Elucidating Structure-Bioactivity Relationships of Methyl-Branched Alkanes in the Contact Sex Pheromone of the Parasitic Wasp Lariophagus distinguendus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Synthesis of Reference Chemicals
2.2.1. General Methods and Information for Synthesis
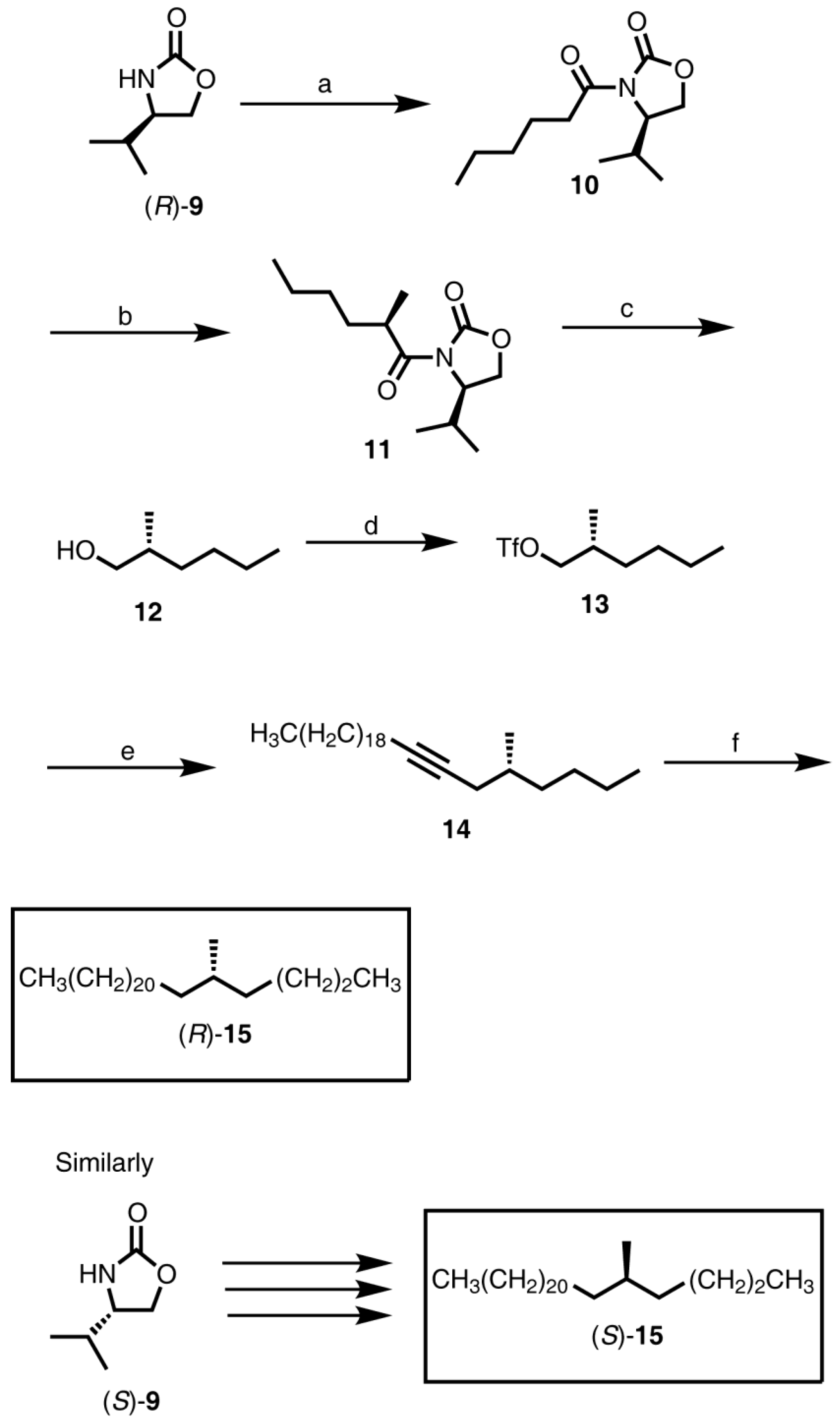
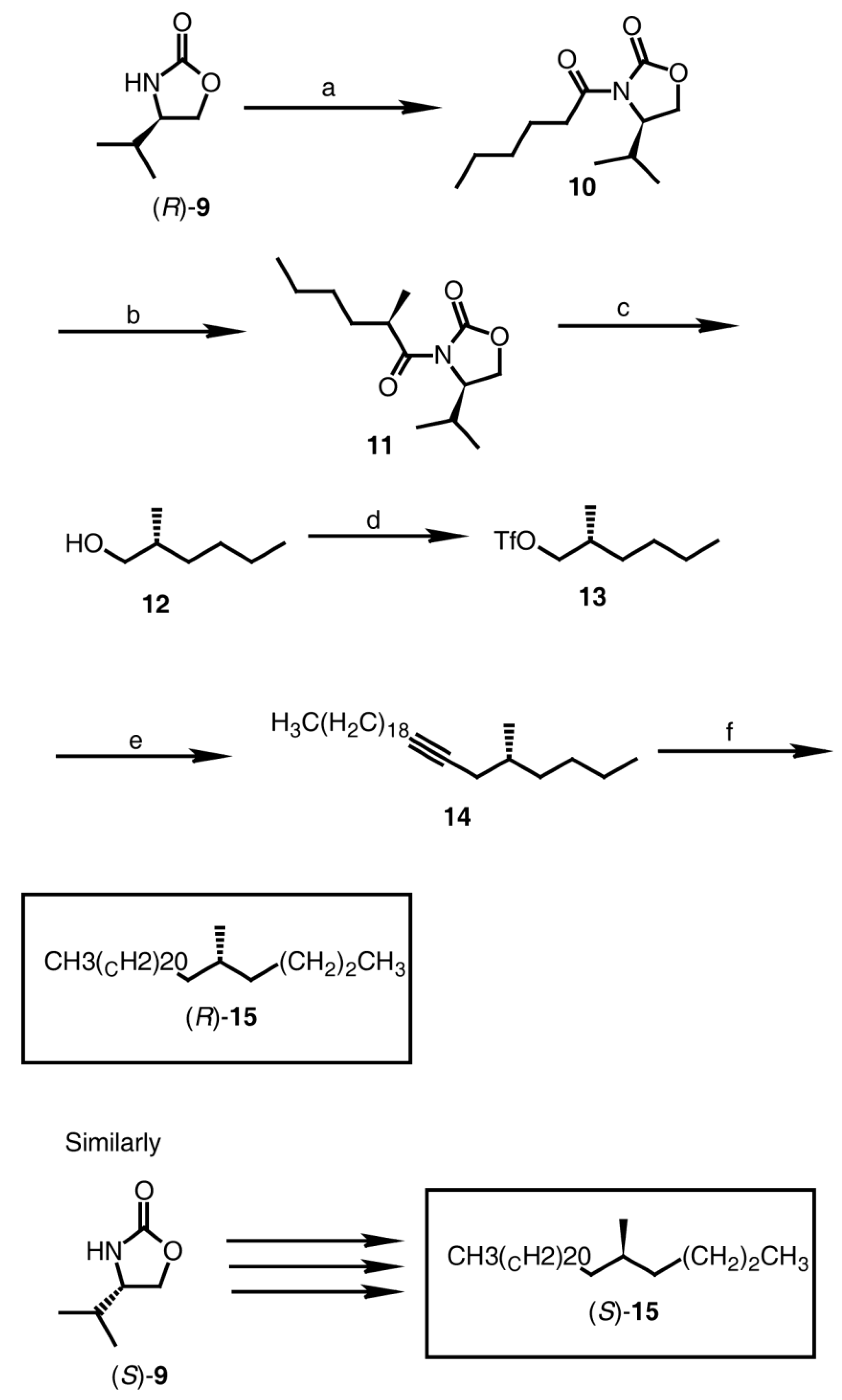
2.2.2. Synthesis of (S)-3-Methylnonacosane [(S)-7], (S)-3-Methylhentriacontane [(S)-8], (R)-3-Methylnonacosane [(R)-7], and (R)-3-Methylhentriacontane [(R)-8] (Figure 1)

2.2.3. Synthesis of (R)-5-Methylheptacosane [(R)-15] and (S)-5-Methylheptacosane [(S)-15] (Figure 2)

2.2.4. Synthesis of (R)- and (S)-7-Methylheptacosane
2.2.5. Synthesis of (R)-and (S)-3-Methylpentacosane, and (R)- and (S)-3-Methylheptacosane
2.3. Bioassays
2.3.1. General Procedures for Bioassays
2.3.2. Experiment 1: Structure-Bioactivity Relationship of Methylalkanes for the Restoration of the Pheromone in 4-d-Old Male Dummies
2.3.3. Experiment 2: Interruption of Pheromone Activity in 0-d-Old Male and Female Dummies by the Addition of Individual CHCs
2.3.4. Statistical Analysis
3. Results
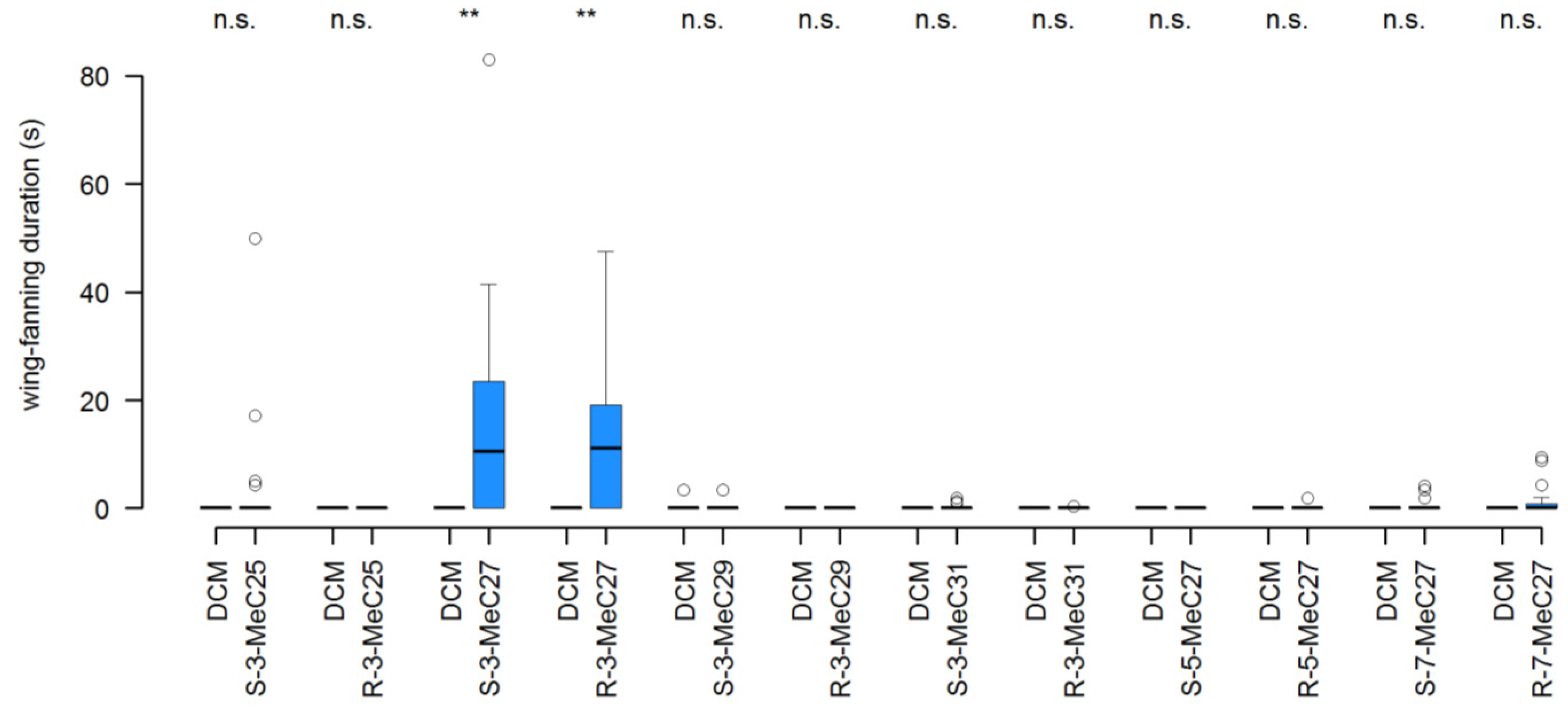
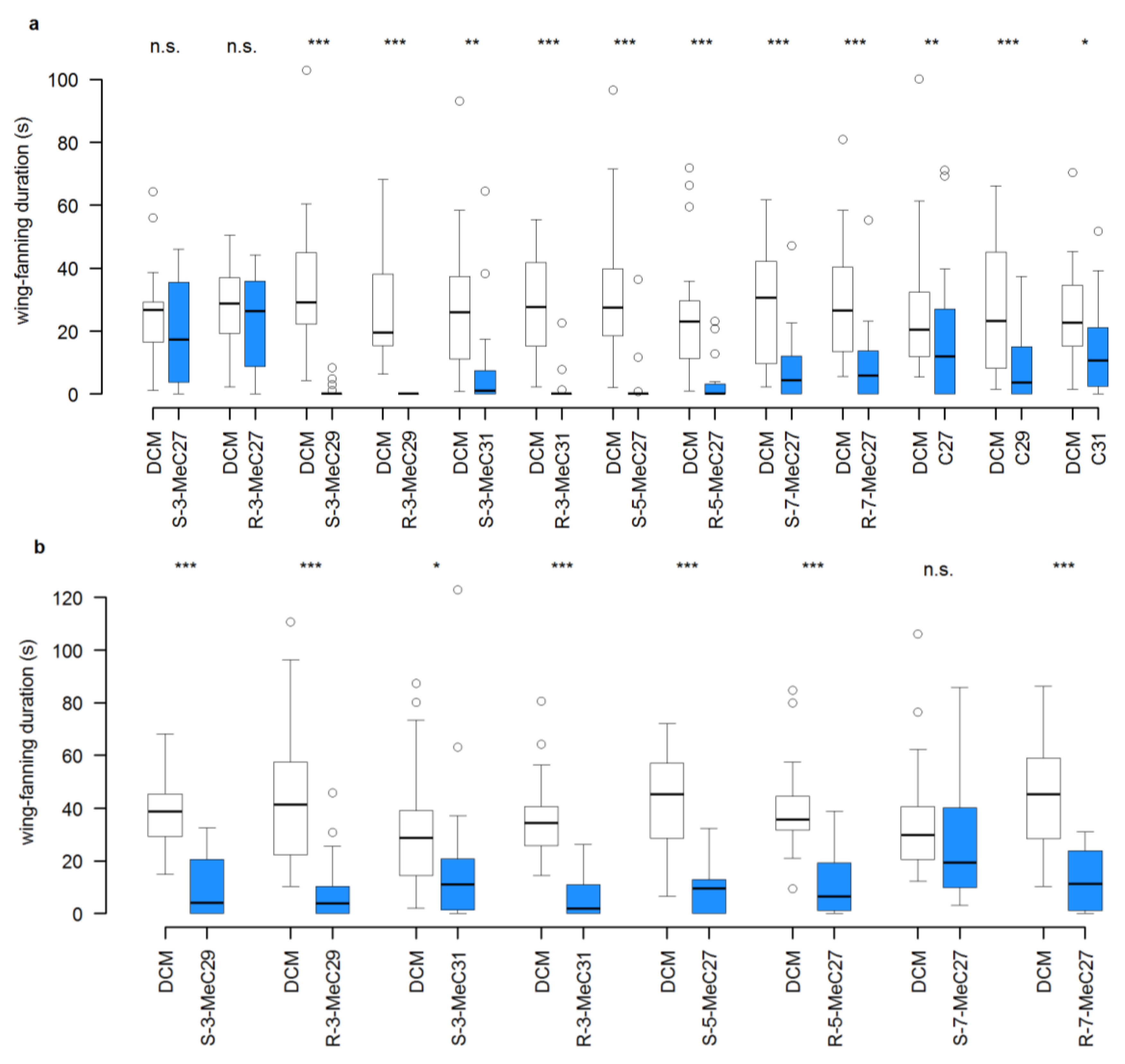
3.1. Experiment 1: Structure-Bioactivity Relationship of Methylalkanes for the Restoration of the Pheromone in 4-d-Old Male Dummies

3.2. Experiment 2: Interruption of Pheromone Activity in 0-d-Old Male and Female Dummies by the Addition of Individual CHCs

4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Buckner, J.S. Oxygenated derivatives of hydrocarbons. In Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology; Blomquist, G.J., Bagnères, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 187–204. [Google Scholar]
- Kühbandner, S.; Sperling, S.; Mori, K.; Ruther, J. Deciphering the signature of cuticular lipids with contact sex pheromone function in a parasitic wasp. J. Exp. Biol. 2012, 215, 2471–2478. [Google Scholar] [CrossRef]
- Yew, J.Y.; Dreisewerd, K.; de Oliveira, C.C.; Etges, W.J. Male-specific transfer and fine scale spatial differences of newly identified cuticular hydrocarbons and triacylglycerides in a Drosophila species pair. PLoS One 2011, 6, e16898. [Google Scholar]
- Mori, K. Pheromone synthesis. Part 253: Synthesis of the racemates and enantiomers of triglycerides of male Drosophila fruit flies with special emphasis on the preparation of enantiomerically pure 1-monoglycerides. Tetrahedron 2012, 68, 8441–8449. [Google Scholar] [CrossRef]
- Gibbs, A.G. Lipid melting and cuticular permeability: New insights into an old problem. J. Insect Physiol. 2002, 48, 391–400. [Google Scholar] [CrossRef]
- Howard, R.W.; Blomquist, G.J. Chemical ecology and biochemistry of insect hydrocarbons. Annu. Rev. Entomol. 1982, 27, 149–172. [Google Scholar] [CrossRef]
- Singer, T.L. Roles of hydrocarbons in the recognition systems of insects. Am. Zool. 1998, 38, 394–405. [Google Scholar]
- Blomquist, G.J.; Bagnères, A.G. Introduction: History and overview of insect hydrocarbons. In Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology; Howard, R.W., Blomquist, G.J., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 3–18. [Google Scholar]
- Dani, F.R.; Jones, G.R.; Destri, S.; Spencer, S.H.; Turillazzi, S. Deciphering the recognition signature within the cuticular chemical profile of paper wasps. Anim. Behav. 2001, 62, 165–171. [Google Scholar] [CrossRef]
- Van Wilgenburg, E.; Sulc, R.; Shea, K.J.; Tsutsui, N.D. Deciphering the chemical basis of nestmate recognition. J. Chem. Ecol. 2010, 36, 751–758. [Google Scholar] [CrossRef]
- Dani, F.R.; Jones, G.R.; Corsi, S.; Beard, R.; Pradella, D.; Turillazzi, S. Nestmate recognition cues in the honey bee: Differential importance of cuticular alkanes and alkenes. Chem. Senses 2005, 30, 477–489. [Google Scholar] [CrossRef]
- Van Wilgenburg, E.; Felden, A.; Choe, D.H.; Sulc, R.; Luo, J.; Shea, K.J.; Elgar, M.A.; Tsutsui, N.D. Learning and discrimination of cuticular hydrocarbons in a social insect. Biol. Lett. 2012, 8, 17–20. [Google Scholar] [CrossRef]
- Ozaki, M.; Wada-Katsumata, A.; Fujikawa, K.; Iwasaki, M.; Yokohari, F.; Satoji, Y.; Nisimura, T.; Yamaoka, R. Ant nestmate and non-nestmate discrimination by a chemosensory sensillum. Science 2005, 309, 311–314. [Google Scholar] [CrossRef]
- Ozaki, M.; Wada-Katsumata, A. Perception and olfaction of cuticular compounds. In Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Blomquist, G.J., Bagnères, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 207–221. [Google Scholar]
- Espelie, K.E.; Wenzel, J.W.; Chang, G. Surface lipids of social wasp Polistes metricus Say and its nest and nest pedicel and their relation to nestmate recognition. J. Chem. Ecol. 1990, 16, 2229–2241. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Blomquist, G.J.; Millar, J.G.; Hanks, L.M. Role of contact pheromones in mate recognition in Xylotrechus colonus. J. Chem. Ecol. 2003, 29, 533–545. [Google Scholar] [CrossRef]
- Sugeno, W.; Hori, M.; Matsuda, K. Identification of the contact sex pheromone of Gastrophysa atrocyanea (Coleoptera: Chrysomelidae). Appl. Entomol. Zool. 2006, 41, 269–276. [Google Scholar] [CrossRef]
- Lacey, E.S.; Ginzel, M.D.; Millar, J.G.; Hanks, L.M. 7-Methylheptacosane is a major component of the contact sex pheromone of the cerambycid beetle Neoclytus acuminatus acuminatus. Physiol. Entomol. 2008, 33, 209–216. [Google Scholar] [CrossRef]
- Silk, P.J.; Ryall, K.; Barry Lyons, D.; Sweeney, J.; Wu, J. A contact sex pheromone component of the emerald ash borer Agrilus planipennis Fairmaire (Coleoptera: Buprestidae). Naturwissenschaften 2009, 96, 601–608. [Google Scholar] [CrossRef]
- Châline, N.; Sandoz, J.C.; Martin, S.J.; Ratnieks, F.L.W.; Jones, G.R. Learning and discrimination of individual cuticular hydrocarbons by honeybees (Apis mellifera). Chem. Senses 2005, 30, 327–335. [Google Scholar]
- Ginzel, M.D.; Moreira, J.A.; Ray, A.M.; Millar, J.G.; Hanks, L.M. (Z)-9-Nonacosene—Major component of the contact sex pheromone of the beetle Megacyllene caryae. J. Chem. Ecol. 2006, 32, 435–451. [Google Scholar] [CrossRef]
- Espelie, K.E.; Gamboa, G.J.; Grudzien, T.A.; Bura, E.A. Cuticular hydrocarbons of the paper wasp Polistes fuscatus: A search for recognition pheromones. J. Chem. Ecol. 1994, 20, 1677–1687. [Google Scholar] [CrossRef]
- Bos, N.; Dreier, S.; Jørgensen, C.G.; Nielsen, J.; Guerrieri, F.J.; D’Ettorre, P. Learning and perceptual similarity among cuticular hydrocarbons in ants. J. Insect Physiol. 2012, 58, 138–146. [Google Scholar] [CrossRef]
- Ablard, K.; Gries, R.; Khaskin, G.; Schaefer, P.W.; Gries, G. Does the stereochemistry of methylated cuticular hydrocarbons contribute to mate recognition in the egg parasitoid wasp Ooencyrtus kuvanae? J. Chem. Ecol. 2012, 38, 1306–1317. [Google Scholar] [CrossRef]
- Duff, C.M.; Gries, G.; Mori, K.; Shirai, Y.; Seki, M.; Takikawa, H.; Sheng, T.; Slessor, K.N.; Gries, R.; Maier, C.T.; et al. Does pheromone biology of Lambdina athasaria and L. pellucidaria contribute to their reproductive isolation? J. Chem. Ecol. 2001, 27, 431–442. [Google Scholar] [CrossRef]
- Hansen, L.S.; Lövei, G.L.; Székács, A. Survival and development of a stored-product pest, Sitophilus zeamais (Coleoptera: Curculionidae), and its natural enemy, the parasitoid Lariophagus distinguendus (Hymenoptera: Pteromalidae), on transgenic Bt maize. Pest Manag. Sci. 2013, 69, 602–606. [Google Scholar]
- Steidle, J.L.M.; Schöller, M. Olfactory host location and learning in the granary weevil parasitoid Lariophagus distinguendus (Hymenoptera: Pteromalidae). J. Insect Behav. 1997, 10, 331–342. [Google Scholar] [CrossRef]
- Ruther, J.; Homann, M.; Steidle, J.L.M. Female-derived sex pheromone mediates courtship behaviour in the parasitoid Lariophagus distinguendus. Entomol. Exp. Appl. 2000, 96, 265–274. [Google Scholar]
- Benelli, G.; Bonsignori, G.; Stefanini, C.; Dario, P.; Canale, A. Male wing fanning performance during successful and unsuccessful mating in the parasitic wasp Lariophagus distinguendus Förster (Hymenoptera: Pteromalidae). J. Insect Behav. 2013, 26, 228–237. [Google Scholar] [CrossRef]
- Steiner, S.; Steidle, J.L.M.; Ruther, J. Female sex pheromone in immature insect males—A case of pre-emergence chemical mimicry? Behav. Ecol. Sociobiol. 2005, 58, 111–120. [Google Scholar] [CrossRef]
- Steiner, S.; Mumm, R.; Ruther, J. Courtship pheromones in parasitic wasps: Comparison of bioactive and inactive hydrocarbon profiles by multivariate statistical methods. J. Chem. Ecol. 2007, 33, 825–838. [Google Scholar] [CrossRef]
- Van Zweden, J.S.; D’Ettore, P. Nestmate recognition in social insects and the role of cuticular hydrocarbons. In Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Blomquist, G.J., Bagnères, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 222–243. [Google Scholar]
- Wang, S.; Zhang, A. An improved copper-catalyzed cross-coupling reaction of alkyl triflates with primary alkyl Gringnard reagents. Org. Prep. Proced. Int. 2008, 40, 293–301. [Google Scholar] [CrossRef]
- Cahiez, G.; Chaboche, C.; Jézéquel, M. Cu-catalyzed alkylation of Grignard reagents: A new efficient procedure. Tetrahedron 2000, 56, 2733–2737. [Google Scholar] [CrossRef]
- Aizpurua, J.M.; Cossio, F.P.; Palomo, C. Reagents and synthetic methods. 61. Reaction of hindered trialkylsilyl esters and trialkylsilyl ethers with triphenylphosphine dibromide: Preparation of carboxylic acid bromides and alkyl bromides under mild neutral conditions. J. Org. Chem. 1986, 51, 4941–4943. [Google Scholar] [CrossRef]
- Buck, M.; Chong, J.M. Alkylation of 1-alkynes in THF. Tetrahedron Lett. 2001, 42, 5825–5827. [Google Scholar] [CrossRef]
- Zou, Y.; Millar, J.G. Stereoselective synthesis of the obscure mealybug pheromone by hydrogenation of a tetrasubstituted alkene precursor. Tetrahedron Lett. 2011, 52, 4224–4226. [Google Scholar] [CrossRef]
- Sonnet, P.E.; Heath, R.R. Stereospecific synthesis of (Z,Z)-11,13-hexadecadienal, a female sex pheromone of the navel orangeworm, Amyelois transitella (Lepidoptera: Pyralidae). J. Chem. Ecol. 1980, 6, 221–228. [Google Scholar] [CrossRef]
- Barth, S.; Effenberger, F. Lipase-catalyzed resolution of racemic 2-alkyl substituted 1-alkanols. Tetrahedron Asymmetry 1993, 4, 823–833. [Google Scholar] [CrossRef]
- Benoit, D.; Coulbeck, E.; Eames, J.; Motevalli, M. On the structure and chiroptical properties of (S)-4-isopropyl-oxazolidin-2-one. Tetrahedron Asymmetry 2008, 19, 1068–1077. [Google Scholar] [CrossRef]
- Kanomata, N.; Maruyama, S.; Tomono, K.; Anada, S. A simple method removing 2-oxazolidinone and 2-hydroxyethylamine auxiliaries in methoxide-carbonate systems for synthesis of planar-chiral nicotinate. Tetrahedron Lett. 2003, 44, 3599–3603. [Google Scholar]
- Evans, D.A.; Ennis, M.D.; Mathre, D.J. Asymmetric alkylation reactions of chiral imide enolates. A practical approach to the enantioselective synthesis of .alpha.-substituted carboxylic acid derivatives. J. Am. Chem. Soc. 1982, 104, 1737–1739. [Google Scholar] [CrossRef]
- Williams, D.R.; Patnaik, S.; Plummer, S.V. Leucascandrolide A: A second generation formal synthesis. Org. Lett. 2003, 5, 5035–5038. [Google Scholar] [CrossRef]
- Corey, E.J.; Fuchs, P.L. A synthetic method for formyl→ethynyl conversion (RCHO→RC≡CH or RC≡CR′). Tetrahedron Lett. 1972, 13, 3769–3772. [Google Scholar] [CrossRef]
- Armstrong-Chong, R.J.; Matthews, K.; Chong, J.M. Sequential alkynylation of ω-bromoalkyl triflates: Facile access to unsymmetrical non-conjugated diynes including precursors to diene pheromones. Tetrahedron 2004, 60, 10239–10244. [Google Scholar] [CrossRef]
- Bello, J.E.; Millar, J.G. Efficient asymmetric synthesis of long chain methyl-branched hydrocarbons, components of the contact sex pheromone of females of the cerambycid beetle, Neoclytus acuminatus acuminatus. Tetrahedron Asymmetry 2013, 24, 822–826. [Google Scholar] [CrossRef]
- Marukawa, K.; Takikawa, H.; Mori, K. Synthesis of the enantiomers of some methyl-branched cuticular hydrocarbons of the ant Diacamma sp. Biosci. Biotechnol. Biochem. 2001, 65, 305–314. [Google Scholar] [CrossRef]
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Ruther, J.; Steiner, S. Costs of female odour in males of the parasitic wasp Lariophagus distinguendus (Hymenoptera: Pteromalidae). Naturwissenschaften 2008, 95, 547–552. [Google Scholar] [CrossRef]
- Carlson, D.A.; Langley, P.A.; Huyton, P. Sex pheromone of the tsetse fly: Isolation, identification, and synthesis of contact aphrodisiacs. Science 1978, 201, 750–753. [Google Scholar]
- Carlson, D.A.; Offor, I.I.; El Messoussi, S.; Matsuyama, K.; Mori, K.; Jallon, J.M. Sex pheromone of Glossina tachinoides: Isolation, identification, and synthesis. J. Chem. Ecol. 1998, 24, 1563–1575. [Google Scholar] [CrossRef]
- Rutledge, C.E.; Millar, J.G.; Romero, C.M.; Hanks, L.M. Identification of an important component of the contact sex pheromone of Callidiellum rufipenne (Coleoptera: Cerambycidae). Environ. Entomol. 2009, 38, 1267–1275. [Google Scholar] [CrossRef]
- Spikes, A.E.; Paschen, M.A.; Millar, J.G.; Moreira, J.A.; Hamel, P.B.; Schiff, N.M.; Ginzel, M.D. First contact pheromone identified for a longhorned beetle (Coleoptera: Cerambycidae) in the subfamily Prioninae. J. Chem. Ecol. 2010, 36, 943–954. [Google Scholar] [CrossRef]
- Guerrieri, F.J.; Nehring, V.; Jorgensen, C.G.; Nielsen, J.; Galizia, C.G.; d'Ettorre, P. Ants recognize foes and not friends. Proc. R. Soc. B 2009, 276, 2461–2468. [Google Scholar] [CrossRef]
- Steiner, S.; Hermann, N.; Ruther, J. Characterization of a female-produced courtship pheromone in the parasitoid Nasonia vitripennis. J. Chem. Ecol. 2006, 32, 1687–1702. [Google Scholar] [CrossRef]
- Ruther, J.; Döring, M.; Steiner, S. Cuticular hydrocarbons as contact sex pheromone in the parasitoid Dibrachys cavus. Entomol. Exp. Appl. 2011, 140, 59–68. [Google Scholar] [CrossRef]
- Hefetz, A.; Wicker-Thomas, C.; Bagnères, A.G. Future directions in hydrocarbon research. In Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Blomquist, G.J., Bagnères, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 477–485. [Google Scholar]
- Millar, J.G. Chemical synthesis of insect cuticular hydrocarbons. In Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Blomquist, G.J., Bagnères, A.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 163–186. [Google Scholar]
- Ruther, J. Novel insights into pheromone-mediated communication in parasitic hymenopterans. In Chemical Ecology of Insect Parasitoids; Wajnberg, E., Colazza, S., Eds.; Wiley-Blackwell: Chichester, UK, 2013; pp. 112–144. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kühbandner, S.; Bello, J.E.; Mori, K.; Millar, J.G.; Ruther, J. Elucidating Structure-Bioactivity Relationships of Methyl-Branched Alkanes in the Contact Sex Pheromone of the Parasitic Wasp Lariophagus distinguendus. Insects 2013, 4, 743-760. https://doi.org/10.3390/insects4040743
Kühbandner S, Bello JE, Mori K, Millar JG, Ruther J. Elucidating Structure-Bioactivity Relationships of Methyl-Branched Alkanes in the Contact Sex Pheromone of the Parasitic Wasp Lariophagus distinguendus. Insects. 2013; 4(4):743-760. https://doi.org/10.3390/insects4040743
Chicago/Turabian StyleKühbandner, Stephan, Jan E. Bello, Kenji Mori, Jocelyn G. Millar, and Joachim Ruther. 2013. "Elucidating Structure-Bioactivity Relationships of Methyl-Branched Alkanes in the Contact Sex Pheromone of the Parasitic Wasp Lariophagus distinguendus" Insects 4, no. 4: 743-760. https://doi.org/10.3390/insects4040743