Climatic Niche Model for Overwintering Monarch Butterflies in a Topographically Complex Region of California

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Monarch Butterfly Overwintering, Population Decline, and Climate
1.2. Attributes of Occupied Monarch Butterfly Overwintering Groves
1.3. Santa Barbara County Is a Powerful Case Study
2. Materials and Methods
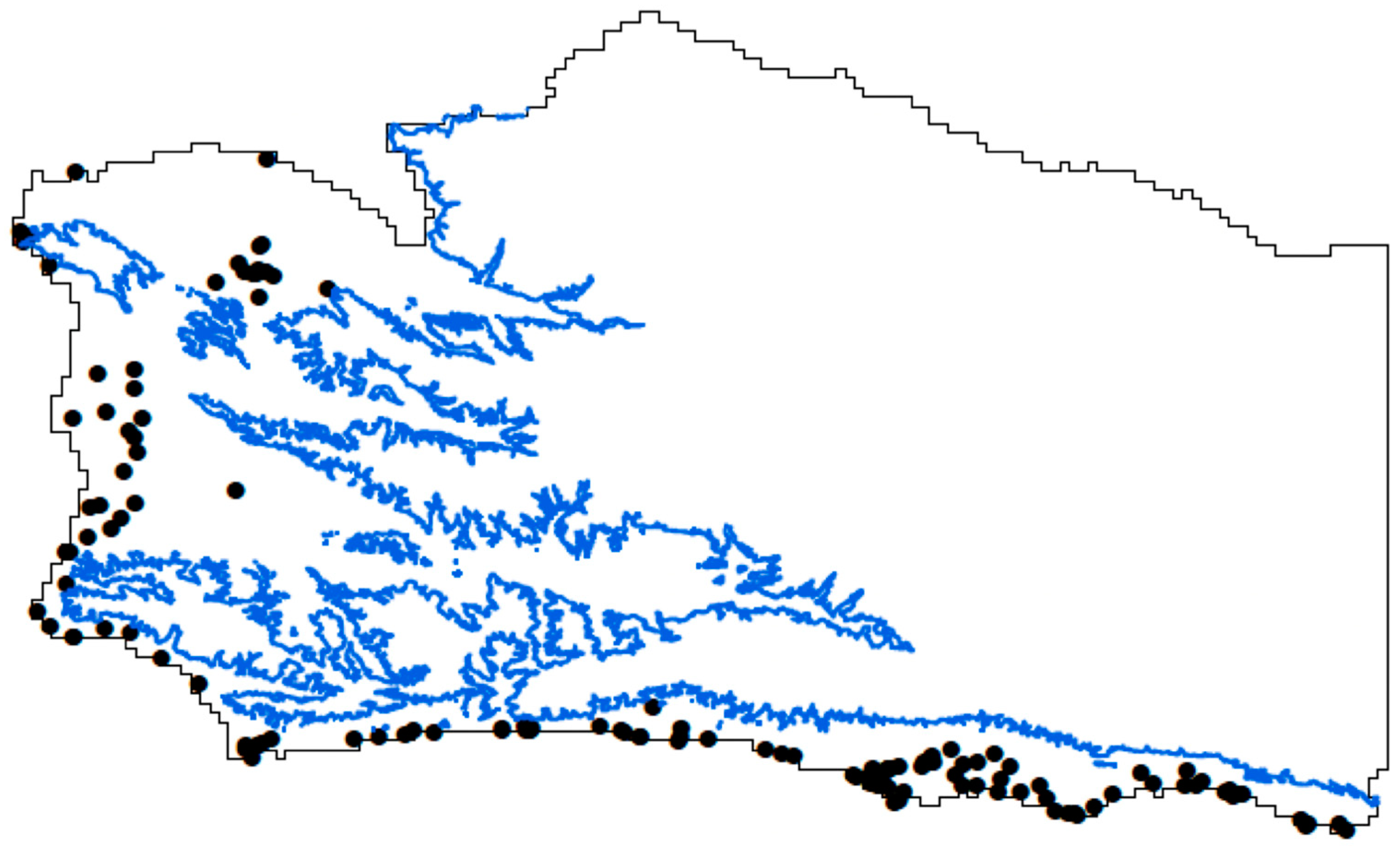
2.1. Distribution Data and Models for Species That Aggregate
2.2. Species Distribution Models
2.3. Attribute Layers in Present-Day Analysis
2.4. Attribute Layers in 2050 Analysis
2.5. Controlling Model Complexity
2.6. Optimal Regularization Coefficient for the Base Model
2.7. Eliminating Correlated Variables
2.8. Final Models
3. Results
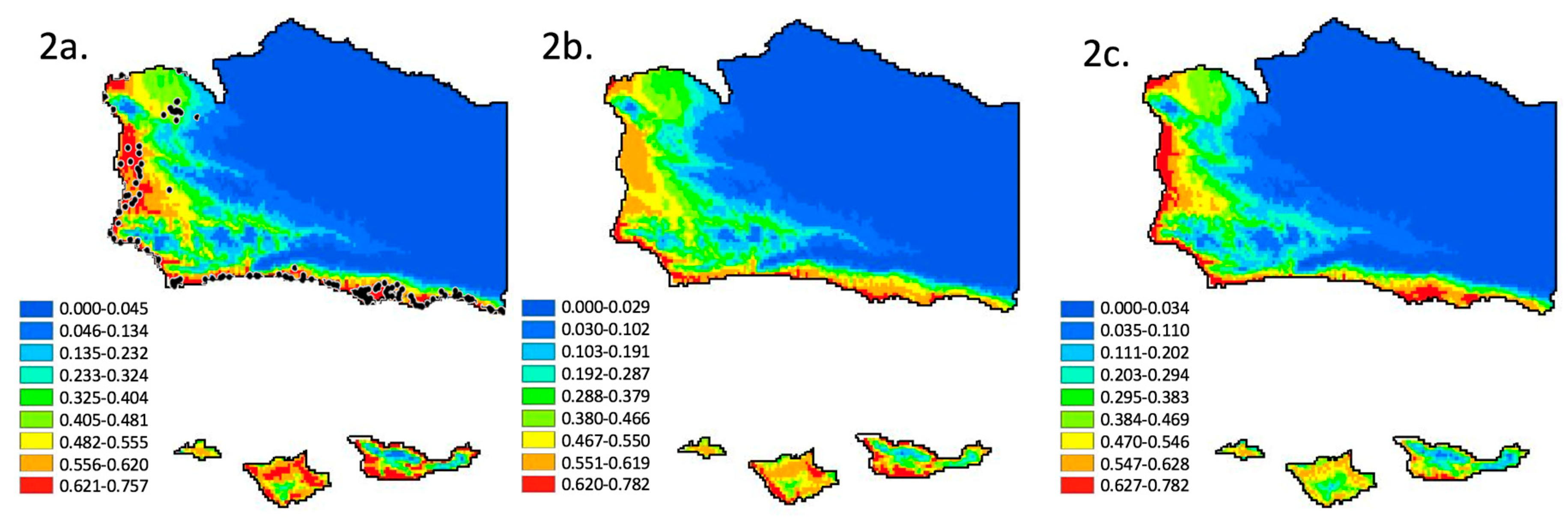
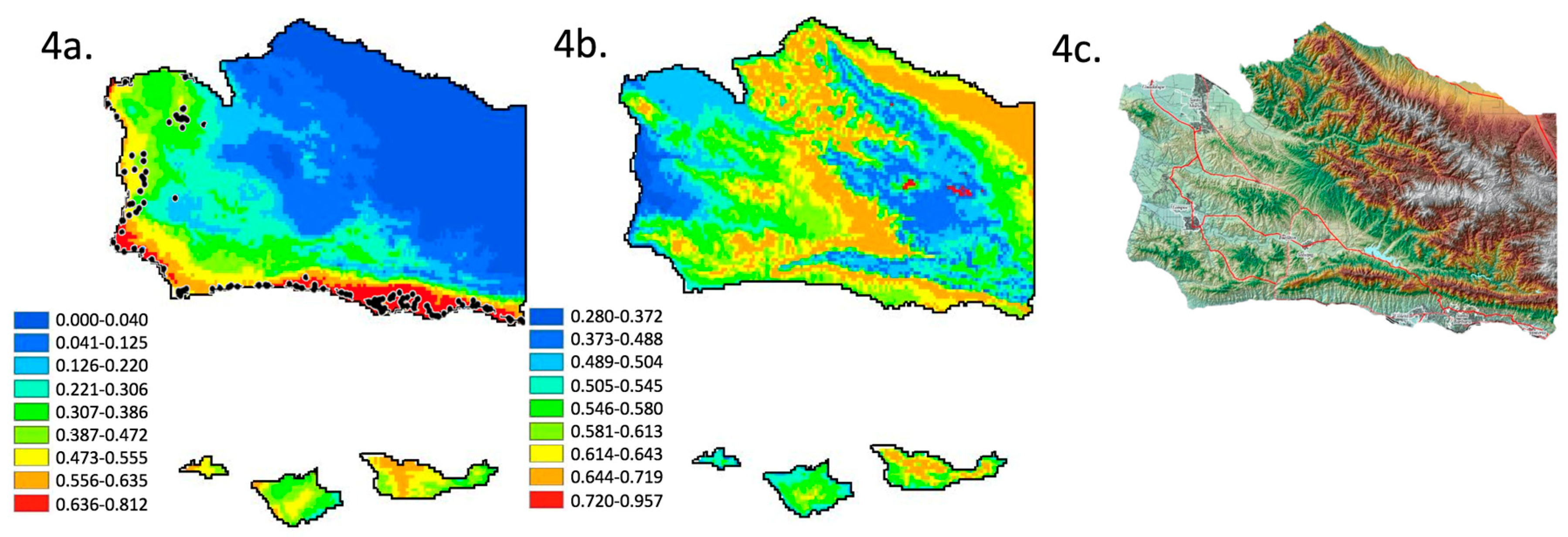
3.1. Species Distribution Model—Climate Variables and Current Distribution
3.2. Species Distribution Model—Moderate Climate Change Scenario
3.3. Species Distribution Model—Extreme Climate Change Scenario
3.4. Optimizing Beta, Removing Auto Correlated and Low Importance Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pelton, E.; Jepsen, S.; Schultz, C.; Fallon, C.; Black, S.H. State of the Monarch Butterfly Overwintering Sites in California; Xerces Society for Invertebrate Conservation: Portland, OR, USA, 2016. [Google Scholar]
- Thogmartin, W.E.; Wiederholt, R.; Drum, R.G.; Diffendorfer, J.E.; Altizer, S.; Taylor, O.R.; Pleasants, J.; Semmens, D.; Semmens, B.; Erickson, R.; et al. Monarch butterfly population decline in North America: Identifying the threatening processes. R. Soc. Open Sci. 2017, 4, 170760. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.B.; Brown, L.M.; Pelton, E.; Crone, E.E. Citizen science monitoring demonstrates dramatic declines of monarch butterflies in western North America. Biol. Conserv. 2017, 214, 343–346. [Google Scholar] [CrossRef]
- Flockhart, D.T.T.; Pichancourt, J.B.; Norris, D.R.; Martin, T.G. Unravelling the annual cycle in a migratory animal: Breeding-season habitat loss drives population declines of monarch butterflies. J. Anim. Ecol. 2015, 84, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Urquhart, F.A.; Urquhart, N.R. Autumnal migration routes of the eastern population of the monarch butterfly (Danaus p. plexippus L.; Danaidae; Lepidoptera) in North America to the overwintering site in the Neovolcanic Plateau of Mexico. Can. J. Zool. 1978, 56, 1759–1764. [Google Scholar] [CrossRef]
- Brower, L.P. Understanding and misunderstanding the migra non of the monarch butterfly (Nymphalidae) in North America: 1857-1995. J. Lepidopterists’ Soc. 1995, 49, 304–385. [Google Scholar]
- Reppert, S.M.; de Roode, J.C. Demystifying monarch butterfly migration. Curr. Biol. 2018, 28, R1009–R1022. [Google Scholar] [CrossRef] [PubMed]
- Lascelles, B.; Notarbartolo Di Sciara, G.; Agardy, T.; Cuttelod, A.; Eckert, S.; Glowka, L.; Hoyt, E.; Llewellyn, F.; Louzao, M.; Ridoux, V.; et al. Migratory marine species: Their status, threats and conservation management needs. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 111–127. [Google Scholar] [CrossRef]
- Brower, L.P. New perspectives on the migration biology of the monarch butterfly, Danaus plexippus L. Pac. Discovery 1985, 27, 748–785. [Google Scholar]
- Snook, L.C. Conservation of the monarch butterfly reserves in Mexico: Focus on the forest. Biol. Conserv. Monarch Butterfly 1993, 38, 362. [Google Scholar]
- Tuskes, P.M.; Brower, L.P. Overwintering ecology of the monarch butterfly, Danaus plexippus L., in California. Ecol. Entomol. 1978, 3, 141–153. [Google Scholar] [CrossRef]
- Chaplin, S.B.; Wells, P.H. Energy reserves and metabolic expenditures of monarch butterflies overwintering in southern California. Ecol. Entomol. 1982, 7, 249–256. [Google Scholar] [CrossRef]
- Nagano Nagano, C.D.; Sakai, W.H.; Malcolm, S.B.; Cockrell, B.J.; Donahue, J.P.; Brower, L.P. Spring migration of monarch butterflies in California. In Biology and Conservation of the Monarch Butterfly; Natural Histo: Los Angeles, CA, USA, 1993. [Google Scholar]
- Griffiths, J.; Villablanca, F. Managing monarch butterfly overwintering groves: Making room among the eucalyptus. Calif. Fish Game 2015, 101, 40–50. [Google Scholar]
- Pierce, A.A.; Zalucki, M.P.; Bangura, M.; Udawatta, M.; Kronforst, M.R.; Altizer, S.; Haeger, J.F.; de Roode, J.C. Serial founder effects and genetic differentiation during worldwide range expansion of monarch butterflies. Proc. R. Soc. B Biol. Sci. 2014, 281, 20142230. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Lennon, J.J. Birds extend their ranges northwards. Nature 1999, 399, 213. [Google Scholar] [CrossRef]
- Warren, M.S.; Hill, J.K.; Thomas, J.A.; Asher, J.; Fox, R.; Huntley, B. Rapid responses of British butteries to opposing forces of climate and habitat change. Nature 2001, 414, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.L.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Pollard, E.; Greatorex-Davies, J.N.; Thomas, J.A. Drought reduces breeding success of the butterfly Aglais urticae. Ecol. Entomol. 1997, 22, 315–318. [Google Scholar] [CrossRef]
- Parmesan, C. Poleward shifts in geographic ranges of butternfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Hill, J.K.; Thomas, C.D.; Fox, R.; Telfer, M.G.; Willis, S.G.; Asher, J.; Huntley, B. Responses of butterflies to twentieth century climate warming: Implications for future ranges. Proc. R. Soc. B Biol. Sci. 2002, 269, 2163–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, P.R.; Hanski, I. On the Wings of the Checkerspots: A Model System for Population Biology; 2004; Oxford University Press: Oxford, UK,. [Google Scholar]
- Sparks, T.H.; Huber, K.; Dennis, R.L.H. Complex phenological responses to climate warming trends? Lessons from history. Eur. J. Entomol. 2006, 103, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Powney, G.D.; Roy, D.B.; Chapman, D.; Oliver, T.H. Synchrony of butterfly populations across species’ geographic ranges. Oikos 2010, 119, 1690–1696. [Google Scholar] [CrossRef]
- Srygley, R.B.; Dudley, R.; Oliveira, E.G.; Aizprúa, R.; Pelaez, N.Z.; Riveros, A.J. El Niño and dry season rainfall influence hostplant phenology and an annual butterfly migration from Neotropical wet to dry forests. Glob. Chang. Biol. 2010, 16, 936–945. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, N.P. Climate change may alter breeding ground distributions of eastern migratory monarchs (Danaus plexippus) via range expansion of Asclepias host plants. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Sakai, W.H.; Calvert, W.C. Department of Parks and Recreation Statewide Monarch Butterfly Management Plan for the State of California: Final Report; California Department of Parks and Recreation: Sacramento, CA, USA, 1991. [Google Scholar]
- Leong, K.L.H. Microenvironmental factors associated with the winter habitat of the monarch butterfly (Lepidoptera: Danaidae ) in Central California. Ann. Entomol. Soc. Am. 1990, 83, 906–910. [Google Scholar] [CrossRef]
- Rich, P.M.; Weiss, S.B. Spatial models of microclimate and habitat suitability: Lessons from threatened species. In Proceedings of the Eleventh Annual ESRI User Conference, Palm Springs, CA, USA, 20–24 May 1991; pp. 95–99. [Google Scholar]
- Leong, K.L.; Sakai, W.H.; Bremer, W.; Feuerstein, D.; Yoshimura, G. Analysis of the pattern of distribution and abundance of monarch overwintering sites along the California coastline. In The Monarch Butterfly: Biology and Conservation; Cornell University Press: Ithaca, NY, USA, 2004; pp. 177–185. [Google Scholar]
- Leong, K.L.H. Evaluation and management of California monarch winter sites. Western Wildl. 2016, 3, 19–26. [Google Scholar]
- Leong, K.L.H.; Frey, D.; Brenner, G.; Baker, S.; Fox, D. Use of multivariate analyses to characterize the monarch butterfly (Lepidoptera: Danaidae) winter habitat. Ann. Entomol. Soc. Am. 1991, 84, 263–267. [Google Scholar] [CrossRef]
- Badik, K.J.; Shapiro, A.M.; Bonilla, M.M.; Jahner, J.P.; Harrison, J.G.; Forister, M.L. Beyond annual and seasonal averages: Using temporal patterns of precipitation to predict butterfly richness across an elevational gradient. Ecol. Entomol. 2015, 40, 585–595. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Mysterud, A. Weather packages: Finding the right scale and composition of climate in ecology. J. Anim. Ecol. 2005, 74, 1195–1198. [Google Scholar] [CrossRef]
- Warren, D.L.; Wright, A.N.; Seifert, S.N.; Shaffer, H.B. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern. Divers. Distrib. 2014, 20, 334–343. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.L.; Seifert, S. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Soc. Am. 2011, 21, 335–342. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Searcy, C.A.; Shaffer, H.B. Do ecological niche models accurately identify climatic determinants of species ranges? Am. Nat. 2016, 187, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Sakai, W.; Nagano, C.; Evans, A.V.; Schrumpf, J.; Lane, J.; Monroe, M. The Wintering Colonies of the Monarch Butterfly (Danaus plexippus) in the State of California, USA; Final Report; California Department of Fish and Game: Sacramento, CA, USA, 1989. [Google Scholar]
- Meade, D.E. Monarch Butterfly Overwintering Sites in Santa Barbara County, California; Althouse and Meade Inc.: Paso Robles, CA, USA, 1999. [Google Scholar]
- Daniel, E.; Griffiths, J.; van der Heide, C.; Villablanca, F.; Meade, D.E.; Griffiths, J.; van der Heide, C.; Villablanca, F. Monarch Butterfly Overwintering Sites, Santa Barbara County, Santa Barbara; Althouse and Meade, Inc.: Paso Robles, CA, USA, 2018. [Google Scholar]
- Phillips, S.B.; Aneja, V.P.; Kang, D.; Arya, S.P. Modelling and analysis of the atmospheric nitrogen deposition in North Carolina. Int. J. Glob. Environ. Issues 2006, 6, 231–252. [Google Scholar] [CrossRef]
- Dormann, C.F.; McPherson, J.M.; Araújo, M.B.; Bivand, R.; Bolliger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Kissling, W.D.; Kühn, I. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.F.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Jobst, A.M.; Kingston, D.G.; Cullen, N.J.; Sirguey, P. Combining thin-plate spline interpolation with a lapse rate model to produce daily air temperature estimates in a data-sparse alpine catchment. Int. J. Climatol. 2017, 37, 214–229. [Google Scholar] [CrossRef]
- Hayhoe, K.; Cayan, D.; Field, C.B.; Frumhoff, P.C.; Maurer, E.P.; Miller, N.L.; Moser, S.C.; Schneider, S.H.; Cahill, K.N.; Cleland, E.E.; et al. Emissions pathways, climate change, and impacts on California. Proc. Natl. Acad. Sci. USA 2004, 101, 12422–12427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cayan, D.R.; Maurer, E.P.; Dettinger, M.D.; Tyree, M.; Hayhoe, K. Climate change scenarios for the California region. Clim. Chang. 2007, 87. [Google Scholar] [CrossRef]
- Kissling, W.D.; Ahumada, J.A.; Bowser, A.; Fernandez, M.; Fernández, N.; García, E.A.; Guralnick, R.P.; Isaac, N.J.B.; Kelling, S.; Los, W.; et al. Building essential biodiversity variables (EBVs) of species distribution and abundance at a global scale. Biol. Rev. 2018, 93, 600–625. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.A.; Inamine, H. Mechanisms behind the monarch’s decline. Science 2018, 360, 1294–1296. [Google Scholar] [CrossRef] [PubMed]
- Valle, D.; Albuquerque, P.; Zhao, Q.; Barberan, A.; Fletcher, R.J. Extending the Latent Dirichlet Allocation model to presence/absence data: A case study on North American breeding birds and biogeographic shifts expected from climate change. Glob. Chang. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dingle, H.; Zalucki, M.; Rochester, W.; Armijo-Prewitt, T. Distribution of the monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Nymphalidae), in western North America. Biol. J. Linnean Soc. 2005, 85, 491–500. [Google Scholar] [CrossRef]
- Pyle, R. Chasing Monarchs: Migrating with Butterflies of Passage; Yale University Press: New Haven, CT, USA, 2014. [Google Scholar]
- Wenner, A.M.; Harris, A.M. Do California Monarchs Undergo Long-Distance Directed Migration? In Biology and Conservation of the Monarch Butterfly; Los Angeles County Museum of Natural History: Los Angeles, CA, USA, 1993; pp. 209–218. [Google Scholar]
- Davis, A.J.; Jenkinson, L.S.; Lawton, J.H.; Shorrocks, B.; Wood, S. Making mistakes when predicting shifts in species range in response to global warming. Nature 1998, 391, 783–786. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fisher, A.; Saniee, K.; Van der Heide, C.; Griffiths, J.; Meade, D.; Villablanca, F. Climatic Niche Model for Overwintering Monarch Butterflies in a Topographically Complex Region of California. Insects 2018, 9, 167. https://doi.org/10.3390/insects9040167
Fisher A, Saniee K, Van der Heide C, Griffiths J, Meade D, Villablanca F. Climatic Niche Model for Overwintering Monarch Butterflies in a Topographically Complex Region of California. Insects. 2018; 9(4):167. https://doi.org/10.3390/insects9040167
Chicago/Turabian StyleFisher, Ashley, Kiana Saniee, Charis Van der Heide, Jessica Griffiths, Daniel Meade, and Francis Villablanca. 2018. "Climatic Niche Model for Overwintering Monarch Butterflies in a Topographically Complex Region of California" Insects 9, no. 4: 167. https://doi.org/10.3390/insects9040167