3.1. Habitat Characteristics
In the Swengel dataset (
Table 1),
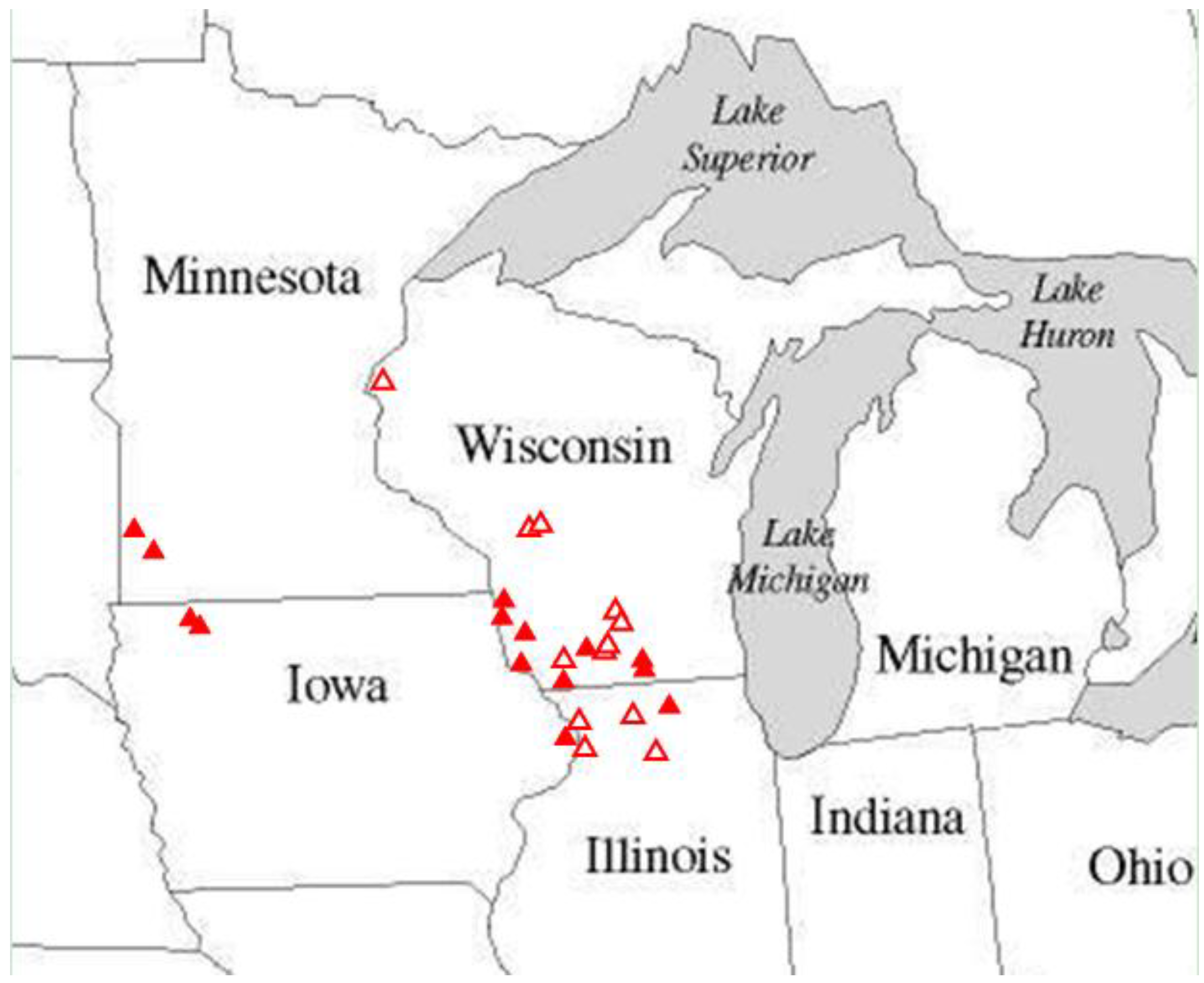
H. ottoe occurred only in dry upland and sand prairie. All Illinois and Wisconsin sites in this study (
Table 2) contained only those prairie types. The Iowa and Minnesota sites (
Table 2) contained both lowland and upland prairie. The few records there in the Swengel dataset and all records attributable to a vegetative type in the other datasets occurred only in upland prairie. In the Swengel dataset,
H. ottoe was significantly more abundant in undegraded prairie (
Table 1), and all sites where we found
H. ottoe contained undegraded dry or sand prairie.
H. ottoe was also significantly more abundant in sites with discontinuous sod (with numerous unvegetated areas due to bare sand and/or rock outcrops) than in continuous sod (with few or no small bare spots and/or rocks). Small sites had higher
H. ottoe abundance compared to medium and large sites. When the sample was limited to high-quality prairie analyzed separately by sod type, the preference for small sites persisted, significantly so for discontinuous sod, which had the greater range of site sizes. Because most populations in this study declined to subdetectability either before or during Swengel surveys (
Table 2), habitat analysis (
Table 1) was limited to extant sites only in Wisconsin in the years the species was still detectable, in order to make any habitat preferences more identifiable statistically. We recorded no
H. ottoe at a number of other sites appearing to contain appropriate vegetation (
Table 3).
Table 1.
Mean ± SD of H. ottoe individuals per hr in Wisconsin on unit surveys in the Swengel dataset. Analysis only includes years at each site before subdetection occurred (if subdetection occurred). Within each vegetative characteristic, variates sharing any of the same letters are not statistically different by the Mann-Whitney U test (two-tailed p < 0.05).
Table 1.
Mean ± SD of H. ottoe individuals per hr in Wisconsin on unit surveys in the Swengel dataset. Analysis only includes years at each site before subdetection occurred (if subdetection occurred). Within each vegetative characteristic, variates sharing any of the same letters are not statistically different by the Mann-Whitney U test (two-tailed p < 0.05).
| Prairie type | N Unit surveys | Mean observation rate | SD | Statistical grouping |
|---|
| Prairie type |
| dry prairie | 549 | 4.58 | 13.31 | A |
| “extra dry” sand prairie | 71 | 2.74 | 14.08 | A |
| Prairie quality |
| degraded | 40 | 0.43 | 1.06 | AB |
| semi-degraded | 241 | 1.81 | 6.49 | B |
| undegraded | 340 | 6.64 | 16.93 | A |
| Type of sod 1 |
| continuous | 385 | 2.20 | 7.26 | B |
| discontinuous | 236 | 7.90 | 19.16 | A |
| Prairie size 2 |
| Small | 107 | 14.29 | 24.71 | A |
| medium | 377 | 2.01 | 6.65 | B |
| large | 137 | 3.10 | 11.20 | B |
| Prairie size 2 limited to high-quality continuous sod |
| small | 18 | 7.49 | 14.72 | A |
| medium | 168 | 2.21 | 6.46 | A |
| Prairie size 2 limited to high-quality discontinuous sod |
| small | 65 | 20.27 | 29.07 | A |
| medium | 9 | 5.20 | 7.84 | AB |
| large | 80 | 4.48 | 14.33 | B |
| Management |
| idling | 38 | 1.17 | 2.90 | AB |
| grazing | 96 | 0.49 | 1.13 | B |
| manual cutting | 5 | 0.00 | 0.00 | AB |
| fire+mow/graze | 69 | 2.45 | 8.00 | AB |
| Fire | 413 | 5.94 | 15.83 | A |
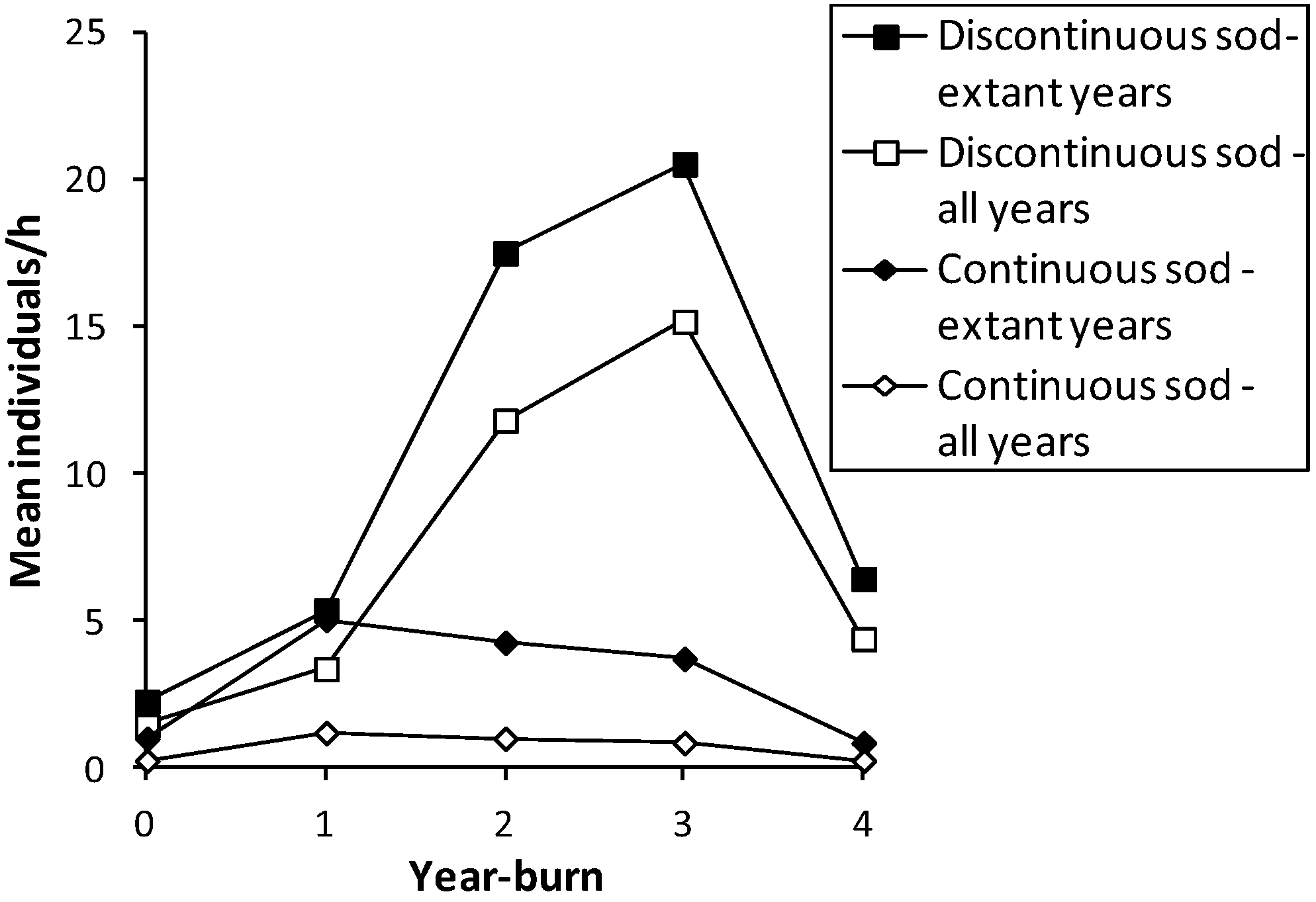
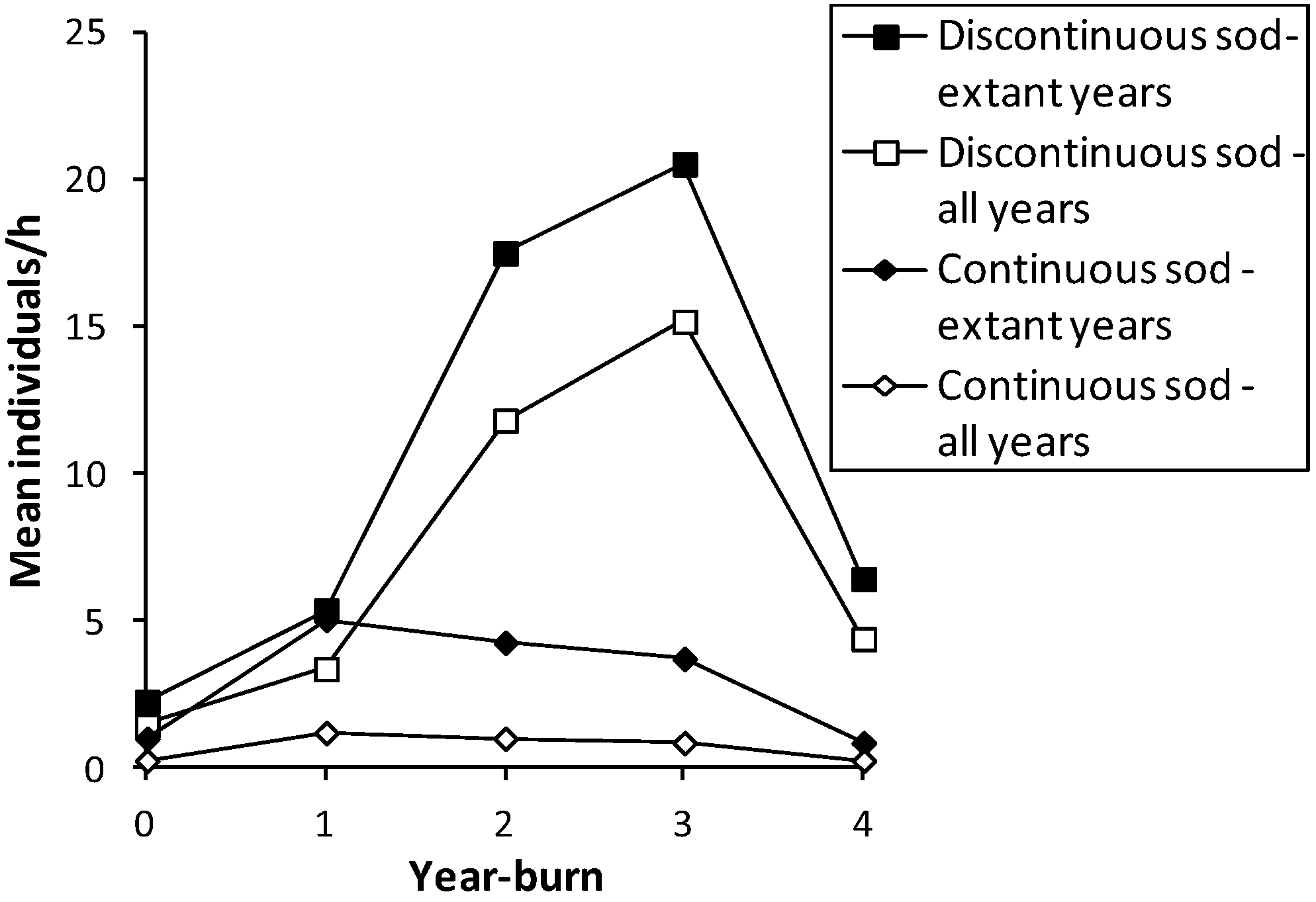
Figure 1.
Mean abundance (individuals/h per unit survey) of
H. ottoe in each year-burn after fire in all survey years before subdetection at each site in the Swengel dataset (see
Table 2), by continuous or discontinuous sod.
Figure 1.
Mean abundance (individuals/h per unit survey) of
H. ottoe in each year-burn after fire in all survey years before subdetection at each site in the Swengel dataset (see
Table 2), by continuous or discontinuous sod.
Table 2.
Study sites where H. ottoe was recorded in this multi-team dataset through 2011. This table identifies the last known year the species was detected (it may not have been detected in all years visited prior to this year), and the first and last year and number of years when all visits had no detection (called “subdetection”).
Table 2.
Study sites where H. ottoe was recorded in this multi-team dataset through 2011. This table identifies the last known year the species was detected (it may not have been detected in all years visited prior to this year), and the first and last year and number of years when all visits had no detection (called “subdetection”).
| Site | State | Year preserved | Last year detected | N years detected | 1st year subdetectable | N years to subdetection | Year of last survey | N years of subdetection |
|---|
| Continuous sod |
| Cayler 1 | IA | 1971 | 1974 | 3 | 1980 | 9 | 2005 | 8 |
| Haffner | IA | 1976 | 1989 | 13 | 1994 | 18 | 2005 | 4 |
| Harlem Hill | IL | 1973 | 1993 | 20 | 1994 | 21 | 1997 | 3 |
| Hole-in-the- Mountain (new) | MN | 1990 | 1991 | 1 | 1992 | 2 | 2006 | 6 |
| Hole-in-the- Mountain (old) | MN | 1978 | 1995 | 17 | 1996 | 18 | 2007 | 4 |
| Prairie Coteau | MN | 1986 | 1995 | 9 | 1996 | 10 | 2007 | 6 |
| Hardscrabble 2 | WI | 1997 | | | 2006 | 9 | 2011 | 4 |
| Hogback | WI | 1997 | 2011 | 14 | | | 2011 | 0 |
| Muralt Bluff | WI | 1977 | 1997 | 20 | 1998 | 21 | 2011 | 14 |
| Oliver | WI | 1962 | 1992 | 30 | 1993 | 31 | 2011 | 19 |
| Discontinuous sod |
| Manikowski | IA | 1985 | 2008 | 23 | | | 2008 | 0 |
| Battle Bluff | WI | 1983 | 2011 | 28 | | | 2011 | 0 |
| Dewey Heights | WI | 1952 | 2007 | 55 | 2008 | 56 | 2011 | 4 |
| N Rush Creek | WI | 1981 | 2007 | 26 | 2008 | 27 | 2008 | 1 |
| Rush Creek | WI | 1981 | 2011 | 30 | | | 2011 | 0 |
| Spring Green 3 | WI | 1972 | 1996 | 24 | 1997 | 25 | 2011 | 6 |
Table 3.
Sites of plausible habitat in Illinois and Wisconsin for
H. ottoe surveyed during its flight period in 173.6 h and 403.8 km of survey effort, but with no records of the species in this study. See also Swengel and Swengel [
8] for surveying at numerous sites of plausible habitat in Iowa, Minnesota, and North Dakota, but with no.
H. ottoe recorded.
Table 3.
Sites of plausible habitat in Illinois and Wisconsin for H. ottoe surveyed during its flight period in 173.6 h and 403.8 km of survey effort, but with no records of the species in this study. See also Swengel and Swengel [8] for surveying at numerous sites of plausible habitat in Iowa, Minnesota, and North Dakota, but with no. H. ottoe recorded.
| Site | State | Years surveyed | Latitude | Longitude | Size (ha) |
|---|
| Ayers Sand Prairie | IL | 1991–1993 | 42.06 | 90.10 | 47 |
| Bicentennial Prairie | IL | 1991–1993 | 42.13 | 89.20 | 3 |
| Nachusa Prairie | IL | 1991–1996 | 41.88 | 89.35 | 162 |
| Thomson-Fulton Prairie | IL | 1991–1993 | 41.92 | 90.11 | 15 |
| Bauer-Brockway Barrens | WI | 1991–2012 | 44.29 | 90.75 | 97 |
| Black Earth Rettenmund Prairie | WI | 1990–1993 | 43.14 | 89.77 | 4 |
| Blue River Cactus Flats | WI | 1991–1993 | 43.15 | 90.53 | 36 |
| Crex Meadows | WI | 1991–2012 | 45.88 | 92.60 | 7000 |
| Dike 17 | WI | 1990–2012 | 44.31 | 90.56 | 4 |
| Schluckebier Prairie | WI | 1991–1993 | 43.29 | 89.79 | 9 |
| Thomson | WI | 1990–2012 | 42.98 | 89.83 | 40 |
| Thousand's II | WI | 1990–2012 | 42.98 | 89.84 | 2 |
H. ottoe nectared on a wide variety of primarily native flowers (
Table 4). We observed oviposition (
i.e., confirmed an egg was laid) on a blade of
Schizachyrium scoparium (little bluestem) three times. We observed oviposition behavior but did not confirm an egg got laid on the underside of
Andropogon gerardii (big bluestem), on little bluestem, and on a big bluestem blade and adjacent forb foliage.
The significantly higher numbers of
H. ottoe in undegraded prairie vegetation (
Table 1) confirms its categorization at the highest level of prairie dependence [
30]. On surveys in 2004–2005 at two conserved sites and adjacent private land in western Iowa,
H. ottoe significantly related positively with warm-season (native) grasses and bare ground, and negatively with cool-season (primarily non-native) grasses and litter depth [
31]. Others have noted the association of
H. ottoe with high-quality native prairie with short or sparse grass and often some bare areas [
32,
33,
34]. However, the species’ larval food plants are common native prairie grasses [
6,
30,
35] and its nectar sources are widely occurring prairie flowers (
Table 4; [
6,
8]). Undegraded dry or upland prairie, often rocky or sandy, that
H. ottoe associates with (
Table 1; 30) is more widely occurring (even as an intact undegraded prairie flora) than
H. ottoe (
Table 2 and
Table 3). Discontinuous sod affords some protection from lethal heat during a fire since the bare sand and/or rock outcrops may prevent some areas from combusting, as evidenced by significantly higher numbers in year-burn 0 in discontinuous sod (
Figure 1). As a result, it is unclear whether the significantly higher abundance in discontinuous than continuous sod (
Table 1) relates to a vegetative preference or to differential fire mortality.
H. ottoe abundance had a negative relationship to prairie size (
Table 1) and both measures of population persistence correlated negatively with prairie size (
Table 5). This is counter to metapopulation theory that population incidence and abundance should increase in larger and more connected habitat patches [
37,
38]. Although there was not a significant difference in prairie size between continuous and discontinuous sites, the continuous-sod sites averaged more than double the size of the discontinuous-sod sites (
Table 6), and
H. ottoe was significantly more abundant in discontinuous than continuous sod (
Table 1). However, when controlling for sod type, the significant negative relationship to patch size persisted in discontinuous sod (
Table 1). We are unable to identify a mechanism for smaller site size to benefit
H. ottoe. Instead, it appears that patch size has a synergistic relationship with other unfavorable factors, such as tall thick turf and management (discussed below).
3.2. Fire Management
H. ottoe was significantly more abundant in fire-managed sites than in the grazed site (Hogback), but otherwise management did not produce significant effects (
Table 1).
H. ottoe was under-represented in year-burn 0 compared to an expected distribution proportional to amount of survey effort there (
Table 7). However,
H. ottoe was also over-represented in units ever fire-managed compared to non-fire-managed units (
Table 8). When limiting the sample to extant years only,
H. ottoe was significantly more abundant in discontinuous than continuous sod in both year-burn 0 and year-burn 2+ (
Figure 1; Mann-Whitney U tests of unit surveys). When including all survey years, this pattern persisted significantly for year-burn 2+ and nearly so (
p < 0.06) in year-burn 0. In pair-wise tests within each sod type, year-burns 0, 1, and 2+ were not significantly different from one another in skipper abundance. No tests of abundance were significant when year-burn 2+ was subdivided into year-burns 2, 3, and 4+. However, year of peak mean abundance post-fire occurred later in discontinuous sod (year-burn 3) and was much higher than in continuous sod (year-burn 1) (
Figure 1). In discontinuous sod, year-burn 0 averaged a lower abundance than in all subsequent year-burns, but in continuous sod, both the soonest and longest since fire had similarly lowest average abundances (
Figure 1).
H. ottoe was significantly under-represented in year-burn 0 (
Table 7,
Figure 1) but also declined in later years of the fire rotation (
Figure 1) and was over-represented in fire-managed units compared to non-fire-managed units (
Table 1 and
Table 8). Likewise, in Vogel
et al.’s [
31] study,
H. ottoe abundance was significantly higher in burn-only units (1–3 fires during 2000–2004) and significantly lower in both cattle-graze-only (light and rotational) and burned+grazed units (1–4 fires during 1997–2004). In Vogel
et al. [
39],
H. ottoe peaked in abundance 30 months after fire in highly variable counts plotted out to a maximum of 60 months.
However, as most populations declined to subdetection in fire-managed sites (
Table 2), these results do not endorse burning as an effective long-term management for this skipper.
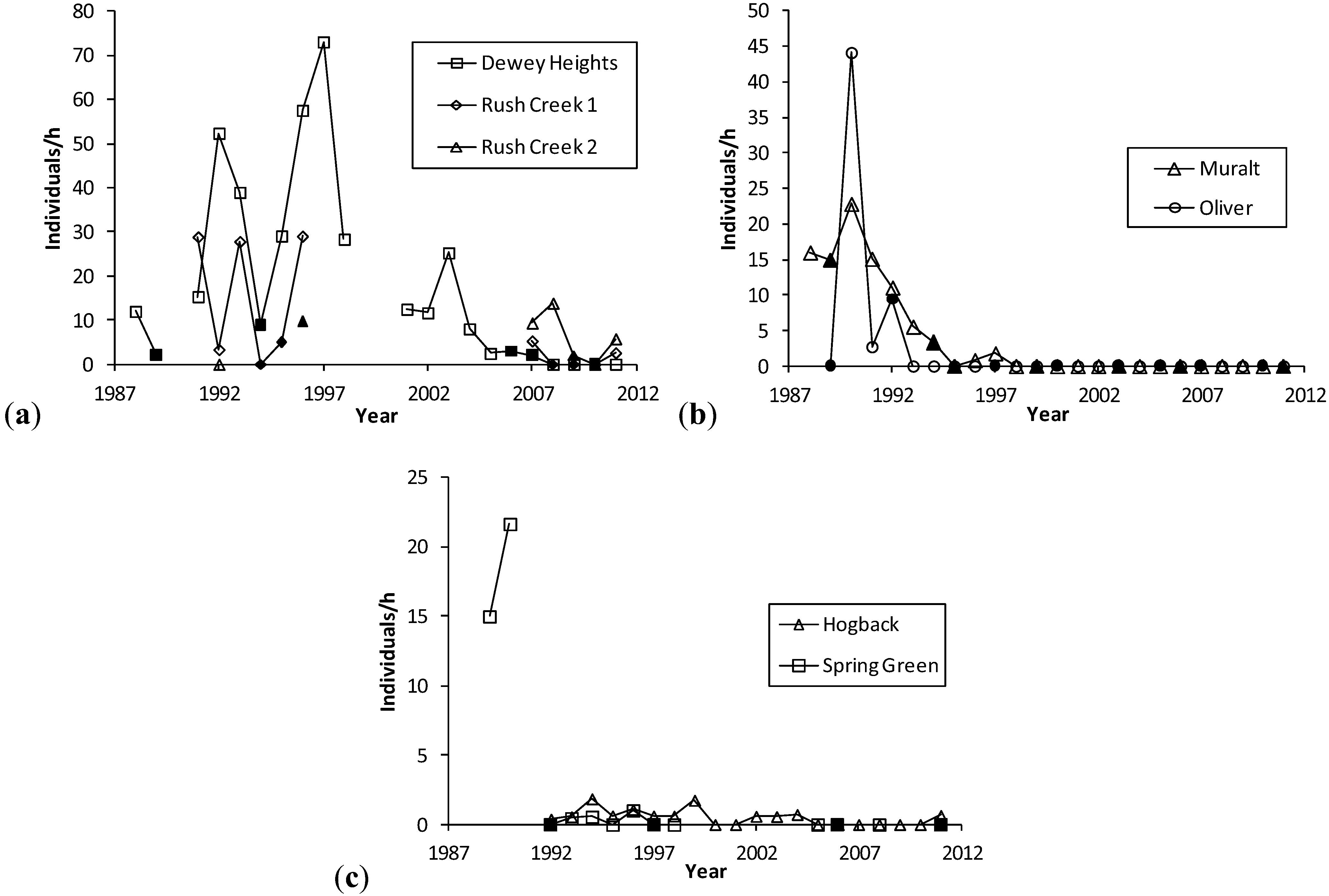
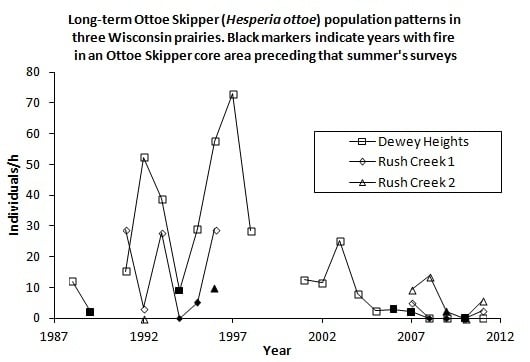
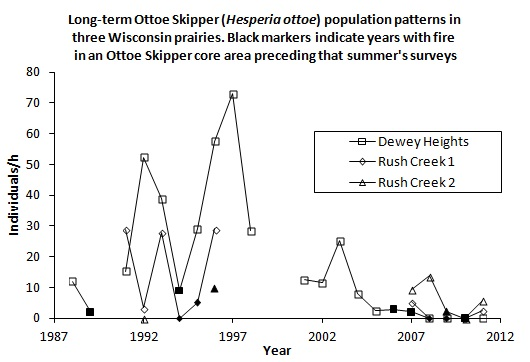
H. ottoe often persisted for some years after the start of fire management, which was usually rotational and partial except that most or all of small sites were more likely to be burned at once during our surveys (e.g., Oliver and Dewey Heights). After a substantial fire in a core area (Oliver and Spring Green in 1992, Muralt Bluff in 1994, Dewey Heights in 2006–2007) the population still existed but then did not recover and became subdetectable within a few years, before the core area burned again (
Figure 2). Battle Bluff, one of the few extant sites, has a permanent non-fire refugium in core habitat, according to management staff [
40] because the area is too steep for managers to work in safely. Likewise, specialist butterfly abundance was significantly higher in fire-managed sites when they contained a permanent non-fire refugium in core habitat [
12,
41], which might have unintensive management if needed to control brush and weeds (e.g., mowing problems areas).
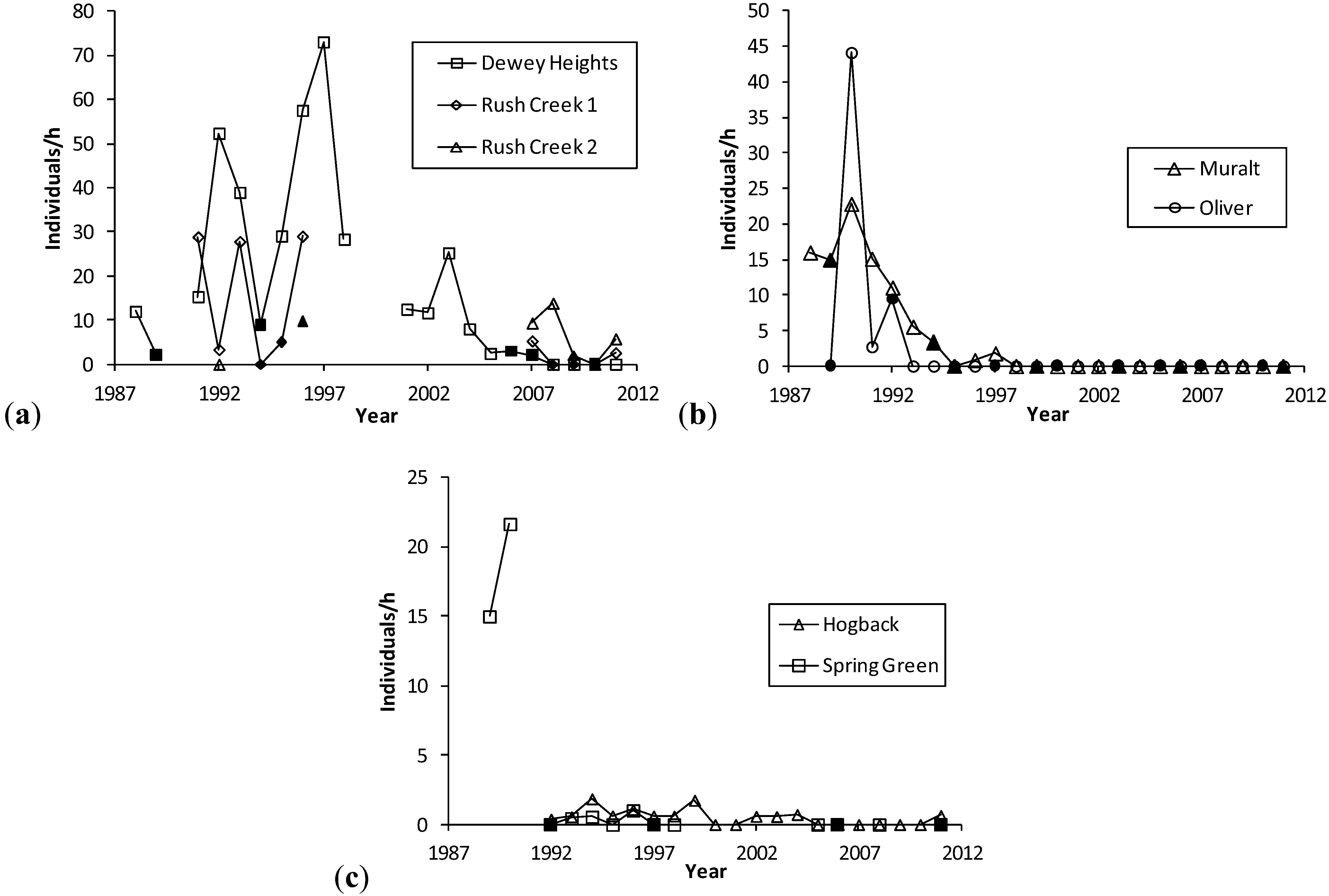
Figure 2.
(a), (b), (c). H. ottoe individuals per h per year, by site in the Swengel dataset. Dark markers indicate a fire in part or all of core H. ottoe habitat in the cool season preceding that summer's survey. The Spring Green Survey in 1991 had H. ottoe but occurred at end of flight period and is not graphed here.
Figure 2.
(a), (b), (c). H. ottoe individuals per h per year, by site in the Swengel dataset. Dark markers indicate a fire in part or all of core H. ottoe habitat in the cool season preceding that summer's survey. The Spring Green Survey in 1991 had H. ottoe but occurred at end of flight period and is not graphed here.
In one experiment, Dana [
6] placed reared
H. ottoe larvae in enclosures in spring. Adult emergences were significantly higher in unburned plots than in plots subsequently burned, with no significant difference between medium and heavy fuel or early and late spring burns. In the next experiment, adult emergences from stocked larvae were significantly lower in plots with moderate fuel that were subsequently burned compared to both burns with light fuel and unburned controls, and in late spring compared to early spring for both fuel loads. Dana [
6] concluded that the prior experiment was flawed and based on the latter experiment, that early spring fires in low to moderate fuels typical of dry prairie have mortality too low for concern, while late spring fires of moderate to heavy fuel could be devastating. However,
H. ottoe has declined and disappeared greatly in burned sites (including Dana’s study site), much more so than that latter experiment would predict. Dana’s [
6] earlier experiment (where early spring burns were not more favorable than late spring ones) may be as representative of preserve conditions as the later experiment. Alternatively, a sufficient number of fires may have occurred in these sites in the most unfavorable circumstances (later and heavier fuel conditions). Also, as discussed by Dana [
6], ongoing fire management can result in grasses increasing and becoming thicker and taller, leading to less favorable grassland structure for
H. ottoe as well as heavier fuels.
Table 4.
N nectar visits by
H. ottoe during 1988-2012, by Wisconsin site and sex (m = male, f = female, u = unknown). Botanical nomenclature follows Cochrane and Iltis [
36]. * = observed in late 1980s; not quantified.
Table 4.
N nectar visits by H. ottoe during 1988-2012, by Wisconsin site and sex (m = male, f = female, u = unknown). Botanical nomenclature follows Cochrane and Iltis [36]. * = observed in late 1980s; not quantified.
| Site Code 1 | B | Dewey | Hog | Muralt | Oliver | Rush | Spring | Total |
|---|
| Nectar plant | m | m | f | u | m | f | m | f | m | f | u | m | f | u | m | f | u | |
| Amorpha canescens (leadplant) | | | | | | | 1 | | | | | | | | | 1 | | 2 |
| Asclepias syriaca | | | | | | | | | | | | 2 | 1 | | | | | 3 |
| (common milkweed) | | | | | | | | | | | | | | | | | | |
| Asclepias tuberosa | | 2 | | | | | | | | | | | | | | | | 2 |
| (butterfly weed) | | | | | | | | | | | | | | | | | | |
| Asclepias viridiflora | 1 | 4 | 2 | | | | | | | | | | | | | | | 7 |
| (green milkweed) | | | | | | | | | | | | | | | | | | |
| Carduus acanthoides | | | | | 3 | | | 1 | | | | | | | | | | 4 |
| (welted thistle) | | | | | | | | | | | | | | | | | | |
| Cirsium hillii (Hill’s thistle) | | | | | 4 | 1 | 1 | 1 | | | | | | | | | | 7 |
| Cirsium (thistle) | | | | | | | | | | | | | | | | | 1 | 1 |
| Coreopsis palmata | 2 | 10 | 1 | | | | | | 3 | | | 21 | 1 | | | | | 38 |
| (prairie coreopsis) | | | | | | | | | | | | | | | | | | |
| Dalea purpurea | | 4 | | | | | | | | | | | | | 1 | 2 | | 7 |
| (purple prairie clover) | | | | | | | | | | | | | | | | | | |
| Erigeron strigosus | | | | | | | 1 | | | | | | | | | 1 | | 2 |
| (daisy fleabane) | | | | | | | | | | | | | | | | | | |
| Euphorbia corollata | | 1 | | | | | | | | | | | | | | | | 1 |
| (flowering spurge) | | | | | | | | | | | | | | | | | | |
| Liatris cylindracea | | | | | | | 5 | 5 | 2 | 3 | 1 | 3 | | | | | | 19 |
| (dwarf blazingstar) | | | | | | | | | | | | | | | | | | |
| Monarda fistulosa | 1 | 39 | 17 | 3 | 1 | | 4 | 3 | | | | | | | | | | 68 |
| (wild bergamot) | | | | | | | | | | | | | | | | | | |
| Opuntia (prickly pear) | | | | | | | | | | | | | | | * | | | * |
| Pycnanthemum | | | | | | 2 | | | | | | | | | | | | 2 |
| (mountain mint) | | | | | | | | | | | | | | | | | | |
| Ratibida pinnata | | 1 | 1 | | | | 1 | | | | | | | | | | | 3 |
| (yellow coneflower) | | | | | | | | | | | | | | | | | | |
| Rudbeckia hirta | | | 3 | | | | 4 | 1 | | | | | | | | | | 8 |
| (black-eyed Susan) | | | | | | | | | | | | | | | | | | |
| Silphium integrifolium | | | | | | | | | 2 | | | | | | | | | 2 |
| (rosinweed) | | | | | | | | | | | | | | | | | | |
| Silphium laciniatum | | | | | | | | | | | | | | 1 | | | | 1 |
| (compass plant) | | | | | | | | | | | | | | | | | | |
| Solidago ptarmicoides | | | | | | | | | | 1 | | | | | | | | 1 |
| (upland white aster) | | | | | | | | | | | | | | | | | | |
| Solidago rigida (stiff goldenrod) | | | | | | | 1 | 3 | | | | | | | | | | 4 |
| Tephrosia virginiana (goat’s rue) | | | | | | | | | | | | | | | 1 | | | 1 |
| Verbena stricta (hoary vervain) | | 1 | | | | | 1 | | | | | | | | | | | 2 |
| Total nectar records | 4 | 62 | 24 | 3 | 8 | 3 | 19 | 14 | 7 | 4 | 1 | 26 | 2 | 1 | 2 | 4 | 1 | 185 |
Table 5.
Spearman rank correlation coefficients (r) of patch size and year preserved with N years detected and N years to subdetection, by sod type (continuous or discontinuous), as in
Table 2.
Table 5.
Spearman rank correlation coefficients (r) of patch size and year preserved with N years detected and N years to subdetection, by sod type (continuous or discontinuous), as in Table 2.
| Site traits | Continuous sod | | Discontinuous sod | |
|---|
| N sites | r | p | N sites | r | p |
|---|
| Patch size | | | | | | |
| N years detected | 9 | −0.705 | <0.05 | 6 | −0.261 | >0.10 |
| N years to subdetection | 9 | −0.540 | >0.10 | 3 | −0.500 | - |
| Year preserved | | | | | | |
| N years detected | 9 | −0.435 | >0.10 | 6 | −0.551 | >0.10 |
| N years to subdetection | 9 | −0.633 | <0.07 | 3 | −0.500 | - |
Table 6.
Mean ± SD, median, and range of year preserved and prairie size (ha), by continuous or discontinuous sod as in
Table 2. Mann-Whitney U test two-tailed
p > 0.20 for both.
Table 6.
Mean ± SD, median, and range of year preserved and prairie size (ha), by continuous or discontinuous sod as in Table 2. Mann-Whitney U test two-tailed p > 0.20 for both.
| Site traits | N sites | mean | SD | median | Range |
|---|
| Year preserved | | | | | |
| Continuous | 10 | 1981 | 11.5 | 1978 | 1962–1997 |
| Discontinuous | 6 | 1976 | 12.4 | 1981 | 1952–1985 |
| Prairie size (ha) | | | | | |
| Continuous | 10 | 57.1 | 62.9 | 20.2 | 1.6–157.9 |
| Discontinuous | 6 | 22.5 | 16.2 | 24.3 | 1.2–38.5 |
Table 7.
Number of H. ottoe individuals observed in the Swengel dataset by whether unit surveys were in the first growing season after burning or not, and expected individuals based on proportion of effort surveying in each sample, only at sites where the species was ever recorded in this dataset. p = 0.0000 in both Chi Square Goodness of Fit tests.
Table 7.
Number of H. ottoe individuals observed in the Swengel dataset by whether unit surveys were in the first growing season after burning or not, and expected individuals based on proportion of effort surveying in each sample, only at sites where the species was ever recorded in this dataset. p = 0.0000 in both Chi Square Goodness of Fit tests.
| Year-burn status | N observed | Time surveying | Distance surveying |
|---|
| h | % | N expected | km | % | N expected |
|---|
| Year-burn 0 | 33 | 32.69 | 15.7 | 105 | 63.21 | 17.8 | 119 |
| Not year-burn 0 | 637 | 175.28 | 84.3 | 565 | 291.59 | 82.2 | 551 |
Table 8.
Number of H. ottoe individuals observed in Swengel dataset by whether unit surveys were in fire management or not, and expected individuals based on proportion of effort surveying in each sample, only at sites where the species was ever recorded in this dataset. p = 0.0000 in both Chi Square Goodness of Fit tests.
Table 8.
Number of H. ottoe individuals observed in Swengel dataset by whether unit surveys were in fire management or not, and expected individuals based on proportion of effort surveying in each sample, only at sites where the species was ever recorded in this dataset. p = 0.0000 in both Chi Square Goodness of Fit tests.
| Management | N observed | Time surveying | Distance surveying |
|---|
| h | % | N expected | km | % | N expected |
|---|
| Fire | 622 | 148.54 | 71.2 | 477 | 273.77 | 77.2 | 517 |
| Non-fire | 48 | 60.20 | 28.8 | 193 | 80.95 | 22.8 | 153 |
Vogel
et al. [
39] identified the direct effect of fire (mortality) as a stronger statistical effect on
H. ottoe abundance than indirect (vegetative) effects. This agrees with the implication of direct mortality in our study because of the significantly lower abundance in year-burn 0 in continuous than discontinuous sod, which affords some firebreak (
Figure 1). However, Vogel
et al. [
31,
39] as well as Dana [
6] studied
H. ottoe early in the conservation management history of their study sites, well before the median half-life of
H. ottoe populations in preserves with discontinuous sod (27 years) and continuous sod (14–18 years), respectively (
Table 9). Our study suggests that later in the fire history at a preserve, the indirect (vegetative) effects may be more decisive in population outcome because subdetection began in a year when fire did not occur in core habitat (
Figure 2).
Table 9.
Analysis of population detectability after preservation based on
Table 2: N sites in samples and mean, median, and SE of N years (detected or to subdetection), and two-tailed
p for Mann-Whitney U test between continuous and discontinuous sod.
Table 9.
Analysis of population detectability after preservation based on Table 2: N sites in samples and mean, median, and SE of N years (detected or to subdetection), and two-tailed p for Mann-Whitney U test between continuous and discontinuous sod.
| Sod type | N years detected | N years to subdetection |
|---|
| Sites | mean | median | SE | p | Sites | mean | median | SE | p |
|---|
| Continuous | 9 | 14.1 | 14 | 3.01 | 0.008 1 | 9 | 15.4 | 18 | 2.91 | 0.042 |
| Discontinuous | 6 | 31.0 | 27 | 6.91 | | 3 | 36.0 | 27 | 10.02 | |
In Vogel
et al. [
31], burn-only units had significantly higher warm season (native) grass cover and bare ground (
Table 3: mean 20.49%) and lower litter and exotic forbs, while the other two management types had significantly higher cool season (primarily non-native) grass cover. No pre-treatment measurements were available, as is often the case in many conservation management studies, including all sites but Hogback in this study. Without pre-treatment data, it is not possible to identify what floristic characteristics are a response to conservation management, as opposed to a skewing of conservation management decisions based on pre-existing floristic characteristics. That is, fire-only management may be focused on flora that was higher quality upon preservation, while grazing may be focused on areas that were more degraded upon preservation. Thus, differences among plots in flora may not be attributable to conservation management type but instead to conservation preferences in what managements should be implemented in what kinds of flora.
3.3. Other Management Types
Idling also did not appear favorable for
H. ottoe (
Table 1). Sites often have brush or weeds at the time of idling and if unaddressed by management, the native floristic quality
H. ottoe prefers (
Table 1) will deteriorate. The small sample for idling could be skewed toward areas deemed lower priority for conservation while the fire-managed sample may be skewed toward areas of higher quality vegetation of higher priority to conservation managers. However, idling could also be unfavorable because it also allows the turf to thicken. As a result, long periods of idling appear more suitable for
H. ottoe in sites of discontinuous sod such as Battle Bluff, Dewey Heights, and Rush Creek, where the harsh microclimate and rocky soil favor short sparse prairie flora. However, data are not sufficient to determine whether even in those circumstances, idled herbaceous prairie flora is sufficiently compatible with long-term persistence of
H. ottoe.
Mowing (either leaving the clippings lie, or removing them as hay) can be relatively favorable for many other prairie-specialist butterflies, including grass-skippers [
8,
15,
17,
42]. However, this has not appeared to be a historical management in the
H. ottoe sites in this study, which do not appear amenable to this agricultural use due to terrain and low vegetative productivity. Theoretically, mowing and brush cutting could be useful for reducing brush and weeds and keeping turf shorter. However, no studies known to us document how
H. ottoe populations have responded to this management.
Moderate to heavy grazing appears highly detrimental [
43], both to the skippers themselves as well as the vegetation they require, and may contribute to the spotty historical distribution of
H. ottoe. For example, county records are remarkably few in central Iowa and Minnesota [
44,
45], given the species’ range in all neighboring states as well as Manitoba. Vogel
et al. [
27] found only two individuals in somewhat degraded grazed grassland and only 13 in burned + grazed grassland.
But some
H. ottoe populations persisted pre-conservation in grazing that was presumably lighter and/or more heterogeneous than typical. Hogback was grazed prior to conservation and had a small population consistently detected each year during 1992−1997 (
Figure 2). Upon conservation in 1998, the unit with
H. ottoe had grazing removed and was idled, with no burning during this study.
H. ottoe has become less detectable with more time since grazing was removed (
Figure 2: detections in 1998−2000, 2002−2004, and 2011). Likewise, McCabe and Post [
46] reported very low numbers in grazed areas of the western Dakotas, but found none on the tops of isolated, ungrazed buttes. The two Minnesota sites in this study (
Table 2) were grazed prior to conservation [
6,
47]. Hole-in-the-Mountain had a large
H. ottoe population at and immediately after conservation [
6]; during this period, a substantial area (“new” part in
Table 2) later found to be occupied by
H. ottoe had not been conserved yet. Prairie Coteau had low
H. ottoe numbers in 1989−1990 in surveys conducted in 1988−1990 [
48,
49], shortly after the site was conserved in 1986 [
50]. No reports are available for the species’ occurrence before and immediately after conservation. Neither site has a record after 1995 ([
51];
Table 2). Relatively few data exist on how
H. ottoe responds to the plethora of possible grazing regimes (species of livestock, intensity and duration of grazing, and frequency if done in rotation).
3.4. Population Status and Trend
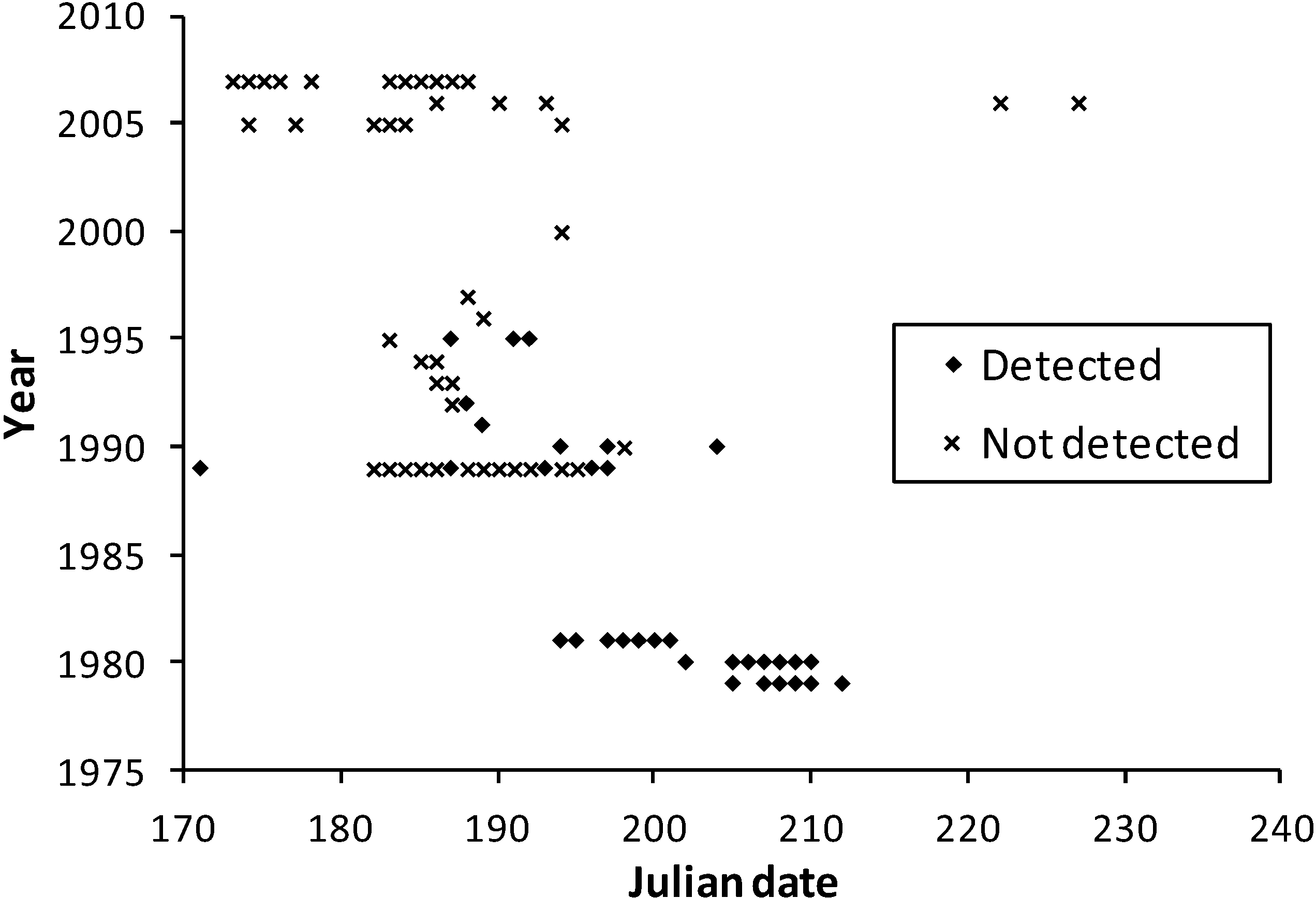
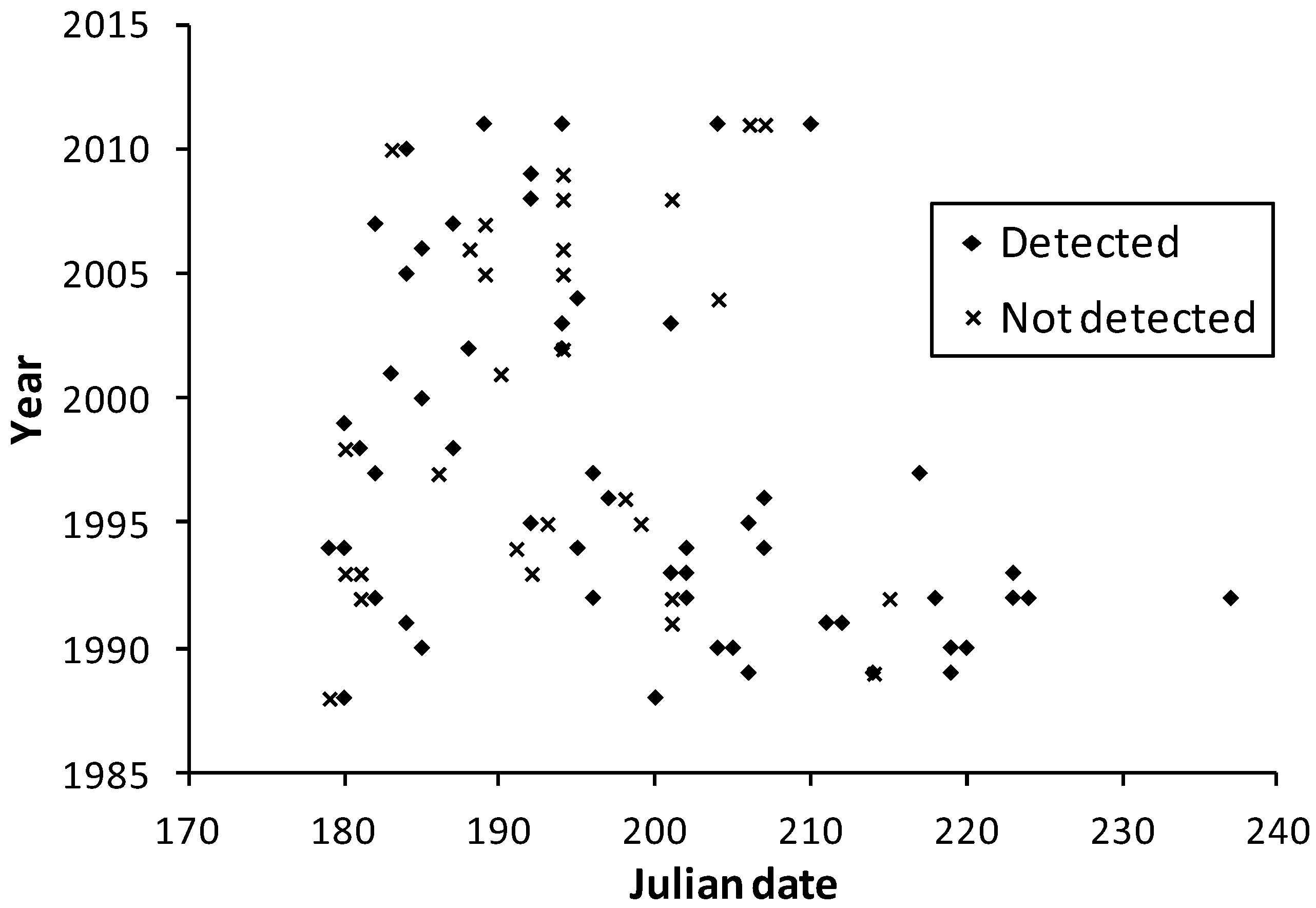
Observation dates in the multi-team dataset were similar between Minnesota and Wisconsin (
Figure 3 and
Figure 4), although many more survey dates and longer survey periods per year occurred in Wisconsin. The few records for Illinois (21 July 1993) and Iowa (1 July 1989, 27 July 2005, 24 June 2008) are consistent with the dates in Minnesota and Wisconsin (
Figure 3 and
Figure 4).
Only 1/9 sites (11%) with continuous sod had detectable
H. ottoe in recent years (
Table 2: Hogback in 2011). In discontinuous sod, 2/6 (33%) did, with two other sites lacking data after 2008. The number of years the species was still detectable after preservation and the number of years to subdetection were both significantly higher in discontinuous than continuous sod (
Table 9). Both measures of population persistence averaged more than twice as long in discontinuous than continuous sod. All pair-wise correlations of both measures of population persistence with prairie size and year of preservation were negative (
Table 5), one significantly so. Year preserved and prairie size did not differ significantly between continuous and discontinuous sod (
Table 6), although sites with continuous sod averaged more than double the size of discontinuous sites.
Abundance varied greatly within site among years prior to population subdetectability (
Figure 2). Years when a core area for
H. ottoe was burned tended to have abundances at the low end of the range observed in the years immediately before and after these fires. However, low abundances also occurred in other years. The pattern of increasing abundance in the first years following some fires, then a subsequent decrease longer since fire (
Figure 1), was apparent in these site time-series (
Figure 2). At Dewey Heights (
Figure 2), a discontinuous sod site, peak abundances occurred in 1992 and 1996 (year-burn 3) and then fell the following year (year-burn 4). At both Muralt Bluff and Oliver (
Figure 2), which are continuous sod sites within 2 km of each other, peak abundance occurred in 1990 (year-burn 1 in the core habitat) and dropped the next year (year-burn 2). At Muralt Bluff, the drop continued each year through 1995 even though fires in the core area did not occur again until 1994−1995.
Figure 3.
Survey dates in the multi-team dataset for Minnesota at H. ottoe sites by whether the species was detected or not.
Figure 3.
Survey dates in the multi-team dataset for Minnesota at H. ottoe sites by whether the species was detected or not.
Figure 4.
Survey dates in the meta-analysis dataset for Wisconsin at H. ottoe sites by whether the species was detected or not.
Figure 4.
Survey dates in the meta-analysis dataset for Wisconsin at H. ottoe sites by whether the species was detected or not.
Most populations did not become subdetectable in year-burn 0 in core habitat. That is, although numbers were low as a result of the fire, the populations survived the fire but then failed to recover in subsequent years of the fire rotation, as had occurred in prior cycles. For populations detected in 1990, these stalled recoveries and subsequent population failures occurred in the mid-1990s at Muralt Bluff (1996–1997), Oliver (1993), and Spring Green (1993–1996) and in the late 2000s at Dewey Heights (2008). At Hogback, no fire has been observed in the core H. ottoe habitat. The population occurred in very low numbers but was detected each year 1992–1999, then became unreliably detectable after that, being recorded only in 2002–2004 and 2011. The site was lightly grazed by cattle through 1997. Then in 1998, the site was preserved and grazing removed.
Continuous sod sites had more rapid declines after preservation and fewer extant populations than discontinuous sod sites (
Table 2 and
Table 9).
H. ottoe populations tended to persist longer after preservation where rocky outcrops and/or sparse fuels afforded more firebreak, fires have been relatively less frequent (e.g., Rush Creek), and/or some occupied habitat was in a permanent non-fire refugium (e.g., Battle Bluff) (
Table 9). Although
H. ottoe declines to subdetectability have been occurring for decades on preserves (1980–2008 in
Table 2), the year subdetection began exhibits some clustering, especially in the mid-1990s. Climatic variation may interact with management to result in particularly adverse circumstances for skipper breeding success. For example, the widespread very wet growing season in 1993 [
13] may have resulted in unfavorably dense and tall grass turfs. Furthermore, the data suggest (non-significantly) that
H. ottoe may have declined to subdetection more quickly in more recently preserved sites (
Table 5). More recently preserved sites may have had more vulnerable populations, or were managed more intensively, or the landscape context has become more hostile in more recent years, adversely affecting new and old preserves alike.
Since these skippers can have relatively high abundance in recently burned units (
Figure 1), Dana [
6] recommended a fire regime in which there would always be a unit <3 years since last fire. To reduce risk from fire mortality on population viability, he advised ensuring that core skipper habitat be placed into at least two units, with one of these units burned every two years (
i.e., a four-year rotation of these units). Dana [
6] also anticipated the possible long-term negative indirect impact of fire (thick tall grass). He noted that episodic brief heavy grazing or rotational haying might be necessary to maintain the appropriate shorter grass structure. The long-term data series in our study bear out this concern, both for the removal of grazing at Hogback and for the longer-term effects of fire at other sites (
Figure 2). These long-term effects are particularly challenging because they seem to present a sharp dichotomy: the prior fire rotation supports an expectation of future recoveries and then no recovery at all occurs following the next fire (
Figure 2). Thus, by the time the effect sets in (hypothesized here to be an unsuitably tall and dense grass structure), there is very little time to act before population failure.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}