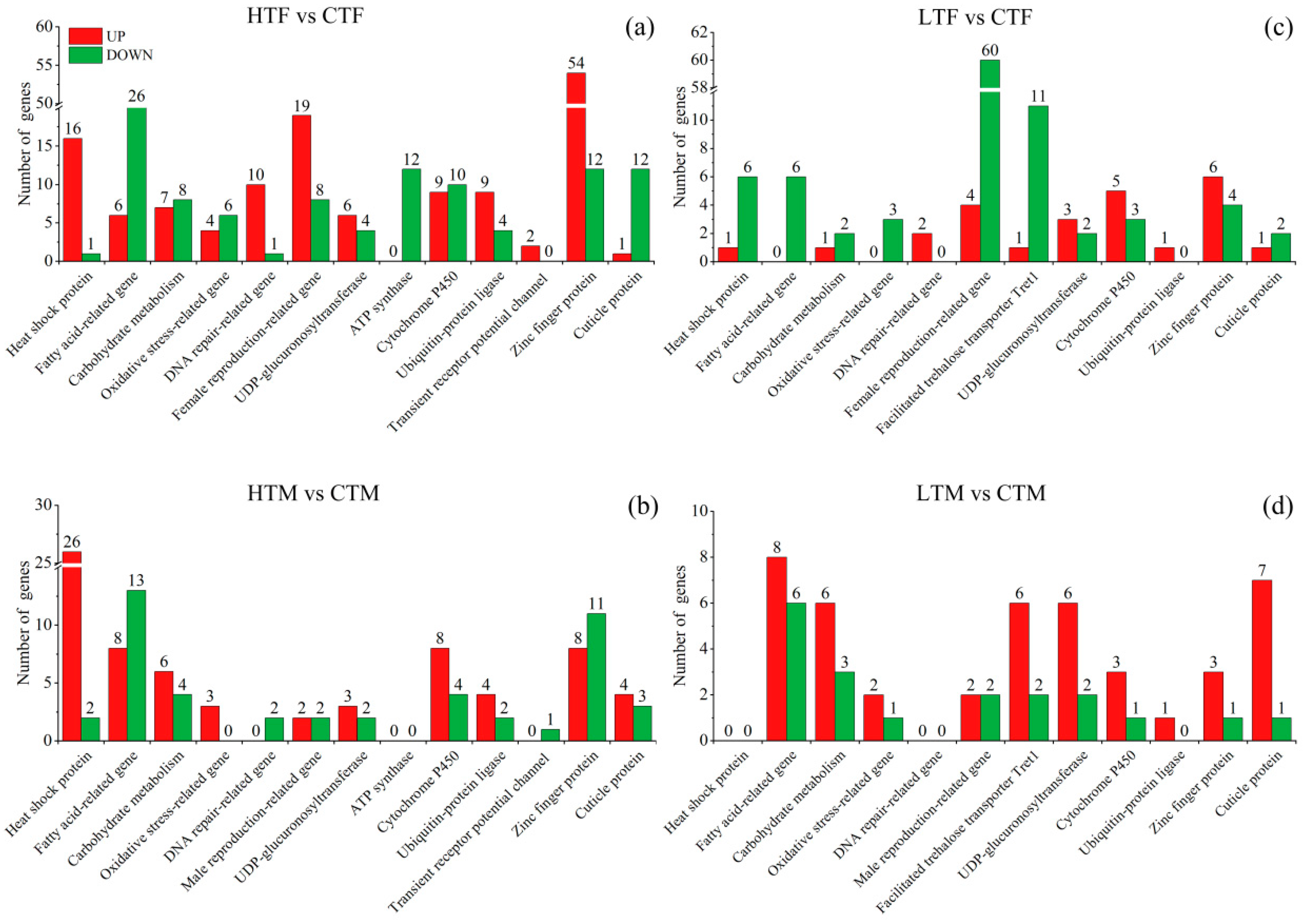
3.4. Transcriptional Changes Induced by Heat Stress in Females
3.5. Transcriptional Changes Induced by Heat Stress in Males
3.6. Transcriptional Changes Induced by Cold Stress in Females
3.7. Transcriptional Changes Induced by Cold Stress in Males
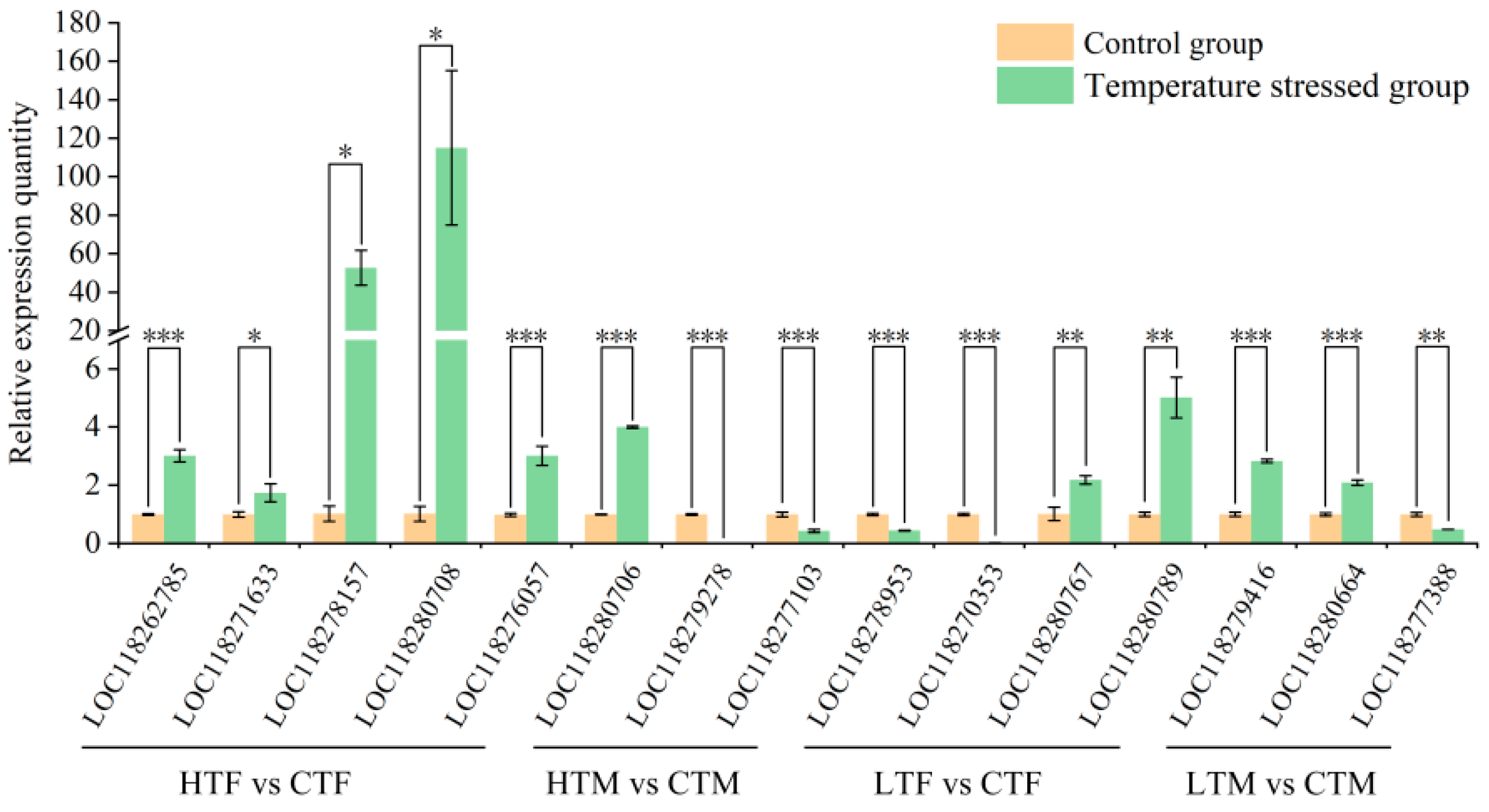
3.8. Validation of RNA-Seq Sequencing Data
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, L.N.; Hu, B.J.; Su, X.Y.; Qi, R.D.; Su, W.H.; Qiu, K.; Zhou, Z.Y.; Zheng, Z.Y.; Zhang, Q.Y.; Hu, F.; et al. Genetic analysis of the fall armyworm Spodoptera frugiperda invaded in Anhui province. Plant Prot. 2019, 45, 47–53. [Google Scholar]
- Jian, Y.Y.; Liu, J.; Wu, Q.L.; Ciren, Z.G.; Zeng, J. Investigation on winter breeding and over wintering areas of Spodoptera frugiperda in China. Plant Prot. 2021, 47, 212–217. [Google Scholar]
- Zanne, A.E.; Flores-Moreno, H.; Powell, J.R.; Cornwell, W.K.; Dalling, J.W.; Austin, A.T.; Classen, A.T.; Eggleton, P.; Okada, K.-I.; Parr, C.L.; et al. Termite sensitivity to temperature affects global wood decay rates. Science 2022, 377, 1440–1443. [Google Scholar] [CrossRef] [PubMed]
- Varga, K.; Elliott, K.W.; Nordyke, C.; Sreter, J.A.; Jovic, K.; Brown, L.M. Biophysical characterization and applications of an insect antifreeze protein. Biophys. J. 2023, 122, 11a. [Google Scholar] [CrossRef]
- Vatanparast, M.; Park, Y. Differential transcriptome analysis reveals genes related to low- and high-temperature stress in the fall armyworm, Spodoptera frugiperda. Front. Physiol. 2022, 12, 827077. [Google Scholar] [CrossRef] [PubMed]
- Purać, J.; Kojić, D.; Petri, E.; Popović, Ž.D.; Grubor-Lajšić, G.; Blagojević, D.P. Cold adaptation responses in insects and other arthropods: An “omics” approach. In Short Views on Insect Genomics and Proteomics: Insect Proteomics; Raman, C., Goldsmith, M.R., Agunbiade, T.A., Eds.; Springer International Publishing: Berlin, Germany, 2016; Volume 2, pp. 89–112. [Google Scholar]
- Gonzalez-Tokman, D.; Cordoba-Aguilar, A.; Dattilo, W.; Lira-Noriega, A.; Sanchez-Guillen, R.A.; Villalobos, F. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world. Biol. Rev. 2020, 95, 802–821. [Google Scholar] [CrossRef] [PubMed]
- Nover, L.; Scharf, K.D. Heat stress proteins and transcription factors. Cell Mol. Life Sci. 1997, 53, 80–103. [Google Scholar] [CrossRef]
- Carmel, J.; Rashkovetsky, E.; Nevo, E.; Korol, A. Differential expression of small heat shock protein genes Hsp23 and Hsp40, and heat shock gene Hsr-omega in fruit flies (Drosophila melanogaster) along a microclimatic gradient. J. Hered. 2011, 102, 593–603. [Google Scholar] [CrossRef]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. Annu. Rev. Entomol. 2015, 60, 59–75. [Google Scholar] [CrossRef]
- Matsuura, H.; Sokabe, T.; Kohno, K.; Tominaga, M.; Kadowaki, T. Evolutionary conservation and changes in insect TRP channels. BMC Evol. Biol. 2009, 9, 228. [Google Scholar] [CrossRef]
- Hori, S.; Tateyama, M.; Shirai, T.; Kubo, Y.; Saitoh, O. Two single-point mutations in Ankyrin Repeat one drastically change the threshold temperature of TRPV1. Nat. Commun. 2023, 14, 2415. [Google Scholar] [CrossRef] [PubMed]
- Adamo, S.A. The effects of the stress response on immune function in invertebrates: An evolutionary perspective on an ancient connection. Horm. Behav. 2012, 62, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.A.B.; Robertson, R.M. A role for octopamine in coordinating thermoprotection of an insect nervous system. J. Therm. Biol. 2006, 31, 149–158. [Google Scholar] [CrossRef]
- Woon, J.S.; Boyle, M.J.W.; Ewers, R.M.; Chung, A.; Eggleton, P. Termite environmental tolerances are more linked to desiccation than temperature in modified tropical forests. Insect. Soc. 2019, 66, 57–64. [Google Scholar] [CrossRef]
- Ferveur, J.-F.; Cortot, J.; Rihani, K.; Cobb, M.; Everaerts, C. Desiccation resistance: Effect of cuticular hydrocarbons and water content in Drosophila melanogaster adults. PeerJ 2018, 6, e4318. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.L.; Guo, Z.Q.; Liu, X.D. Cuticular protein genes involve heat acclimation of insect larvae under global warming. Insect Mol. Biol. 2022, 31, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Dufourc, E.J. Sterols and membrane dynamics. J. Chem. Biol. 2008, 1, 63–77. [Google Scholar] [CrossRef]
- Ma, W.; Li, X.; Shen, J.; Du, Y.; Xu, K.; Jiang, Y. Transcriptomic analysis reveals Apis mellifera adaptations to high temperature and high humidity. Ecotox. Environ. Saf. 2019, 184, 109599. [Google Scholar] [CrossRef]
- Liu, Y.; Su, H.; Li, R.; Li, X.; Xu, Y.; Dai, X.; Zhou, Y.; Wang, H. Comparative transcriptome analysis of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae) reveals novel insights into heat stress tolerance in insects. BMC Genom. 2017, 18, 974. [Google Scholar] [CrossRef]
- Toxopeus, J.; Sinclair, B.J. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef]
- Cha, W.H.; Lee, D.-W. Suppression of glycerol biosynthesis-related genes decreases the effect of rapid cold hardening in Helicoverpa assulta. Entomol. Res. 2022, 52, 24–32. [Google Scholar] [CrossRef]
- Vatanparast, M.; Sajjadian, S.M.; Park, Y. Glycerol biosynthesis plays an essential role in mediating cold tolerance the red imported fire ant, Solenopsis invicta. Arch. Insect. Biochem. 2022, 109, e21861. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Zhang, G.; Nan, J.; Cheng, W.; Zhu-Salzman, K. Characterization of trehalose metabolic genes and corresponding enzymatic activities during diapause of Sitodiplosis mosellana. J. Insect Physiol. 2021, 135, 104324. [Google Scholar] [CrossRef] [PubMed]
- Cha, W.H.; Kim, K.H.; Lee, D.-W. Comparison of gene expression in larval fat body of Helicoverpa assulta in different temperature conditions. Korean J. Appl. Entomol. 2018, 57, 165–175. [Google Scholar]
- Vukašinović, E.L.; Pond, D.W.; Worland, M.R.; Kojić, D.; Purać, J.; Popović, Ž.D.; Grubor-Lajšić, G. Diapause induces remodeling of the fatty acid composition of membrane and storage lipids in overwintering larvae of Ostrinia nubilalis, Hubn. (Lepidoptera: Crambidae). Comp. Biochem. Physiol. B 2015, 184, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Gerhauser, J.; Gaukel, V. Detailed analysis of the ice surface after binding of an insect antifreeze protein and correlation with the Gibbs-Thomson equation. ACS J. Surface. Colloid. 2021, 37, 11716–11725. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Eves, R.; Campbell, R.L.; Davies, P.L. Crystal structure of an insect antifreeze protein reveals ordered waters on the ice-binding surface. Biochem. J. 2020, 477, 3271–3286. [Google Scholar] [CrossRef]
- Goto, S.G.; Philip, B.N.; Teets, N.M.; Kawarasaki, Y.; Lee, R.E., Jr.; Denlinger, D.L. Functional characterization of an aquaporin in the Antarctic midge Belgica antarctica. J. Insect Physiol. 2011, 57, 1106–1114. [Google Scholar] [CrossRef]
- Wu, T.; Cao, D.-H.; Liu, Y.; Yu, H.; Fu, D.-Y.; Ye, H.; Xu, J. Mating-induced common and sex-specific behavioral, transcriptional changes in the moth fall armyworm (Spodoptera frugiperda, Noctuidae, Lepidoptera) in laboratory. Insects 2023, 14, 209. [Google Scholar] [CrossRef]
- Dong, Q.-J.; Zhou, J.-C.; Zhu, K.-H.; Zhang, Z.-T.; Dong, H. A simple method for identifiying sexuality of Spodoptera frugiperda (J. E. Smith) pupae and adults. Plant Prot. 2019, 45, 96–98. [Google Scholar]
- Xiao, H.; Ye, X.; Xu, H.; Mei, Y.; Yang, Y.; Chen, X.; Yang, Y.; Liu, T.; Yu, Y.; Yang, W.; et al. The genetic adaptations of fall armyworm Spodoptera frugiperda facilitated its rapid global dispersal and invasion. Mol. Ecol. Resour. 2020, 20, 1050–1068. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D. The positive false discovery rate: A Bayesian interpretation and the q-value. Ann. Stat. 2003, 31, 2013–2035. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Ganger, M.T.; Dietz, G.D.; Ewing, S.J. A common base method for analysis of qPCR data and the application of simple blocking in qPCR experiments. BMC Bioinform. 2017, 18, 534. [Google Scholar] [CrossRef]
- Holmstrup, M.; Hedlund, K.; Boriss, H. Drought acclimation and lipid composition in Folsomia candida: Implications for cold shock, heat shock and acute desiccation stress. J. Insect Physiol. 2002, 48, 961–970. [Google Scholar] [CrossRef]
- Popović, Ž.D.; Maier, V.; Avramov, M.; Uzelac, I.; Gošić-Dondo, S.; Blagojević, D.; Koštál, V. Acclimations to cold and warm conditions differently affect the energy metabolism of diapausing larvae of the European corn borer Ostrinia nubilalis (Hbn.). Front. Physiol. 2021, 12, 768593. [Google Scholar] [CrossRef]
- Grgac, R.; Rozsypal, J.; Des Marteaux, L.; Štětina, T.; Koštál, V. Stabilization of insect cell membranes and soluble enzymes by accumulated cryoprotectants during freezing stress. Proc. Natl. Acad. Sci. USA 2022, 119, e2211744119. [Google Scholar] [CrossRef]
- Howe, R.W. Temperature effects on embryonic development in insects. Annu. Rev. Entomol. 1967, 12, 15–42. [Google Scholar] [CrossRef]
- Jeffs, C.T.; Leather, S.R. Effects of extreme, fluctuating temperature events on life history traits of the grain aphid, Sitobion avenae. Entomol. Exp. Appl. 2014, 150, 240–249. [Google Scholar] [CrossRef]
- Katsuki, M.; Miyatake, T. Effects of temperature on mating duration, sperm transfer and remating frequency in Callosobruchus chinensis. J. Insect Physiol. 2009, 55, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Gandara, A.C.P.; Drummond-Barbosa, D. Chronic exposure to warm temperature causes low sperm abundance and quality in Drosophila melanogaster. Sci. Rep. 2023, 13, 12331. [Google Scholar] [CrossRef] [PubMed]
- Noer, N.K.; Nielsen, K.L.; Sverrisdottir, E.; Kristensen, T.N.; Bahrndorff, S. Temporal regulation of temperature tolerances and gene expression in an arctic insect. J. Experim. Biol. 2023, 226, jeb245097. [Google Scholar] [CrossRef] [PubMed]
- Waters, J.S.; Harrison, J.F. Insect metabolic rates. In Metabolic Ecology; Sibly, R.M., Brown, J.H., Kodric-Brown, A., Eds.; Wiley-Blackwell: West Sussex, UK, 2012; pp. 198–211. [Google Scholar]
- Frederich, M.; Portner, H.O. Oxygen limitation of thermal tolerance defined by cardiac and ventilatory performance in spider crab, Maja squinado. Am. J. Physiol. Regul.Integr. Comp. Physiol. 2000, 279, R1531–R1538. [Google Scholar] [CrossRef]
- Burton, T.; Killen, S.S.; Armstrong, J.D.; Metcalfe, N.B. What causes intraspecific variation in resting metabolic rate and what are its ecological consequences? Proc. R. Soc. Lond. B Biol. Sci. 2011, 278, 3465–3473. [Google Scholar] [CrossRef] [PubMed]
- Jena, K.; Kar, P.K.; Kausar, Z.; Babu, C.S. Effects of temperature on modulation of oxidative stress and antioxidant defenses in testes of tropical tasar silkworm Antheraea mylitta. J. Therm. Biol. 2013, 38, 199–204. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Metabolic rate depression in animals: Transcriptional and translational controls. Biol. Rev. 2004, 79, 207–233. [Google Scholar] [CrossRef]
- Karl, I.; Stoks, R.; De Block, M.; Janowitz, S.A.; Fischer, K. Temperature extremes and butterfly fitness: Conflicting evidence from life history and immune function. Global. Change Biol. 2011, 17, 676–687. [Google Scholar] [CrossRef]
- Catalan, T.P.; Wozniak, A.; Niemeyer, H.M.; Kalergis, A.M.; Bozinovic, F. Interplay between thermal and immune ecology: Effect of environmental temperature on insect immune response and energetic costs after an immune challenge. J. Insect Physiol. 2012, 58, 310–317. [Google Scholar] [CrossRef]
- Wang, L.; Etebari, K.; Walter, G.H.; Furlong, M.J. Sex dependent transcriptome responses of the diamondback moth, Plutella xylostella L. to cold stress. Comp. Biochem. Physiol. D 2023, 45, 101053. [Google Scholar] [CrossRef]
- Thomas, M.B.; Blanford, S. Thermal biology in insect-parasite interactions. Trends Ecol. Evol. 2003, 18, 344–350. [Google Scholar] [CrossRef]
- Chen, Y.-R.; Jiang, T.; Zhu, J.; Xie, Y.-C.; Tan, Z.-C.; Chen, Y.-H.; Tang, S.-M.; Hao, B.-F.; Wang, S.-P.; Huang, J.-S.; et al. Transcriptome sequencing reveals potential mechanisms of diapause preparation in bivoltine silkworm Bombyx mori (Lepidoptera: Bombycidae). Comp. Biochem. Physiol. D 2017, 24, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, X.; Qiao, H.; He, X.; Tan, J.; Hao, D. Comparative transcriptome analysis of the heat stress response in Monochamus alternatus Hope (Coleoptera: Cerambycidae). Front. Physiol. 2020, 10, 1568. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, J.; Yang, Y.; Lu, Y. Transcriptome analysis of Propylaea quatuordecimpunctata L. (Coleoptera: Coccinellidae) under high temperature stress. Agriculture 2022, 12, 1088. [Google Scholar] [CrossRef]
- Schmitt, A.; Nebreda, A.R. Signalling pathways in oocyte meiotic maturation. J. Cell Sci. 2002, 115, 2457–2459. [Google Scholar] [CrossRef]
- Will, T.; Schmidtberg, H.; Skaljac, M.; Vilcinskas, A. Heat shock protein 83 plays pleiotropic roles in embryogenesis, longevity, and fecundity of the pea aphid Acyrthosiphon pisum. Dev. Genes. Evol. 2017, 227, 1–9. [Google Scholar] [CrossRef]
- Okada, Y.; Teramura, K.; Takahashi, K.H. Heat shock proteins mediate trade-offs between early-life reproduction and late survival in Drosophila melanogaster. Physiol. Entomol. 2014, 39, 304–312. [Google Scholar] [CrossRef]
- Kumar, M.; Gupta, G.P.; Rajam, M.V. Silencing of acetylcholinesterase gene of Helicoverpa armigera by siRNA affects larval growth and its life cycle. J. Insect Physiol. 2009, 55, 273–278. [Google Scholar] [CrossRef]
- Huang, L.-H.; Wang, C.-Z.; Kang, L. Cloning and expression of five heat shock protein genes in relation to cold hardening and development in the leafminer, Liriomyza sativa. J. Insect Physiol. 2009, 55, 279–285. [Google Scholar] [CrossRef]
- Malkeyeva, D.; Kiseleva, E.; Fedorova, S. Small heat shock protein Hsp67Bc plays a significant role in Drosophila melanogaster cold stress tolerance. J. Exp. Biol. 2020, 223, jeb219592. [Google Scholar] [CrossRef]
- Jia, D.; Ji, Z.; Yuan, X.; Zhang, B.; Liu, Y.; Hu, J.; Wang, Y.; Li, X.; Ma, R. Molecular cloning and expression profiles of thermosensitive TRP genes in Agasicles hygrophila. Insects 2020, 11, 531. [Google Scholar] [CrossRef] [PubMed]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschella, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 17071. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Maruyama, K.; Sakuma, Y.; Meshi, T.; Iwabuchi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis Cys2/His2-type zinc-finger proteins function as transcription repressors under drought, cold, and high-salinity stress conditions. Plant. Physiol. 2004, 136, 2734–2746. [Google Scholar] [CrossRef] [PubMed]
- Ciftci-Yilmaz, S.; Morsy, M.R.; Song, L.; Coutu, A.; Krizek, B.A.; Lewis, M.W.; Warren, D.; Cushman, J.; Connolly, E.L.; Mittler, R. The EAR-motif of the Cys2/His2-type zinc finger protein Zat7 plays a key role in the defense response of Arabidopsis to salinity stress. J. Biol. Chem. 2007, 282, 9260–9268. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.-j.; Wang, L.-j.; Wang, C.; Guo, D.-z.; Xu, B.-h.; Guo, X.-q.; Li, H. Identification of an Apis cerana zinc finger protein 41 gene and its involvement in the oxidative stress response. Arch. Insect. Biochem. 2021, 108, e21830. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-R.; Shan, Y.-M.; Tan, Y.; Zhang, Z.-R.; Pang, B.-P. Comparative analysis of transcriptome responses to cold stress in Galeruca daurica (Coleoptera: Chrysomelidae). J. Insect Sci. 2019, 19, 8. [Google Scholar] [CrossRef] [PubMed]
- Dunning, L.T.; Dennis, A.B.; Park, D.; Sinclair, B.J.; Newcomb, R.D.; Buckley, T.R. Identification of cold-responsive genes in a New Zealand alpine stick insect using RNA-Seq. Comp. Biochem. Physiol. D 2013, 8, 24–31. [Google Scholar] [CrossRef]
- Huang, H.-J.; Xue, J.; Zhuo, J.-C.; Cheng, R.-L.; Xu, H.-J.; Zhang, C.-X. Comparative analysis of the transcriptional responses to low and high temperatures in three rice planthopper species. Mol. Ecol. 2017, 26, 2726–2737. [Google Scholar] [CrossRef]
- Cui, M.; Hu, P.; Wang, T.; Tao, J.; Zong, S. Differential transcriptome analysis reveals genes related to cold tolerance in seabuckthorn carpenter moth, Eogystia hippophaecolus. PLoS ONE 2017, 12, e0187105. [Google Scholar] [CrossRef]
- Lou, Y.H.; Pan, P.L.; Ye, Y.X.; Cheng, C.; Xu, H.J.; Zhang, C.X. Identification and functional analysis of a novel chorion protein essential for egg maturation in the brown planthopper. Insect Mol. Biol. 2018, 27, 393–403. [Google Scholar] [CrossRef]
- Paul, S.; Keshan, B. Ovarian development and vitellogenin eene expression under heat stress in silkworm, Bombyx mori. Psyche 2016, 2016, 4242317. [Google Scholar]
- Chen, J.; Du, X.; Xu, X.; Zhang, S.; Yao, L.; He, X.; Wang, Y. Comparative proteomic analysis provides new insights into the molecular basis of thermal-induced parthenogenesis in silkworm (Bombyx mori). Insects 2023, 14, 134. [Google Scholar] [CrossRef] [PubMed]
- Yapici, N.; Kim, Y.J.; Ribeiro, C.; Dickson, B.J. A receptor that mediates the post-mating switch in Drosophila reproductive behaviour. Nature 2008, 451, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, N.; Hua, Y.J.; Roller, L.; Spalovska-Valachova, I.; Mizoguchi, A.; Kataoka, H.; Tanaka, Y. Bombyx prothoracicostatic peptides activate the sex peptide receptor to regulate ecdysteroid biosynthesis. Proc. Natl. Acad. Sci. USA. 2010, 107, 2060–2065. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Sun, L.-l.; Liu, P.; Cao, C.-w. The sex peptide receptor in the Asian gypsy moth, Lymantria dispar, is involved in development and stress resistance. J. Integ. Agric. 2021, 20, 2976–2985. [Google Scholar] [CrossRef]
- Michaud, M.R.; Denlinger, D.L. Shifts in the carbohydrate, polyol, and amino acid pools during rapid cold-hardening and diapause-associated cold-hardening in flesh flies (Sarcophaga crassipalpis): A metabolomic comparison. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2007, 177, 753–763. [Google Scholar] [CrossRef]
- Rodriguez, E.; Weber, J.-M.; Darveau, C.-A. Diversity in membrane composition is associated with variation in thermoregulatory capacity in hymenopterans. Comp. Biochem. Physiol. B 2018, 224, 115–120. [Google Scholar] [CrossRef]
- Khalil, S.; El-Gamal, S.; Ibrahim, S.; Elateek, S. Characterization, expression analysis and RNAi-mediated knockdown of two aquaporin genes in the cotton leafworm, Spodoptera littoralis (Lepidoptera: Noctuidae). Eur. J. Entomol. 2023, 120, 15–25. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Y.-D.; Liu, Y.; Wan, X.-S.; Xu, J.; Fu, D.-Y.; Zhang, J.-Z. High and Low Temperatures Differentially Affect Survival, Reproduction, and Gene Transcription in Male and Female Moths of Spodoptera frugiperda. Insects 2023, 14, 958. https://doi.org/10.3390/insects14120958
Tao Y-D, Liu Y, Wan X-S, Xu J, Fu D-Y, Zhang J-Z. High and Low Temperatures Differentially Affect Survival, Reproduction, and Gene Transcription in Male and Female Moths of Spodoptera frugiperda. Insects. 2023; 14(12):958. https://doi.org/10.3390/insects14120958
Chicago/Turabian StyleTao, Yi-Dong, Yu Liu, Xiao-Shuang Wan, Jin Xu, Da-Ying Fu, and Jun-Zhong Zhang. 2023. "High and Low Temperatures Differentially Affect Survival, Reproduction, and Gene Transcription in Male and Female Moths of Spodoptera frugiperda" Insects 14, no. 12: 958. https://doi.org/10.3390/insects14120958