Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Larval Collection
2.2. Insect Rearing
2.3. Greenhouse Studies
2.4. Data Analysis
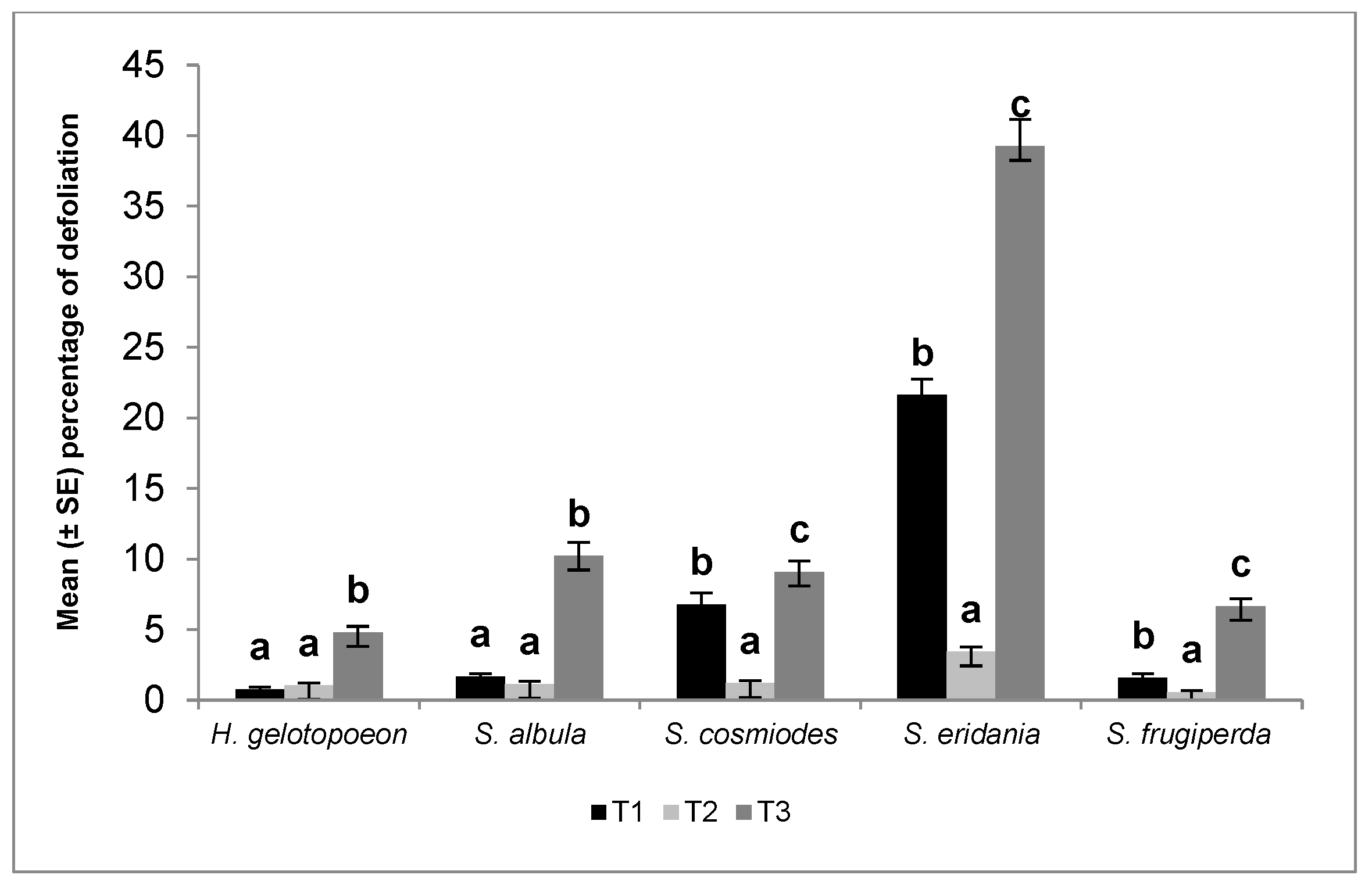
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silva, G.V.; Pasini, A.; Bueno, A.D.F.; Bortolotto, O.C.; Caselato Barbosa, G.; Stopa Cruz, Y.K. No impact of Bt soybean that express Cry1Ac protein on biological traits of Euschistus heros (Hemiptera, Pentatomidae) and its egg parasitoid Telenomus podisi (Hymenoptera, Platygastridae). Rev. Bras. Entomol. 2014, 58, 285–290. [Google Scholar] [CrossRef]
- Argenbio. Gráfico de Evolución de las Superficies Sembradas con OGM en la Argentina, en Porcentajes. Available online: http://www.argenbio.org/adc/uploads/imagenes_doc/planta_stransgenicas/1_Grafico_de_evolucion_de_superficie_cultivada_OGM_en_Arg_en_porcentaje.pdf (accessed on 22 May 2018).
- Bolsa de Cereales. Panorama Agrícola Semanal. Estimaciones Agrícolas. Relevamiento al 29/03/2018. Available online: http://www.bolsadecereales.com.ar (accessed on 22 May 2018).
- Cazado, L.E.; Murúa, M.G.; Casmuz, A.S.; Socías, M.G.; Vera, M.T.; O´Brien, C.W.; Gastaminza, G. Geographical distribution and new associations of Rhyssomatus subtilis (Coleoptera: Curculionidae) in Argentina. Fla. Entomol. 2013, 96, 663–669. [Google Scholar] [CrossRef]
- Casmuz, A.S.; Cazado, L.E.; Scalora, F.S.; Tuzza, M.F.; Fernández, R.A.; Fadda, C.; Fadda, L.A.; Dami, L.; Colledani Toranzo, A.; Gómez, C.H.; et al. Evaluación de diferentes alternativas para el control del complejo de plagas del cultivo de soja. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2013–2014; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 50; EEAOC: San Miguel de Tucumán, Argentina, 2014; pp. 137–142. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/461/10Plagas.html (accessed on 22 May 2018).
- Casmuz, A.S.; Cazado, L.E.; Fadda, L.A.; Tuzza, M.F.; Fadda, C.; Jadur, A.; Merlini, A.; Orce, J.D.; Vera, A.; Marín, M.N.; et al. Momento de aplicación de los insecticidas y su relación con el control ejercido sobre el complejo de orugas defoliadoras de la soja. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2014–2015; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 51, EEAOC: San Miguel de Tucumán, Argentina, 2015; pp. 120–125. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/539/62-Zoologia-621Defoliadoras.html (accessed on 22 May 2018).
- Casmuz, A.S.; Fadda, L.A.; Tuzza, M.F.; Jadur, A.; Fernández, C.; Díaz Arnijas, G.; Vera, M.A.; Cazado, L.E.; Orce, J.D.; Murúa, M.G.; et al. Manejo de plagas en soja Intacta y RR1. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2015–2016; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 52, EEAOC: San Miguel de Tucumán, Argentina, 2016; pp. 134–142. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/645/Capitulo-5.html (accessed on 22 May 2018).
- Casmuz, A.; Fadda, L.; Vera, M.A.; Jadur, A.; Díaz Arnijas, G.; Ramos, L.; Tarulli, L.; Bayos, N.; Fernández Alvarez, C.; Murúa, M.G.; et al. Estrategias para la prevención de la resistencia de insectos en soja Bt: Manejo de plagas en el refugio. In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2016–2017; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 53, EEAOC: San Miguel de Tucumán, Argentina, 2017; pp. 115–121. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/738/Capitulo-5---2017.html (accessed on 22 May 2018).
- Wu, K.; Um, W.; Liang, G.; Gou, Y. Regional reversion of insecticide resistance in Helicoverpa armigera (Lepidoptera: Noctuidae) is associated with the use of Bt cotton in northern China. Pest Manag. Sci. 2005, 61, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Cazado, L.E.; Casmuz, A.S.; Riley, D.G.; Scalora, F.S.; Gastaminza, G.A.; Murúa, M.G. Rhyssomatus subtilis (Coleoptera: Curculionidae) impact in soybean plan stands. J. Entomol. Sci. 2016, 51, 69–78. [Google Scholar] [CrossRef]
- Monsanto. Available online: http://www.intactarr2pro.com.ar/ (accessed on 23 May 2018).
- Lu, Y.; Wu, K.; Jiang, Y.; Xia, B.; Li, P.; Feng, H.; Wyckhuys, K.A.G.; Guo, Y. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China. Science 2010, 328, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- Catangui, M.A.; Berg, R.K. Western bean cutworm, Striacosta albicosta (Smith) (Lepidoptera: Noctuidae), as a potential pest of transgenic Cry1Ab Bacillus thuringiensis corn hybrids in South Dakota. Environ. Entomol. 2006, 35, 1439–1452. [Google Scholar] [CrossRef]
- Hoffmann-Campo, C.B.; Moscardi, F.; Corrêa-Ferreira, B.; Oliveira, L.J.; Sosa-Gómez, D.R.; Panizzi, A.R.; Corso, I.C.; Gazzoni, D.L.; Oliveira, E.B. Pragas da Soja no Brasil e seu Manejo Integrado; Circular Técnica; Embrapa e Centro Nacional de Pesquisa de Soja: Londrina, Brazil, 2000; Volume 30, p. 70. ISSN 1516-7860. [Google Scholar]
- Santos, K.B.; Meneguim, A.M.; Neves, P.M.O.J. Biology and consumption of Spodoptera eridania (Cramer) (Lepidoptera: Noctuidae) in different hosts. Neotrop. Entomol. 2005, 4, 903–910. [Google Scholar] [CrossRef]
- Bueno, R.C.O.F.; Bueno, A.F.; Moscardi, F.; Parra, J.R.P.; Hoffmann-Campo, C.B. Lepidopteran larva consumption of soybean foliage: Basis for developing multiple-species economic thresholds for pest management decisions. Pest Manag. Sci. 2011, 67, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Gómez, D.R.; Correa-Ferreira, B.S.; Hoffmann-Campo, C.B.; Corso, I.C.; Oliveira, L.J.; Moscardi, F.; Panizzi, A.R.; de Freitas Bueno, A.; Hirose, E.; Roggia, S. Manual de Identificação de Insetos e Outros Invertebrados da Cultura de Soja; Embrapa Soja: Londrina, Brazil, 2014; p. 100. ISSN 1516-781X. [Google Scholar]
- Favetti, B.M.; Butnariu, A.R.; Foerster, L.A. Biology and reproductive capacity of Spodoptera eridania (Cramer) (Lepidoptera, Noctuidae) in different soybean cultivars. Rev. Bras. Entomol. 2015, 59, 89–95. [Google Scholar] [CrossRef]
- Cazado, L.E.; Casmuz, A.S.; Scalora, F.S.; Fadda, C.; Fernández, R.A.; Tuzza, M.F.; Fadda, L.A.; Dami, L.; Colledani Toranzo, A.; Jadur, A.; et al. Comportamiento de la soja Bt frente a las principales plagas insectiles y depredadores en el Noroeste Argentino (NOA). In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2013–2014; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial Soja EEAOC Nº 50, EEAOC: San Miguel de Tucumán, Argentina, 2014; pp. 153–161. Available online: http://www.eeaoc.org.ar/publicaciones/categoria/16/463/12Plagas-vs-Bt.html (accessed on 22 May 2018).
- Silva, V.G.; Bueno, F.A.; Bortolottoa, C.O.; Santos, C.A.; Fernandes, P.A. Biological characteristics of black armyworm Spodoptera cosmioides on genetically modified soybean and corn crops that express insecticide Cry proteins. Rev. Bras. Entomol. 2016, 60, 255–259. [Google Scholar] [CrossRef]
- Bentancourt, C.M.; Scatoni, I.B. Lepidópteros de importancia económica. In Reconocimiento, Biología y Daños de las Plagas Agrícolas y Forestales; Hemisferio Sur: Montevideo, Uruguay, 1996; p. 129. ISBN 9974-556-96-6. [Google Scholar]
- Casmuz, A.; Juárez, M.L.; Socías, M.G.; Murúa, M.G.; Prieto, S.; Medina, S.; Willink, E.; Gastaminza, G. Revisión de los hospederos del gusano cogollero del maíz, Spodoptera frugiperda (Lepidoptera: Noctuidae). Rev. Soc. Entomol. Argent. 2010, 69, 209–231. [Google Scholar]
- Nagoshi, R.N.; Meagher, R.L. Behavior and distribution of the two fall armyworm host strains in Florida. Fla. Entomol. 2004, 87, 440–449. [Google Scholar] [CrossRef]
- Murúa, M.G.; Vera, M.T.; Abraham, S.; Juaréz, M.L.; Prieto, S.; Head, G.P.; Willink, E. Fitness and mating compatibility of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) populations from different host plant species and regions in Argentina. Entomol. Soc. Am. 2008, 101, 639–649. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Murúa, M.G.; Hay-Roe, M.; Juárez, M.L.; Willink, E.; Meagher, R.L. Genetic characterization of fall armyworm (Lepidoptera: Noctuidae) host strains in Argentina. J. Econ. Entomol. 2012, 105, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Barros, N.M. Biotic potential and reproductive parameters of Spodoptera eridania (Stoll) (Lepidoptera, Noctuidae) in the laboratory. Rev. Bras. Entomol. 2013, 57, 340–345. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Bortolin, T.M.; Fronza, E.; Pezzi, P.; Luz, P.C.; Barros, N.M. Immature stages of Spodoptera albula (Walker) (Lepidoptera: Noctuidae): Developmental parameters and host plants. An. Acad. Bras. Cienc. 2013, 85, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Barros, N.M. Immature stages of Spodoptera eridania (Lepidoptera: Noctuidae): Developmental parameters and host plants. J. Insect Sci. 2014, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Bortolin, T.M.; Fronza, E.; Pezzi, P.; Luz, P.C.; Barros, N.M. Biotic potential, fertility and life table of Spodoptera albula (Walker) (Lepidoptera: Noctuidae), under controlled conditions. An. Acad. Bras. Cienc. 2014, 86, 723–732. [Google Scholar] [CrossRef]
- Specht, A.; Roque-Specht, V.F. Immature stages of Spodoptera cosmiodes (Lepidoptera: Noctuidae): Developmental parameters and host plants. Zoologia 2016. [Google Scholar] [CrossRef]
- Specht, A.; Montezano, D.G.; Sosa-Gómez, D.R.; Paula-Moraes, S.V.; Roque-Specht, V.F.; Barros, N.M. Reproductive potential of Spodoptera eridania (Stoll) (Lepidoptera: Noctuidae) in the laboratory: Effect of multiple couples and the size. Braz. J. Biol. 2016, 76, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, R.G.; Wan, L.; Knighten, K. Variation in susceptibility of Noctuid (Lepidoptera) larvae attacking cotton and soybean to purified endotoxin proteins and commercial formulations of Bacillus thuringiensis. J. Econ. Entomol. 1999, 92, 21–32. [Google Scholar] [CrossRef]
- Sivasupramaniam, S.; Moar, W.J.; Ruschke, L.G.; Osborn, J.A.; Jiang, C.; Sebaugh, J.L.; Brown, G.R.; Shappley, Z.W.; Oppenhuizen, M.E.; Mullins, J.W.; et al. Toxicity and characterization of cotton expressing Bacillus thuringiensis Cry1Ac and Cry2Ab2 proteins for control of lepidopteran pests. J. Econ. Entomol. 2008, 101, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.A.C.; Santana, G.K.S.; Neiva, M.M.; Moscardi, F.; Martinelli, S.; Sosa Gómez, D.R. Tolerância de Anticarsia gemmatalis Hübner, Pseudoplusia includens (Walker) e Rachiplusia nu (Guenée) à proteína Cry1Ac. In Proceedings of the 6th Congresso Brasileiro de Soja, Cuiabá, Brazil, 11–14 June 2012; pp. 1–5. [Google Scholar]
- Sosa-Gómez, D.; Omoto, C. Resistencia a insecticidas e otros agentes de controle em artrópodes asociados a cultura da soja. In Soja: Manejo Integrado de Insetos e Outros Artópodes-Praga; Hoffmann-Campo, C.B., Correa-Ferreira, B.S., Moscardi, F., Eds.; Embrapa Press: Brasilia, Brazil, 2012; pp. 673–724. ISBN 978-85-7035-139-5. [Google Scholar]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; Dos Santos, A.C.; Omoto, C. Field evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Chandrasena, D.; Signorini, A.; Abratti, G.; Storer, N.P.; Lopez Olaciregui, M.; Alves, A.P.; Pilcher, C.D. Characterization of field-evolved resistance to Bacillus thuringiensis- derived Cry1F δ-endotoxin in Spodoptera frugiperda populations from Argentina. Pest Manag. Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- González, B.J.B. Aspectos Importantes sobre la Evolución y Combate de las Plagas del Algodonero en Colombia. Rev. Peru. Entomol. 1966, 9, 145–155. [Google Scholar]
- Aziz, S.A. Toxicity of certain insecticide standards against the Southern armyworm. J. Econ. Entomol. 1973, 66, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, P.A.; Vasquez, C.H. Prueba de laboratorio con 30 insecticidas en larvas de Prodenia eridania (Cramer) (Lepidoptera: Noctuidae) en alfalfa. Idesia 1974, 3, 133–140. [Google Scholar]
- Gloria, B.R. Control químico del gusano ejercito Prodenia sunia (G.) en tomateira. Rev. Peru. Entomol. 1975, 18, 120–123. [Google Scholar]
- Campos, L.E. Toxicidad de cuatro insecticidas contra Spodoptera frugiperda (J.E. Smith) y Prodenia eridania (Cram.) del Valle de Lluta, Chile. Idesia 1982, 6, 13–19. [Google Scholar]
- Savoie, K.L. Alimentación selectiva por especies de Spodoptera (Lepidoptera: Noctuidae) en un campo de frijol con labranza mínima. Turrialba 1988, 38, 67–70. [Google Scholar]
- Zenner de Polanía, I.; Álvarez Rodriguez, J.A.; Arévalo Maldonado, H.A.; Mejía Cruz, R.; Bayona, M.A. Susceptibilidad de cuatro noctuidos plaga (Lepidoptera) al gene Cry1Ac del Bacillus thuringiensis incorporado al algodonero. Rev. Colomb. Entomol. 2008, 34, 41–50. [Google Scholar]
- Santos-Amaya, O.; Delgado-Restrepo, O.; Argüelles, J.; Aguilera-Garramuño, E. Evaluación del comportamiento del complejo Spodoptera con la introducción de algodón transgénico al Tolima, Colombia. Corpoica cienc. Tecnol. Agropecu. 2009, 10, 24–32. [Google Scholar] [CrossRef]
- Levy, R.; Habeck, D.H. Descriptions of the larvae of Spodoptera sunia and S. latifascia with a key to the mature Spodoptera larvae of the Eastern United States. Ann. Entomol. Soc. Am. 1976, 69, 585–588. [Google Scholar] [CrossRef]
- Passoa, S. Color identification of economically important Spodoptera larvae in Honduras (Lepidoptera: Noctuidae). Insecta Mundi. 1991, 5, 185–195. [Google Scholar]
- Pogue, G.M. A world revision of the genus Spodoptera Guenée (Lepidoptera: Noctuidae). Mem. Am. Entomol. Soc. 2002, 43, 1–202. [Google Scholar]
- Navarro, F.R.; Saini, E.D.; Leiva, P.D. Clave pictórica de polillas de interés agrícola, agrupadas por relación de semejanza; INTA-EEA Pergamino: Buenos Aires, Argentina, 2009; p. 100. ISBN 978-987-521-344-9. [Google Scholar]
- Scalora, F.; Casmuz, A.; Cazado, L.; Socías, G.; Tolosa, G.; Aralde, M.; Guchea, M.A.; Fadda, L.; Gómez, M.; Gómez, H.; et al. Evaluación de diferentes insecticidas para el control de la oruga bolillera, Helicoverpa gelotopoeon Dyar (Lepidoptera: Noctuidae). In El Cultivo de la Soja en el Noroeste Argentino: Campaña 2011/2012; Devani, M.R., Ledesma, F., Sánchez, J.R., Eds.; Publicación Especial EEAOC nº 45. EEAOC: San Miguel de Tucumán, Argentina, 2012; pp. 147–151. Available online: http://www.eeaoc.org.ar/upload/publicaciones/archivos/286/20121122085717000000.pdf (accessed on 25 April 2018).
- Murúa, M.G.; Cazado, L.E.; Casmuz, A.; Herrero, M.I.; Villagrán, M.E.; Vera, A.; Sosa Gómez, D.; Gastaminza, G. Species from the Heliothinae complex (Lepidoptera: Noctuidae) in Tucumán, Argentina, and update of geographical distribution of Helicoverpa armigera. J. Insect Sci. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.I.; Fogliata, S.V.; Vera, A.; Casmuz, A.; Sosa Gómez, D.; Castagnaro, A.P.; Gastaminza, G.; Murúa, M.G. Biological characterization and mating compatibility of Helicoverpa gelotopoeon (D.) (Lepidoptera: Noctuidae) populations from different regions in Argentina. Bull. Entomol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pietrantonio, P.V.; Junek, T.A.; Parker, R.; Mott, D.; Siders, K.; Troxclair, N.; Vargas-Camplis, J.; Westbrook, J.K.; Vassiliou, V.A. Detection and evolution of resistance to the pyrethroid cypermethrin in Helicoverpa zea (Lepidoptera: Noctuidae) populations in Texas. Environ. Entomol. 2007, 36, 1174–1188. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, K.; Gould, F.; Shen, Z. Cry2Ab tolerance response of Helicoverpa armigera (Lepidoptera: Noctuidae) populations from Cry1Ac cotton planting region. J. Econ. Entomol. 2009, 102, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xu, Z.; Zhu, Y.C.; Huang, F.; Wang, Y.; Li, H.; Gao, C.; Zhou, W.; Shen, J. Evidence of field-evolved resistance to Cry1Ac-expressing Bt cotton in Helicoverpa armigera (Lepidoptera: Noctuidae) in northern China. Pest Manag. Sci. 2010, 66, 155–161. [Google Scholar] [PubMed]
- Bird, L.J.; Downes, S.J. Toxicity and cross-resistance of insecticides to Cry2Ab-resistant and Cry2Ab-susceptible Helicoverpa armigera and Helicoverpa punctigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 107, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Brévault, T.; Tabashnik, B.E.; Carrière, Y. A seed mixture increases dominance of resistance to Bt cotton in Helicoverpa zea. Sci. Rep. 2015, 5, 9807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisig, D.D.; Reay-Jones, F.P.F. Inhibition of Helicoverpa zea (Lepidoptera: Noctuidae) growth by transgenic corn expressing Bt toxins and development of resistance to Cry1Ab. Environ. Entomol. 2015, 44, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, O.; Sorgatto, R.J.; Barbosa, A.D.; Domingues, F.A.; Dourado, P.M.; Carvalho, R.A.; Martinelli, S.; Head, G.P.; Omoto, C. Low susceptibility of Spodoptera cosmiodes, Spodoptera eridania and Spodoptera frugiperda (Lepidoptera: Noctuidae) to genetically-modified soybean expressing Cry1Ac protein. Crop Prot. 2014, 58, 33–40. [Google Scholar] [CrossRef]
- Informativo Céleres. Available online: http://www.celeres.com.br/category/analise-de-safra-soja/ (accessed on 23 May 2018).
- Drees, B.M.; Rice, M.E. The vertical beat sheet: A new device for sampling soybeans insects. J. Econ. Entomol. 1985, 78, 1507–1510. [Google Scholar] [CrossRef]
- Velasco de Estacul, M.; Barral, J.M.; Orfila, R.N. Taxonomía, especificidad y caracteres biológicos diferenciados del complejo de especies denominadas “oruga del capullo” del algodón, “oruga de la espiga” del maíz, “oruga del brote” del tabaco y “bolillera” del lino. Rev. Invest. Agropec. Ser. Patol. Veg. 1969, 6, 19–68. [Google Scholar]
- Zenker, M.M.; Specht, A.; Corseuil, E. Estagios imaturos de Spodoptera cosmiodes (Walker), (Lepidoptera: Noctuidae). Rev. Bras. Zool. 2007, 24, 99–107. [Google Scholar] [CrossRef]
- Murúa, M.G.; Virla, E.G.; Defagó, V. Evaluación de cuatro dietas artificiales para la cría de Spodoptera frugiperda (Lep.: Noctuidae) destinada a mantener poblaciones experimentales de himenópteros parasitoides. Bol. San. Veg. Plagas 2003, 29, 43–51. [Google Scholar]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Kogan, M.; Turnipseed, S.G. Soybean growth and assessment of damage by arthropods. In Sampling Methods in Soybean Entomology; Kogan, M., Herzog, D.C., Eds.; Springer Press: New York, NY, USA, 1980; p. 17. ISBN 978-1-4613-8069-6. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 1999; p. 750, ISBN-13 978-0321656865. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; Robledo, C.W. InfoStat, Versión 2008; Grupo InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2008. [Google Scholar]
- Justiniano, W.; Fernandes, M.G.; Tigre Pereira Viana, C.L. Diversity, composition and population dynamics of arthropods in the genetically modified soybeans Roundup Ready® RR1 (GT 40-3-2) and Intacta RR2 PRO® (MON87701 x MON89788). J. Agric. Sci. 2014, 6, 33–44. [Google Scholar] [CrossRef]
- Massoni, F.A.; Schlie, G.; Frana, J.E. Cultivo de soja Bt (RR2 PRO) y convencional (RR1) expuestos a poblaciones naturales de organismos plaga y depredadores. Información Técnica de Cultivos de Verano. Campaña 2013/2014, INTA-Estación Experimental Agropecuaria Rafaela, Argentina. Publ. Misc. 2015, 130, 98–103. Available online: http://www.aapresid.org.ar/wp-content/uploads/2015/01/Soja-Bt-vs-convencional-expuestos-a-organismos-plaga-y-depredadores_MASSONI_F-4.pdf (accessed on 22 May 2018).
- Perotti, E.; Russo, R.; López, R.; Zari, F.; Pradolini, E.; Sanmarti, N.; Maccari, G.; Boero, L.; Gamundi, J. Evaluación de estrategias de manejo de refugios asociados a cultivos de soja Bt sobre plagas blanco, no blanco y sus depredadores. Para Mejorar Producción Cultivos Estivales 2014, 52, 177–182. Available online: http://inta.gob.ar/sites/default/files/script-tmp-inta-evaluacin-estrategias-manejo-refugios-asociados-.pdf (accessed on 22 May 2018).
- Forti, L.A.; Sosa-Gómez, D.R. Suscetibilidade relativa de Spodoptera frugiperda, S. eridania, S. albula, S. cosmiodes e Anticarsia gemmatalis a os inseticidas chlorantraniliprole e flubendiamida. In Proceedings of the 9th Jornada Acadêmica da Embrapa Soja, Londrina, Brazil, 12 June 2014; pp. 126–129. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/105824/1/JORN.2014.pdf (accessed on 22 May 2018).
- Pastrana, J.A. Los Lepidópteros Argentinos, Sus Plantas Hospedadoras y Otros Sustratos Alimenticios; Sociedad Entomológica Argentina: Buenos Aires, Argentina, 2004; p. 334. ISBN 987-21319-0-2. [Google Scholar]
- Sparks, A. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Silva, M.D.; Bueno, F.A.; Stecca, S.C.; Andrade, K.; Neves, J.O.M.P.; Oliveira, N.M.C. Biology of Spodoptera eridania and Spodoptera cosmioides (Lepidoptera: Noctuidae) on different host plants. Fla. Entomol. 2017, 100, 752–760. [Google Scholar] [CrossRef]
- Angulo, A.O.; Olivares, T.S.; Weigert, G.T.H. Estados Inmaduros de Lepidópteros Noctuídos de Importancia Agrícola y Forestal en Chile y Claves Para su Identificación (Lepidoptera: Noctuidae); Universidad de Concepción: Concepción, Chile, 2008; p. 114. [Google Scholar]


{kind=link}
| Study | Species * | Identity of Species ** | Environmental Conditions (Controlled or Field Conditions) | Type of Infestation | Crop or Artificial Diet Used | Evaluations | Impacts |
|---|---|---|---|---|---|---|---|
| Present study | Sf, Se, Sa, Sc, Hg | individual | controlled (greenhouse) | artificial | soybean (Bt and non-Bt) | Performance and potencial risk | Bt soybean had the best control of Hg and a suppressive effect on Sf and Sa. Se and Sc were not susceptible to the Bt soybean. Se represents a potential risk in soybean crops |
| [60] | Sc, Se, Sf | individual | controlled | artificial | soybean (Bt and non-Bt) and diet | Susceptibility to Cry1Ac protein | Bt soybean showed poor control of Sc, Se, Sf |
| [16] | Sf, Se, Sc | individual | controlled | artificial | soybean genotypes | Larva consumption foliage | Sc defoliated nearly twice the leaf area of Sa and Sf |
| [5] | S. spp. | general | field conditions | natural | soybean | Different chemical alternatives for the management of the complex pest | Diamide + neonicotinoid had the best control |
| [6] | Sc | individual | field conditions | natural | soybean | Moment of application of different insecticides | Early application of diamide delayed the damage of larvae |
| [7] | H. spp., S. spp. | general | field conditions | natural | soybean (Bt and non-Bt) | Pest management | Early application of diamide delayed the damage of larvae |
| [8] | H. spp. | general | field conditions | natural | soybean (Bt and non-Bt) | Strategies for prevention of insect resistance | Low presence of H. spp. in Bt soybean and refuge |
| [19] | S. spp., H. spp. | general | field conditions | natural | soybean (Bt and non-Bt) | Behavior of Bt soybean on the pests and its predators | Bt soybean provided a good control of S. spp. and H. spp. and did not affect its predators |
| [18] | Se | individual | controlled | artificial | soybean cultivars (non-Bt) | Development, survival, and reproductive capacity | The development of Se was affected by the cultivar |
| [73] | Sf, Se, Sa, Sc | individual | controlled | artificial | diet | Susceptibility to chlorantraniliprole and flubendiamide | Chlorantraniliprole showed a higher mortality than flubendiamide for all Lepidoptera species tested |
| [70] | S. spp., Sf | general | field conditions | natural | soybean (Bt and non-Bt) | Diversity, composition, and population dynamics | Bt soybean reduced the target insect pests and favored populations of natural enemies |
| [71] | Sf, Sc, Hg | individual | field conditions | natural | soybean (Bt and non-Bt) | Insect abundance | Densities of the species were low in both treatment |
| [27] | Sa | controlled | artificial | diet | Developmental parameters and host plants | Complete detail of biological parameters of Sa and 55 host plant species of Sa are listed | |
| [26] | Se | individual | controlled | artificial | diet | Biotic potential and reproductive parameters | Complete detail of reproductive and population parameters of Se |
| [28] | Se | individual | controlled | artificial | diet | Developmental parameters and host plants | Complete detail of biological parameters of Se and 202 host plant species of Se are listed |
| [29] | Sa | controlled | artificial | diet | Biotic potential, life table parameters and fertility | Complete detail of reproductive and population parameters of Sa | |
| [72] | Sf, Sc, Hg | individual | field conditions | natural | soybean (Bt and non-Bt) | Strategies of refuge management | The management of refuges with selective insecticides and high persistence allowed to reduce the number of applications and to achieve greater survival of predators and adults of target pests |
| [15] | Se | individual | controlled | artificial | cotton, soybean, and Ipomoea grandifolia (L.) (Convolvulaceae) | Biology on different host plant | Soybean was the least suitable for the development of Se, and I. grandifolia was shown a suitable alternate host for Se |
| [20] | Sc | individual | controlled | artificial | corn (Bt and non-Bt), soybean (Bt and non-Bt), and diet | Development and reproduction | Bt and non-Bt corn adversely affect the development of Sc, and Bt soybean did not affect its biology, suggesting that Sc has major potential to become an important pest in Bt soybean crops |
| [76] | Se, Sc | individual | controlled | artificial | soybean, cotton, corn, Triticum aestivum (L.) (Poaceae), Avena sativa (L.), and diet | Biology on different host plants | Soybean and cotton were more suitable hosts for the development of Se and Sc |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murúa, M.G.; Vera, M.A.; Herrero, M.I.; Fogliata, S.V.; Michel, A. Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests. Insects 2018, 9, 93. https://doi.org/10.3390/insects9030093
Murúa MG, Vera MA, Herrero MI, Fogliata SV, Michel A. Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests. Insects. 2018; 9(3):93. https://doi.org/10.3390/insects9030093
Chicago/Turabian StyleMurúa, María G., Martín A. Vera, María I. Herrero, Sofia V. Fogliata, and Andrew Michel. 2018. "Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests" Insects 9, no. 3: 93. https://doi.org/10.3390/insects9030093
APA StyleMurúa, M. G., Vera, M. A., Herrero, M. I., Fogliata, S. V., & Michel, A. (2018). Defoliation of Soybean Expressing Cry1Ac by Lepidopteran Pests. Insects, 9(3), 93. https://doi.org/10.3390/insects9030093