Field and Laboratory Studies on the Ecology, Reproduction, and Adult Diapause of the Asian Comma Butterfly, Polygonia c-aureum L. (Lepidoptera: Nymphalidae)


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Rearing Methods
2.3. Field Examination
2.4. Experimental Protocols
2.4.1. Measurement of Forewing Length
2.4.2. Diapause Status
2.4.3. Testis Size
2.4.4. Development of Accessory Glands and Simplex
2.4.5. Spermatophore
2.4.6. Mating Experiments
2.4.7. Spermatogenesis
2.5. Phenology of Host Plant
2.6. Statistical Analysis
3. Results
3.1. Forewing Length
3.2. Diapause
3.3. Spermatophore and Mating in Females
3.4. Male Spermatophore Production Ability
3.5. Development of Male Reproductive Organs
3.6. Spermatogenesis
3.7. Sperm Movement
3.8. Population Dynamics
3.9. Phenology of the Host Plant
4. Discussion
4.1. Spermatogenesis
4.2. Development of Accessory Glands and Simplex
4.3. Mating
4.4. Spermatophore
4.5. Oogenesis and Mating Status in Females
4.6. Changes of Color or Size of Male Reproductive Organs
4.7. Estimation of Male Mating Status
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saunders, D.S. Photoperiodism in insects: Migration and diapause responses. In Photoperiodism: The Biological Calendar; Nelson, R.J., Denlinger, D.L., Somers, D.E., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 218–257. [Google Scholar]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptation of Insects; Oxford University Press: Oxford, UK, 1986. [Google Scholar]
- Danks, H.V. Insect Dormancy: An Ecological Perspective. Ottawa, Canada Biological Survey of Canada Monograph Series No. 1; National Museum of Natural Sciences: Ottawa, ON, Canada, 1987. [Google Scholar]
- Evenden, M.L.; Armitage, G.; Lau, R. Effects of nutrition and methoprene treatment upon reproductive diapause in Caloptilia fraxinella (Lepidoptera: Gracillariidae). Physiol. Entomol. 2007, 32, 275–282. [Google Scholar] [CrossRef]
- Pener, M.P. Environmental cues, endocrine factors, and reproductive diapause in male insects. Chronobiol. Int. 1992, 9, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.; Benz, G. Control of spermatogenesis resumption in post-diapausing larvae of the codling moth. J. Insect Physiol. 1982, 28, 349–355. [Google Scholar] [CrossRef]
- Shimoda, M.; Kubo-Irie, M.; Ohta, K.; Irie, M.; Mohri, H. Spermatogenesis in the testes of diapause and non-diapause pupae of the sweet potato hornworm, Agrius convolvuli (L.) (Lepidoptera: Sphingidae). Zool. Sci. 2007, 24, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Kubo-Irie, M.; Yamaguchi, T.; Tanaka, Y.; Yamazaki, I.; Irie, M.; Mohri, H.; Shimoda, M. Identification of the starting point for spermatogenesis resumption in the post-diapause development of the sweet potato hornworm, Agrius convolvuli L. J. Insect Physiol. 2011, 57, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Santa, H.; Otuka, M. Studies on the diapause in the cabbage armyworm, Barathra brassicae L.IV. Development of the male sex cells under the condition inducing diapause or non-diapause. Bull. Nat. Inst. Agric. Sci. Ser. C 1955, 5, 57–65. [Google Scholar]
- Reissig, W.H.; Kamm, J.A. Reproductive development of adult male Draeculacephala crassicornis. Ann. Entomol. Soc. Am. 1975, 68, 58–60. [Google Scholar] [CrossRef]
- Sims, S.R. Aspects of mating frequency and reproductive maturity in Papilio zelicaon. Am. Midl. Nat. 1979, 102, 36–50. [Google Scholar] [CrossRef]
- Hill, H.F., Jr.; Wenner, A.M.; Wells, P.H. Reproductive behavior in an overwintering aggregation of monarch butterflies. Am. Midl. Nat. 1976, 95, 10–19. [Google Scholar] [CrossRef]
- Thibout, E. Evolution and role of apyrene sperm cells of lepidopterans: Their activation and denaturation in the leek moth, Acrolepiopsis assectella (Hypomeutoïdae). In Advances in Invertebrate Reproduction; Clark, W.H., Adams, T.S., Eds.; Elsevier North Holland, Inc.: Amsterdam, The Netherlands, 1980; pp. 231–242. [Google Scholar]
- Mancini, K.; Dolder, H. Ultrastructure of apyrene and eupyrene spermatozoa from the seminal vesicle of Euptoieta hegesia (Lepidoptera: Nymphalidae). Tissue Cell 2001, 33, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Fernadez, F.C.; Cruz-Landim, C. Apyrene spermatozoa in Achroia grisella (Lepidoptera: Piralidae): A honeybee waxmoth. Braz. J. Biol. 2005, 65, 739–741. [Google Scholar] [CrossRef]
- Dallai, R.; Gottardo, M.; Beutel, R.G. Structure and evolution of insect sperm: New interpretations in the age of phylogenomics. Annu. Rev. Entomol. 2016, 61, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sonnenschein, M.; Häuser, C.L. Presence of only eupyrene spermatozoa in adult males of the genus Micropterix hubner and its phylogenetic significance (Lepidoptera: Zeugloptera, Micropterigidae). Int. J. Insect Morphol. Embryol. 1990, 19, 269–276. [Google Scholar] [CrossRef]
- Hamon, C.; Chauvin, G. Ultrastructural analysis of spermatozoa of Korscheltellus lupulinus L. (Lepidoptera: Hepialidae) and Micropterix calthella L. (Lepidoptera: Mircopterigidae). Int. J. Morphol. Embryol. 1992, 21, 149–160. [Google Scholar] [CrossRef]
- Curril, I.M.; LaMunyon, C.W. Sperm storage and arrangement within females of the arctiid moth Utetheisa ornatrix. J. Insect Physiol. 2006, 52, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Silberglied, R.E.; Shepherd, J.G.; Dickinson, J.L. Eunuchs: The role of apyrene sperm in Lepidoptera? Am. Nat. 1984, 123, 255–265. [Google Scholar] [CrossRef]
- Osanai, M.; Kasuga, H.; Aigaki, T. Physiological role of apyrene spermatozoa of Bomvyx mori. Experientia 1987, 43, 593–596. [Google Scholar] [CrossRef]
- Snook, R.R. Is the production of multiple sperm types adaptive? Evolution 1997, 51, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, M.; Delisle, J.; McNeil, J.N. Effects of different male remating intervals on the reproductive success of Choristoneura rosaceana males and females. J. Insect Physiol. 2007, 53, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Tschudi-Rein, K.; Benz, G. The bipotentiality of lepidopteran primary spermatocytes questioned. Mitteil. Schweiz. Entomol. Gesel. Bull. Soc. Entomol. Suisse 1990, 63, 81–85. [Google Scholar]
- Cook, P.A.; Wedell, N. Non-fertile sperm delay female remating. Nature 1999, 397, 486. [Google Scholar] [CrossRef]
- Konagaya, T.; Idogawa, N.; Watanabe, M. Motility and dynamics of eupyrene and apyrene sperm in the spermatheca of the monandrous swallowtail butterfly Byasa alcinous. Physiol. Entomol. 2016, 41, 185–192. [Google Scholar] [CrossRef]
- Konagaya, T.; Watanabe, M. Adaptive significance of the mating of autumn-morph females with non-overwintering summer-morph males in the Japanese common grass yellow, Eurema mandarina (Lepidoptera: Pieridae). Appl. Entomol. Zool. 2015, 50, 41–47. [Google Scholar] [CrossRef]
- Sasaki, N.; Watanabe, M. Alternative ejaculate allocation tactics in relation to male mating history of the swallowtail butterfly, Papilio xuthus L. (Lepidoptera: Papilionidae). J. Res. Lepid. 2015, 48, 1–8. [Google Scholar]
- Cardoso, M.Z.; Silva, E.S. Spermatophore quality and production in two Heliconius butterflies with contrasting mating systems. J. Insect Behav. 2015, 28, 693–703. [Google Scholar] [CrossRef]
- LaMunyon, C.W.; Huffman, T.S. Determinants of sperm transfer by males of the noctuid moth Heliothis virescens. J. Insect Behav. 2001, 14, 187–199. [Google Scholar] [CrossRef]
- Spielman, A.; Leahy, M.G.; Skaff, V. Failure of effective insemination of young female Aedes aegypti mosquitoes. J. Insect Physiol. 1969, 15, 1471–1479. [Google Scholar] [CrossRef]
- Herman, W.S. Juvenile hormone stimulation of tubular and accessory glands in male monarch butterflies. Comp. Biochem. Physiol. 1975, 51A, 507–510. [Google Scholar] [CrossRef]
- Herman, W.S.; Barker, J.F. Ecdysterone antagonism, mimicry, and synergism of juvenile hormone action on the monarch butterfly reproductive tract. J. Insect Physiol. 1976, 22, 643–648. [Google Scholar] [CrossRef]
- Brower, L.P.; Calvert, W.H.; Hedrick, L.E.; Christian, J. Biological observations on an overwintering colony of monarch butterflies (Danaus plxippus, Danaidae) in Mexico. J. Lepid. Soc. 1977, 31, 232–242. [Google Scholar]
- Urquhart, F.A.; Urquhart, N.R. Overwintering areas and migratory routes of the monarch butterfly (Danaus p. plexippus, Lepidoptera: Danaidae) in north America, with special reference to the western population. Can. Entomol. 1977, 109, 1583–1589. [Google Scholar] [CrossRef]
- Urquhart, F.A.; Urquhart, N.R. Vernal migration of the monarch butterfly (Danaus plexippus, Lepidoptera: Danaidae) in north America from the overwintering site in the neo-volcanic plateau of Mexico. Can. Entomol. 1979, 111, 15–18. [Google Scholar] [CrossRef]
- Walton, R.K.; Brower, L.P. Monitoring the fall migration of the monarch butterfly Danaus plexippus L. (Nymphalidae: Danainae) in Eastern North America: 1991–1994. J. Lepid. Soc. 1996, 50, 1–20. [Google Scholar]
- Tuskes, P.M.; Brower, L.P. Overwintering ecology of the monarch butterfly, Danaus plexippus L., in California. Ecol. Entomol. 1978, 3, 141–153. [Google Scholar] [CrossRef]
- Leopold, R.A. The role of male accessory glands in insect reproduction. Annu. Rev. Entomol. 1976, 21, 199–221. [Google Scholar] [CrossRef]
- Herman, W.S.; Peng, P. Juvenile hormone stimulation of sperm activator production in male monarch butterflies. J. Insect Physiol. 1976, 22, 579–581. [Google Scholar] [CrossRef]
- Hiroyoshi, S.; Reddy, G.V.P.; Mitsuhashi, J. Effects of juvenile hormone analogue (methoprene) and 20-hidroxyecdysone on reproduction in Polygonia c-aureum (Lepidoptera: Nymphalidae) in relation to adult diapause. J. Comp. Physiol. A 2017, 203, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Taylor, O.R., Jr. Relationship of multiple mating to fertility in Atteva punctella (Lepidoptera: Yponomeutidae). Ann. Entomol. Soc. Am. 1967, 60, 583–590. [Google Scholar] [CrossRef]
- Oberhauser, K.S. Rate of ejaculate breakdown and intermating intervals in monarch butterflies. Behav. Ecol. Sociobiol. 1992, 31, 367–373. [Google Scholar] [CrossRef]
- Pease, R.W., Jr. The evolutionary and biological significance of multiple pairing in Lepidoptera. J. Lepid. Soc. 1968, 22, 197–210. [Google Scholar]
- Drummond, B.A. Multiple mating and sperm competition in the Lepidoptera. In Sperm Competition and the Evolution of Animal Mating Systems; Smith, R.L., Ed.; Academic: Orlando, FL, USA, 1984; pp. 290–370. [Google Scholar]
- Zeh, J.A.; Zeh, D.W. Reproductive mode and the genetic benefits of polyandry. Anim. Behav. 2001, 61, 1051–1063. [Google Scholar] [CrossRef] [Green Version]
- Svärd, L.; Wiklund, C. Mass and production rate of ejaculates in relation to monandry. Behav. Ecol. Sociobiol. 1989, 24, 395–402. [Google Scholar] [CrossRef]
- Svärd, L.; McNeil, J.N. Female benefit, male risk: Polyandry in the true armyworm Pseupolyandry in butterflies. Daletia unipuncta. Behav. Ecol. Sociobiol. 1994, 35, 319–326. [Google Scholar]
- Sadek, M.M. Polyandry in field-collected Spotoptera littoralis moths and laboratory assessment of the effects of male mating history. Entomol. Exp. Appl. 2001, 98, 165–172. [Google Scholar] [CrossRef]
- Letsinger, J.T.; Gromko, M.H. The role of sperm numbers in sperm competition and female remating in Drosophila melanogaster. Genetica 1985, 66, 195–202. [Google Scholar] [CrossRef]
- Abraham, S.; Goane, L.; Rull, J.; Cladera, J.; Willink, E.; Vera, M.T. Multiple mating in Anastrepha fraterculus females and its relationship with fecundity and fertility. Entomol. Exp. Appl. 2011, 141, 15–24. [Google Scholar] [CrossRef]
- He, Y.; Miyata, T. Variations in sperm number in relation to larval crowding and spermatophore size in the armyworm, Pseudaletia separata. Ecol. Entomol. 1997, 22, 41–46. [Google Scholar] [CrossRef]
- Torres-Vila, L.M.; Rodríguez-Molina, M.C.; Gragera, J.; Bielza-Lino, P. Polyandry in Lepidoptera: A heritable trait in Spodoptera exigua Hübner. Heredity 2001, 86, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Ajula, M.; Rull, J.; Sivinski, J.; Trujillo, G.; Perez-Staples, D. Male and female condition influence mating performance and sexual receptivity in two tropical fruit flies (Diptera: Tephritidae) with contrasting life histories. J. Insect Physiol. 2009, 55, 1091–1098. [Google Scholar] [CrossRef]
- Knight, A.L. Multiple mating of male and female codling moth (Lepidoptera: Torticidae) in apple orchards treated with sex pheromone. Environ. Entomol. 2007, 36, 157–164. [Google Scholar] [CrossRef]
- Fortes, P.; Cônsoli, F. Are there costs in the repeated mating activities of female southern stink bugs Nezara viridula? Physiol. Entomol. 2011, 36, 215–219. [Google Scholar] [CrossRef]
- Oberhauser, K.S. Male monarch butterfly spermatophore mass and mating strategies. Anim. Behav. 1988, 36, 1384–1388. [Google Scholar] [CrossRef]
- Leahy, S.M.; Craig, G.B., Jr. Accessory gland substance as a stimulant for oviposition in Aedes aegypti and A. albopictus. Mosquito News 1965, 25, 448–452. [Google Scholar]
- Craig, G.B., Jr. Mosquitoes: Female monogamy induced by male accessory gland substance. Science 1967, 156, 1499–1501. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, J.G. Sperm activation in saturniid moths: Some aspects of the mechanism of activation. J. Insect Physiol. 1974, 20, 2321–2328. [Google Scholar] [CrossRef]
- Markow, T.A.; Ankney, P.F. Drosophila males contribute to oogenesis in a multiple mating species. Science 1984, 224, 302–303. [Google Scholar] [CrossRef] [PubMed]
- Young, A.D.M.; Downe, A.E.R. Male accessory gland substances and the control of sexual receptivity in female Culex tarsalis. Physiol. Entomol. 1987, 12, 233–239. [Google Scholar] [CrossRef]
- Herndon, L.A.; Wolfner, M.F. A Drosophila seminal fluid protein, Acp26Aa, stimulates egg laying in females for 1 day after mating. Proc. Natl. Acad. Sci. USA 1995, 92, 10114–10118. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.B. Effects of mating and accessory gland injections on olfactory-mediated behavior in the female Mediterranean fruit fly, Ceratitis capitata. J. Insect Physiol. 1995, 41, 705–710. [Google Scholar] [CrossRef]
- Bertram, M.J.; Neubaum, D.M.; Wolfner, M.F. Localization of the Drosophila male accessory gland protein Acp36DE in the mated female suggests a role in sperm storage. Insect Biochem. Mol. Biol. 1996, 26, 971–980. [Google Scholar] [CrossRef]
- Wolfner, M.F. Tokens of love: Functions and regulation of Drosophila male accessory gland product. Insect Biochem. Mol. Biol. 1997, 27, 179–192. [Google Scholar] [CrossRef]
- Delisle, J.; Picimbon, J.-F.; Simard, J. Regulation of pheromone inhibition in mated females of Choristoneura fumiferana and C. rosaceana. J. Insect Physiol. 2000, 46, 913–921. [Google Scholar] [CrossRef]
- Lung, O.; Kuo, L.; Wolfner, M.F. Drosophila males transfer antibacterial proteins from their accessory gland and ejaculatory duct to their mates. J. Insect Physiol. 2001, 47, 617–622. [Google Scholar] [CrossRef]
- Jin, Z.-Y.; Gong, H. Male accessory gland derived factors can stimulate oogenesis and enhance oviposition in Helicoverpa armigera (Lepidoptera: Noctuidae). Archiv. Insect Biochem. Phsiol. 2001, 469, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Sheng-li, S.I.; Shao-Fu, X.U.; Jia-Wei, D.U. Pheromonostatic activity of male accessory gland factors in female Helicoverpa assulta. Acta Entomol. Sin. 2000, 43, 120–126. [Google Scholar]
- Himuro, C.; Fujisaki, K. Males of the seed bug Togo hemipterus (Heteroptera: Lygaeidae) use accessory gland substances to inhibit remating by females. J. Insect Physiol. 2008, 54, 1538–1542. [Google Scholar] [CrossRef] [PubMed]
- Lamunyon, C.W. Increased fecundity, as a function of multiple mating, in an arctiid moth, Utetheisa ornatrix. Environ. Entomol. 1997, 22, 69–73. [Google Scholar] [CrossRef]
- Hidaka, T.; Aida, S. Day length as the main factor of seasonal form determination in Polygonia c-aureum (Lepidoptera, Nymphalidae). Zool. Mag. 1963, 72, 77–83. [Google Scholar]
- Fukuda, S.; Endo, K. Hormonal control of the development of seasonal forms in the butterfly, Polygonia c-aureum L. Proc. Jpn. Acad. 1966, 42, 1082–1087. [Google Scholar] [CrossRef]
- Fujita, K.; Inoue, M.; Watanabe, M.; Islam, A.T.M.F.; Shahjahan, R.M.; Endo, K.; Yamanaka, A. Photoperiodic regulation of reproductive activity in summer- and autumn-morph butterflies of Polygonia c-aureum L. Zool. Stud. 2009, 48, 291–297. [Google Scholar]
- Riemann, J.G.; Thorson, B.J.; Ruud, R.L. Daily cycle of release of sperm from the testes of the Mediterranean flour moth. J. Insect Physiol. 1974, 20, 195–207. [Google Scholar] [CrossRef]
- Hiroyoshi, S.; Mitsuhashi, J. Effects of environmental conditions and aging on eupyrene sperm movement in male adults of Polygonia c-aureum (Lepidoptera: Nymphalidae). Zool. Sci. 1998, 15, 561–565. [Google Scholar] [CrossRef]
- Thorson, B.J.; Riemann, J.G. Abdominal entrained periodicities of testis and vas deferens activity in the Mediterranean flour moth. J. Insect Physiol. 1977, 23, 1189–1197. [Google Scholar] [CrossRef]
- Riemann, J.G.; Johnson, M.; Thorson, B. Recovery of fertility by Mediterranean flour moths transferred from continuous light to light:dark. Ann. Entomol. Soc. Am. 1981, 74, 274–278. [Google Scholar] [CrossRef]
- Giebultowicz, J.M.; Brooks, N.L. The circadian rhythm of sperm release in the codling moth, Cydia pomonella. Entomolol. Exp. Appl. 1998, 88, 229–234. [Google Scholar] [CrossRef]
- Riemann, J.G.; Giebultowicz, J.M. Secretion in the upper vas deferens of the gypsy moth correlated with the circadian rhythm of sperm release from the testes. J. Insect Physiol. 1991, 37, 53–62. [Google Scholar] [CrossRef]
- Giebultowicz, J.M.; Riemann, J.G.; Raina, A.K.; Ridgway, R.L. Circadian system controlling release of sperm in the inset testes. Science 1989, 245, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Giebultowicz, J.M.; Bell, R.A.; Imberski, R.B. Circadian rhythm of sperm movement in the male reproductive tract of the gypsy moth, Lymantia dispar. J. Insect Physiol. 1988, 34, 527–553. [Google Scholar] [CrossRef]
- Giebultowicz, J.M.; Weyda, F.; Erbe, E.F.; Wergin, W.P. Circadian rhythm of sperm release in the gypsy moth, Lymantria dispar: Ultrastructural study of transepithelial penetration of sperm bundles. J. Insect Physiol. 1997, 43, 1133–1147. [Google Scholar] [CrossRef]
- Bebas, P.; Cymborowski, B.; Giebultowicz, J.M. Circadian rhythm of sperm release in males of the cotton leafworm, Spodoptera littoralis: In vivo and in vitro studies. J. Insect Physiol. 2001, 47, 859–866. [Google Scholar] [CrossRef]
- Herman, W.S.; Brower, L.P.; Calvert, W.H. Reproductive tract development in monarch butterflies overwintering in California and Mexico. J. Lepid. Soc. 1989, 43, 50–58. [Google Scholar]
- Hiroyoshi, S. Effects of aging, temperature and photoperiod on testis development of Polygonia c-aureum (Lepidoptera: Nymphalidae). Entomol. Sci. 2000, 3, 227–236. [Google Scholar]
- Chase, J.A.; Gilliland, F.R., Jr. Testicular development in the budworm. Ann. Entomol. Soc. Am. 1972, 65, 901–906. [Google Scholar] [CrossRef]
- Jeong, S.E.; Kown, S.B. Effects of inhibitors on polyamine metabolism during spermatogenesis of Heliothis assulta. Korean J. Entomol. 1986, 26, 299–311. [Google Scholar]
- Gongyin, Y.; Gu, H.; Shengquan, W. Structure and development of male internal reproductive organs in the Japanese oak silkworm, Antheraea yamamai. Acta Agrif. Zhejiang 1996, 8, 368–371. [Google Scholar]
- Holt, G.G.; North, D.T. Effects of gamma irradiation on the mechanism of sperm transfer in Trichoplusia ni. J. Insect Physiol. 1970, 16, 2211–2222. [Google Scholar] [CrossRef]
- Sridevi, R.; Dutta-Gupta, A.; Ramamurty, P.S. Spermatogenesis in Spodoptera litura (Lepidoptera: Noctuidae). Entomon 1989, 14, 1–10. [Google Scholar]
- Chandhury, M.F.B.; Raun, E.S. Spermatogenesis and testicular development of the European corn borer, Ostrinia nubilalis (Lepidoptera: Pyraustidae). Ann. Entomol. Soc. Am. 1966, 59, 1157–1159. [Google Scholar] [CrossRef]
- Reinecke, L.H.; Reinecke, J.P.; Adams, T.S. Morphology of the male reproductive tract of mature larval, pupal and adult tobacco hornworms (Lepidoptera: Sphingidae), Manduca sexta. Ann. Entomol. Soc. Am. 1983, 76, 365–375. [Google Scholar] [CrossRef]
- Salama, H.S. Spermatogenesis and testicular development in the gypsy moth Porthetria dispar L. Z. Angew. Entomol. 1976, 81, 102–110. [Google Scholar] [CrossRef]
- Scheepens, M.H.M.; Wysoki, M. Testicular development, spermatogenesis and chromosomes of Boarmia selenaria Schiff. (Lepidoptera: Geometridae). Int. J. Invertebr. Reprod. Dev. 1985, 8, 337–348. [Google Scholar] [CrossRef]
- Nishiitstsusuji-Uwo, J. Developmental features of the male germ cells in the prospective diapausing and non-diapausing swallowtails, Papilio xuthus. Mem. Coll. Sci. Univ. Kyoto Ser. B. 1959, 26, 9–14. [Google Scholar]
- Hidaka, T.; Takahashi, H. Temperature conditions and maternal effect as modifying factors in photoperiodic control of the seasonal form in Polygonia c-aureum (Lepidoptera, Nymphalidae). Annot. Zool. Jpn. 1967, 40, 200–204. [Google Scholar]
- Hiroyoshi, S. Effects of photoperiod and temperature on several pupal characters associated with imaginal polyphenism in Polygonia c-aureum (Lepidoptera: Nymphalidae). Appl. Entomol. Zool. 1992, 27, 155–159. [Google Scholar] [CrossRef]
- Hiroyoshi, S. Effects of photoperiod on the testis fusion in the Asian comma butterfly, Polygonia c-aureum Linnaeus (Lepidoptera: Nymphalidae). Entomon 2016, 41, 159–168. [Google Scholar]
- Ashrafi, S.H.; Brower, J.H.; Tilton, E.W. Gamma radiation effects on testes and on mating success of the Indian meal moth, Plodia interpunctella. Ann. Entomol. Soc. Am. 1972, 65, 1144–1149. [Google Scholar] [CrossRef]
- Cook, P.A.; Gage, M.J.G. Effects of risks of sperm competition on the numbers of eupyrene and apyrene sperm ejaculated by the moth Plodia interpunctella (Lepidoptera: Pyralidae). Behav. Ecol. Sociobiool. 1995, 36, 261–268. [Google Scholar] [CrossRef]
- Etman, A.A.M.; Hooper, G.H.S. Developmental and reproductive biology of Spodoptera litura (F.) (Lepidoptera: Noctuidae). J. Aust. Entomol. Soc. 1979, 18, 363–372. [Google Scholar] [CrossRef]
- Friedländer, M. Control of the eupyrene-apyrene sperm dimorphism in Lepidoptera. J. Insect Physiol. 1997, 43, 1085–1092. [Google Scholar] [CrossRef]
- Katsuno, S. Studies on eupyrene and apyrene spermatozoa in the silkworm, Bombyx mori L. (Lepidoptera: Bombycidae). IThe intratesticular behaviour of the spermatozoa at various stages from the 5th-instar to the adult. Appl. Entomol. Zool. 1977, 12, 142–153. [Google Scholar] [CrossRef]
- Numata, H.; Hidaka, T. Development of male sex cells in the swallowtail, Papilio Xuthus L. (Lepidoptera: Papilionidae) after the termination of diapause. Appl. Entomol. Zool. 1981, 16, 313–314. [Google Scholar] [CrossRef]
- Katsuno, S. Time of differentiation and determination of eupyrene and apyrene spermatozoa and resulting spermatogenesis in the silkworm, Bombyx mori L. (Lepidoptera: Bombycidae). Appl. Entomol. Zool. 1987, 12, 142–153. [Google Scholar] [CrossRef]
- Endo, K. Relation between ovarian maturation and activity of the corpora allata in seasonal forms of the butterfly, Polygonia c-aureum L. Dev. Growth Differ. 1970, 11, 297–304. [Google Scholar] [CrossRef]
- Endo, K. Activation of the corpora allata in relation to ovarian maturation in the seasonal forms of the butterfly, Polygonia c-aureum L. Dev. Growth Differ. 1972, 14, 263–274. [Google Scholar] [CrossRef]
- Endo, K. Hormonal regulation of mating in the butterfly, Polygonia c-aureum L. Dev. Growth Differ. 1973, 15, 1–10. [Google Scholar] [CrossRef]
- Hiroyoshi, S. Eupyrene and apyrene spermatogenesis in the Asian comma butterfly, Polygonia c-aureum (Lepidoptera: Nymphalidae). Entomol. Sci. 1999, 2, 297–305. [Google Scholar]
- Hiroyoshi, S.; Yoshimura, J.; Iwabuchi, K.; Reddy, G.V.P.; Mitsuhashi, J. Effects of pre-overwintering conditions on eupyrene and apyrene spermatogenesis after overwintering in Polygonia c-aureum (Lepidoptera: Nymphalidae). J. Insect Physiol. 2017, 100, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hiroyoshi, S. Effects of photoperiod and age on the initiation of sperm movement in male Polygonia c-aureum LINNAEUS (Lepidoptera: Nymphalidae). Appl. Entomol. Zool. 1997, 32, 19–25. [Google Scholar] [CrossRef]
- Tsubaki, Y.; Yamagishi, M. “Longevity” of sperm within the female of the melon fly, Dacus cucurbitae (Diptera: Tephritidae), and its relevance to sperm competition. J. Insect Behav. 1991, 4, 243–250. [Google Scholar] [CrossRef]
- Stewart, A.D.; Hannes, A.M.; Rice, W.R. An assessment of sperm survival in Drosophila melanogaster. Evolution 2007, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Bertin, S.; Scolari, F.; Guglielmino, C.R.; Bonizzoni, M.; Bonomi, A.; Marchini, D.; Gomulski, L.M.; Gasperi, G.; Malacrida, A.R.; Matessi, C. Sperm storage and use in polyandrous females of the globally invasive fruitfly, Ceratitis capitata. J. Insect Physiol. 2010, 56, 1452–1551. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, P.; Fedorka, K.M. Influence of female age, sperm senescence and multiple mating on sperm viability in female Drosophila melanogaster. J. Insect Physiol. 2011, 57, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Riemann, J.G.; Thorson, B.J. Ultrastructure of the vasa deferentia of the Mediterranean flour moth. J. Morphol. 1976, 149, 483–506. [Google Scholar] [CrossRef] [PubMed]
- Riemann, J.G.; Thorson, B.J. Ultrastructure of the ductus ejaculatoris duplex of the Mediterranean flour moth, Anagasta kühniella (Zeller) (Lepidoptera: Pyralidae). Int. J. Insect Morphol. Embryol. 1976, 5, 227–240. [Google Scholar] [CrossRef]
- Burns, J.M. Mating frequency in natural populations of skippers and butterflies as determined by spermatophore counts. Proc. Nat. Acad. Sci. USA 1968, 61, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Janowitz, S.A.; Fischer, K. Costing reproduction: Effects of mating opportunity on mating success in male Bicyclus anynana butterflies. Behav. Ecol. Sociobiol. 2010, 64, 1999–2006. [Google Scholar] [CrossRef]
- Takeuchi, T. Cost of reproduction in males of a satyrine butterfly Lethe diana. Physiol. Entomol. 2012, 37, 171–176. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M.; McNeil, J.N. Lifetime mating potential and reproductive success in males of the true armyworm, Pseudaletia unipuncta (Haw.) (Lepidoptera: Noctuidae). Funct. Ecol. 1989, 3, 37–44. [Google Scholar] [CrossRef]
- Royer, L.; McNeil, J.N. Male investment in the European corn borer, Ostrinia nubilalis (Lepidoptera: Pyralidae): Impact on female longevity and reproductive performance. Funct. Ecol. 1993, 7, 209–215. [Google Scholar] [CrossRef]
- Reinhardt, K. Determinants of ejaculates size in a grasshopper (Chorthippus parallelus). Behav. Ecol. Sociobiol. 2001, 50, 503–510. [Google Scholar] [CrossRef]
- Amin, M.R.; Bussière, L.F.; Goulson, D. Effects of male age and size on mating success in the Bumblebee Bombus terrestris. J. Insect Behav. 2012, 25, 362–374. [Google Scholar] [CrossRef]
- Proshold, F.I. Number of sperm bundles in the duplex of tobacco budworms (Lepidoptera: Noctuidae) as a function of age. J. Econ. Entomol. 1991, 84, 1485–1491. [Google Scholar] [CrossRef]
- Proshold, F.I. Mating status of tobacco budworm (Lepidoptera: Noctuidae) males captured in pheromone traps compared with males and females collected at night by hand. J. Econ. Entomol. 1991, 84, 1492–1498. [Google Scholar] [CrossRef]
- Spurgeon, D.W.; Raulston, J.R.; Lingren, P.D.; Shaver, T.N.; Proshold, F.I.; Gillespie, J.M. Temporal aspects of sperm transfer and spermatophore condition in Mexican rice borers (Lepidoptera: Pyralidae). J. Econ. Entomol. 1994, 87, 371–376. [Google Scholar] [CrossRef]
- Chien, C.I.; Chow, Y.S. Changes in the ductus ejaculatorius simplex and testes of adult males of Spodoptera litura (F.) with age and mated status. Plant Prot. Bull. 1985, 27, 379–387. [Google Scholar]
- Snow, J.W.; Carlysle, T.C. A characteristic indicating the mating status of male fall armyworm moths. Ann Entomol. Soc. Am. 1967, 60, 1071–1074. [Google Scholar] [CrossRef]
- Haines, L.C. Changes in colour of a secretion in the reproductive tract of adult males of Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae) with age and mated status. Bull. Ent. Res. 1981, 71, 591–598. [Google Scholar] [CrossRef]
- Krishnananda, N.; Satyanrayana, S.V.V. Differentiation between mated and virgin males in Spodoptera litura. Phytoparasitica 1988, 16, 351–354. [Google Scholar] [CrossRef]
- Goehring, L.; Oberhauser, K.S. Effects of photoperiod, temperature, and host plant age on induction of reproductive diapause and development time in Danaus plexippus. Ecol. Entomol. 2002, 27, 674–685. [Google Scholar] [CrossRef]
- Koyama, J. The sexual and physiological character in the mating behavior of Fannia scalaris FABRICIUS. Jpn. J. Ecol. 1974, 24, 92–115, (Japanese with English Synopsis). [Google Scholar]
- Kato, Y. The prediapause copulation and its significance in the butterfly Eurema hecabe. J. Ethol. 1986, 4, 81–90. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photoperiod/Adult Age | No. of Insects Used | No. of Eggs (Mean ± S.D.) | Oocyte Diameter (mm) (Mean ± S.D.) |
|---|---|---|---|
| Short day | |||
| 0 | 9 | 0.00 ± 0.00 | 0.159 ± 0.019 |
| 10 | 40 | 0.00 ± 0.00 * | 0.199 ± 0.014 |
| 20 | 32 | 0.00 ± 0.00 * | 0.187 ± 0.029 * |
| 30 | 26 | 0.00 ± 0.00 * | 0.148 ± 0.011 * |
| 40 | 20 | 0.80 ± 0.60 * | 0.294 ± 0.059 * |
| 50 | 21 | 10.20 ± 5.20 * | 0.331 ± 0.059 * |
| 60 | 19 | 9.80 ± 5.00 * | 0.400 ± 0.057 * |
| Long day | |||
| 10 | 29 | 3.70 ± 2.00 | 0.302 ± 0.041 |
| 20 | 24 | 22.30 ± 5.20 | 0.529 ± 0.045 |
| 30 | 9 | 46.00 ± 11.60 | 0.684 ± 0.041 |
| 40 | 8 | 27.50 ± 7.70 | 0.650 ± 0.043 |
| 50 | 14 | 46.40 ± 7.70 | 0.687 ± 0.023 |
| 60 | 12 | 58.70 ± 8.20 | 0.676 ± 0.029 |
| Seasonal Form | Date of Collection | No. of Insects Used | No. of Insects with Egg in the Ovary (%) | Percentages of Females with Spermatophore |
|---|---|---|---|---|
| Summer | 3-June-1992 | 6 | - | 83.3 |
| Summer | 4-September-1992 | 11 | 11, 100 | 100 |
| Summer | 18-September-1992 | 8 | 8, 100 | 100 |
| Autumn | 3-April-1992 | 5 | - | 100 |
| Autumn | 18-September-1992 | 6 | 0, 0 | 0 |
| Autumn | 11-October-1992 | 20 | 0, 0 | 0 |
| Autumn | 12-November-1992 | 1 | 0, 0 | 0 |
| Seasonal Form/Date | Site | No. of Animals Used | Spermatophore Size (mm3) | |||
|---|---|---|---|---|---|---|
| No. of Spermatophore (Mean ± S.D.) From the Newest To the Oldest | ||||||
| 1 | 2 | 3 | 4 | |||
| Autumn | ||||||
| 3-April-1992 | Tokyo | 6 | 53.9 ± 24.6 (6) | 63.2 ± 50.9 (2) | ||
| 18-September-1992 | Saitama | 6 | ||||
| 11-October-1992 | Saitama | 20 | ||||
| 12-November-1992 | Saitama | 1 | ||||
| Summer | ||||||
| 3-June-1992 | Saitama | 5 | 68.5 ± 26.6 (5) | |||
| 4-September-1992 | Saitama | 11 | 35.4 ± 22.4 a (11) | 30.8 ± 13.0 b (7) | 47.1 ± 21.3 a,c (7) | 45.2 ± 22.1 a,b (3) |
| 18-September-1992 | Saitama | 8 | 34.5 ± 22.9 (11) | 25.1 ± 19.7 (6) | 41.1 ± 25.5 (5) | |
| No. of Insects Used | Min | Max | Mean ± S.D. | |
|---|---|---|---|---|
| Mating frequency | 10 | 0 | 15 | 5.11 ± 4.81 |
| Weight of SP (mg) per mating | 10 | 0 | 3.55 | 1.93 ± 0.46 |
| Volume of SP (mm3) per mating | 10 | 0 | 123.76 | 59.69 ± 48.68 |
| Total volume of SP (mm3) | 10 | 0 | 288.04 | 209.45± 57.93 |
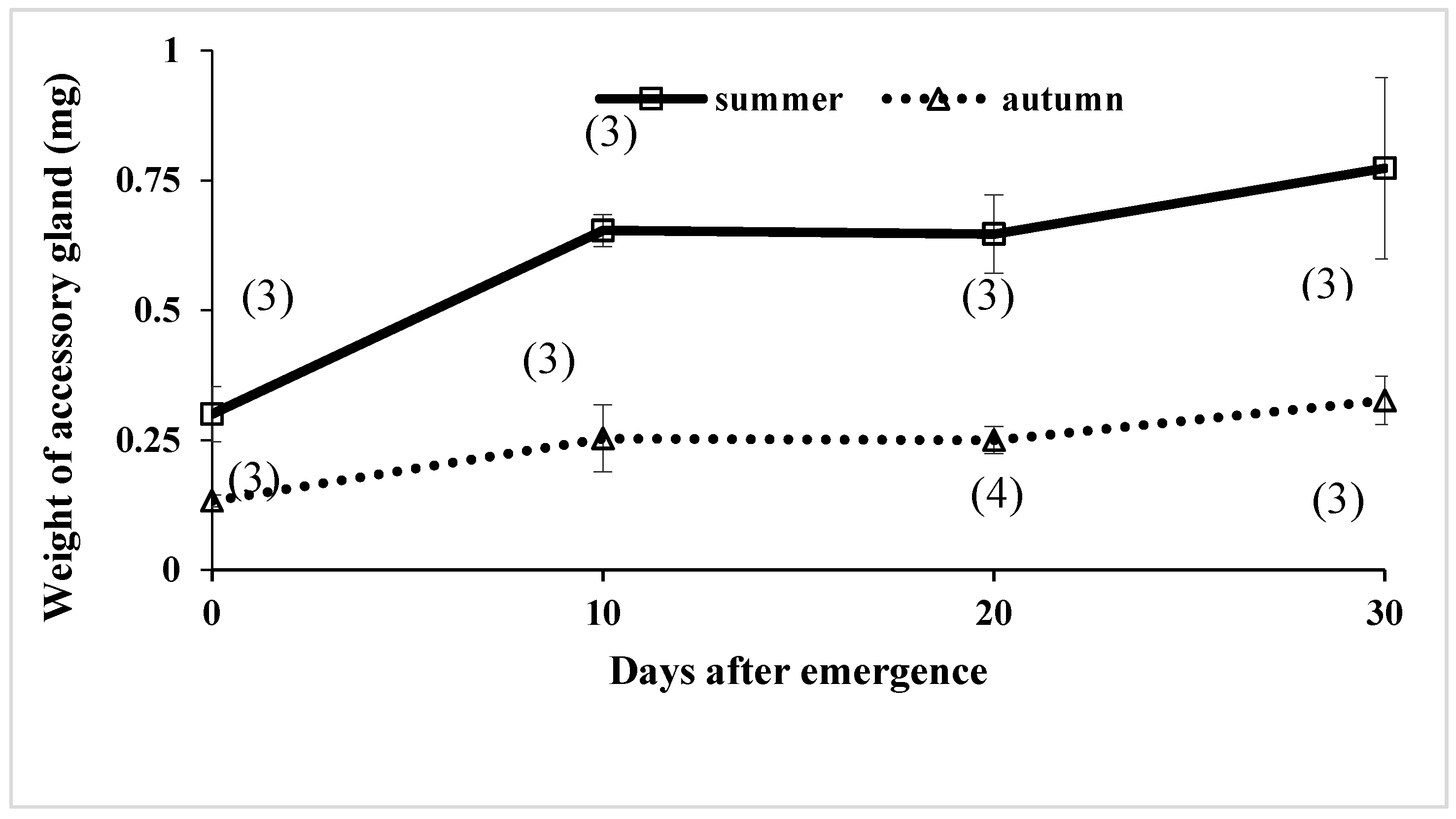
| Date of Collection | Site of Collection | Testis Volume (mm3) (Mean ± S.D.) | Accessory Gland Dry Weight (mg) (Mean ± S.D.) | Simplex Dry Weight (mg) (Mean ± S.D.) |
|---|---|---|---|---|
| Autumn form | ||||
| 22-April-1991 | Akigase | 6.51 ± 1.613 (4) *a | 0.80 ± 0.356(4)a | 4.00 ± 2.592(4)a |
| 31-March-1992 | Akigase | 2.95 ± 1.493(4)a | 0.48 ± 0.382(3)ab | 3.13 ± 2.730(3)ab |
| 4-April-1992 | Akigase | 4.17 ± 1.260(6)a | 0.39 ± 0.163(6)b | 2.65 ± 1.170(6)ab |
| 4-September-1992 | Akigase | 13.19 (1) | 1.60 (1) | |
| 18-September-1992 | Akigase | 12.89 ± 4.079(7)b | 0.20 ± 0.071(6)bc | 1.83 ± 0.523b |
| 11-October-1992 | Akigase | 13.13 ± 2.505(26)b | 0.13 ± 0.094(27)c | 1.90 ± 0.523(27)b |
| 12-November-1992 | Akigase | 5.49 ± 0.950(4)a | 0.20 ± 0.115(4)bc | 2.15 ± 1.158(4)ab |
| Summer form | ||||
| 13-June-1991 | Akigase | 13.12 ± 6.349(13)ab | 0.58 ± 0.269(14)ab | 3.76 ± 1.615(14)a |
| 15-July-1991 | Akigase | 12.85 ± 3.115(12)ab | 0.44 ± 0.152(5)ab | 2.66 ± 0.826(5)ab |
| 19-August-1991 | Akigase | 15.93 ± 4.259(12)ab | 0.39 ± 0.272(12)ab | 3.25 ± 1.566(12)ab |
| 3-September-1991 | Akigase | 14.45 ± 5.575(4)b | 1.83 ± 0.814(4)ab | |
| 3-June-1992 | Akigase | 12.08 ± 2.764(10)ab | 0.70 ± 0.372(10)a | 3.03 ± 1.231(10)ab |
| 20-August-1992 | Tsurugashima | 12.62 ± 1.669(5)ab | 0.48 ± 0.084(5)b | 2.33 ± 1.060(5)ab |
| 4-September-1992 | Akigase | 10.37 ± 2.659(16)abc | 0.35 ± 0.246(16)bc | 2.79 ± 1.407(16)ab |
| 18-September-1992 | Akigase | 6.13 ± 1.489(11)c | 0.30 ± 0.297(11)bc | 1.76 ± 0.988(11)b |
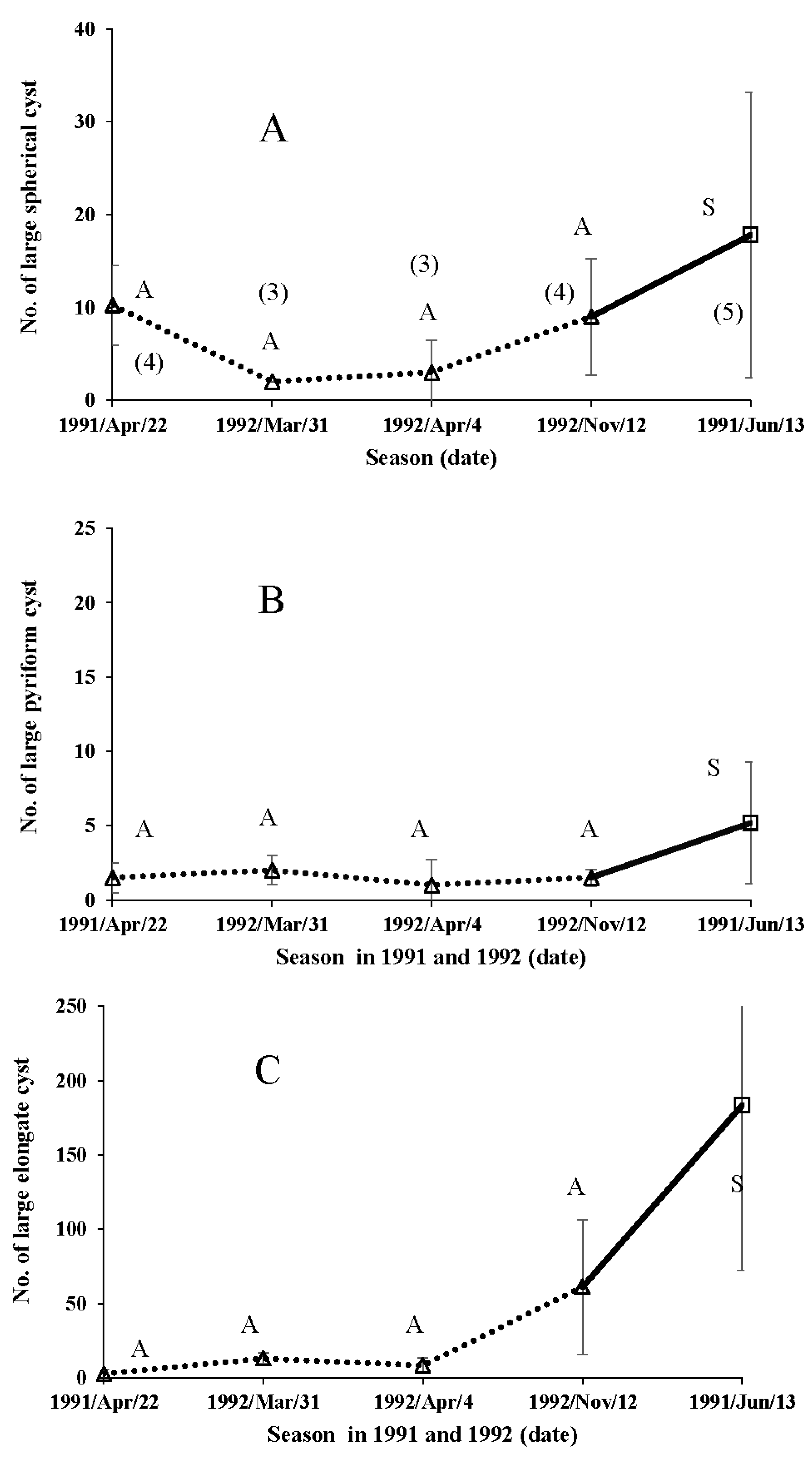
| Date of Collection | Site of Collection | No. of Insects Used | No. of ESB in the Duplex (Mean ± S.D.) |
|---|---|---|---|
| Autumn form | |||
| 22-April-1991 | Akigase | 4 | 312 ± 179.1abc |
| 31-March-1992 | Kominawa | 3 | 427 ± 284.0b |
| 4-April-1992 | Akigase | 6 | 481 ± 163.0b |
| 18-September-1992 | Akigase | 4 | 105 ± 62.3a |
| 11-October-1992 | Akigase | 7 | 86 ± 51ab |
| 12-November-1992 | Akigase | 10 | 403 ± 166.4c |
| Summer form | |||
| 13-June-1991 | Akigase | 8 | 240 ± 206.2a |
| 15-July-1991 | Akigase | 5 | 135 ± 52.2a |
| 19-August-1991 | Akigase | 9 | 166 ± 124.8a |
| 3-September-1991 | Akigase | 4 | 93 ± 79.6a |
| 3-June-1992 | Akigase | 5 | 70 ± 28.4a |
| 20-August-1992 | Tsurugashima | 5 | 266 ± 375.6a |
| 4-September-1992 | Akigase | 9 | 214 ± 134.2a |
| 18-September-1992 | Akigase | 9 | 274 ± 149.6a |
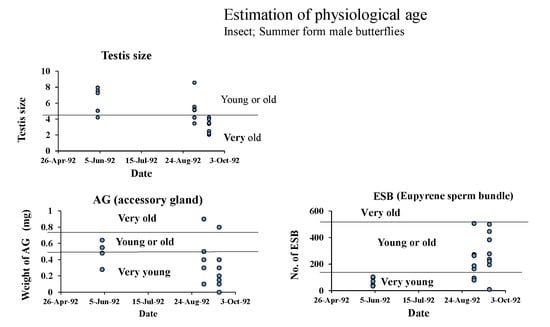
| Date of Collection | Individual No. | Testis Size (mm3) | Accessory Gland Weight (mg) | Simplex Weight (mg) | No. of ESB in the Duplex | Justice Mated or Not |
|---|---|---|---|---|---|---|
| 1991 | ||||||
| 22-April | 29 | 6.40 (O) | 1.0 (VO) | 6.4 (O) | 550 (O) | unmated |
| 30 | 4.45 (VO) | 0.5 (O) | 2.4 (Y or O) | 309 (Y or O) | mated | |
| 31 | 6.84 (O) | 0.5 (O) | 1.2 (VY) | 117 (VY) | mated | |
| 32 | 8.37 (O) | 1.2(VO) | 6.0 (O) | 273 (Y or O) | unmated | |
| 1992 | ||||||
| 31-March | 3415 | 1.86 (VO) | 0.9 (VO) | 5.6 (O) | 754 (O) | unmated |
| 3416 | 2.34 (VO) | 0.15 (VY) | 0.2 (VY) | 245 (Y or O) | mated | |
| 3417 | 0.84 (VO) | 0.4 (O) | 3.6 (Y or O) | 281 (Y or O) | mated | |
| 4-April | 17 | 1.18 (VO) | 0.4 (O) | 1.2 (VY) | 325 (Y or O) | mated |
| 18 | 2.34 (VO) | 0.7 (VO) | 4.7 (O) | 769 (O) | unmated | |
| 19 | 2.38 (VO) | 0.25 (Y or O) | 2.6 (Y or O) | 476 (Y or O) | mated | |
| 20 | 2.90 (VO) | 0.4 (Y or O) | 3.1 (Y or O) | 500 (Y or O) | unmated | |
| 21 | 1.51 (VO) | 0.3 (Y or O) | 2.5 (Y or O) | 498 (Y or O) | mated | |
| 22 | 2.22 (VO) | 0.3 (Y or O) | 1.9 (Y or O) | 323 (Y or O) | mated |
| Date of Collection | Individual No. | Testis Size (mm3) | Accessory Gland Weight (mg) | Simplex Weight (mg) | No. of ESB in the Duplex | Justice Mated or Not |
|---|---|---|---|---|---|---|
| 13-June | 247 | 4.34 (VO) | 0.6 (Y) | 3.9 (Y or O) | 294 (Y or O) | mated |
| 248 | 3.29 (VO) | 0.3 (VY) | 2.4 (VY) | 224 (Y or O) | mated | |
| 249 | 3.55 (VO) | 0.8 (O) | 4.9 (Y or O) | 496 (Y or O) | unmated | |
| 250 | 5.72 (O) | 0.7 (O) | 4.8 (Y or O) | 87 (VY) | mated | |
| 251 | 4.03 (VO) | 0.2 (VY) | 1.3 (VY) | 43 (VY) | mated | |
| 252 | 11.43 (Y) | 0.4 (VY) | 2.4 (VY) | 81 (VY) | unmated | |
| 253 | 12.58 (Y) | 0.8 (O) | 5.2 (Y or O) | 102(VY) | mated | |
| 254 | 4.50 (VO) | 0.2 (VY) | 6.9 (O) | 590 (O) | unmated? | |
| 15-July | 695 | 7.00 (O) | 0.6 (Y or O) | 3.8 (Y or O) | 140 (VY) | mated |
| 696 | 9.31 (O) | 0.4 (VY) | 1.7 (VY) | 104 (VY) | mated | |
| 697 | 10.02 (Y) | 0.6 (Y) | 3.1 (VY) | 148 (VY) | unmated | |
| 698 | 6.07 (O) | 0.3 (VY) | 2.6 (VY) | 72 (VY) | mated | |
| 699 | 5.41 (O) | 0.3 (VY) | 2.1(VY) | 211 (Y or O) | mated | |
| 19-August | 1068 | 9.82 (O) | 1.1 (O) | 6.3 (O) | 191 (VY) | mated |
| 1069 | 9.54 (O) | 0.3 (VY) | 4.5 (Y or O) | 79 (VY) | mated | |
| 1070 | 10.37 (Y) | 0.4 (VY) | 4.1 (Y or O) | 105 (VY) | unmated | |
| 1071 | 7.22 (O) | 0.25 (VY) | 1.4 (VY) | 218 (Y or O) | mated | |
| 1072 | 8.30 (O) | 0.5 (Y) | 2.6 (VY) | 78 (VY) | mated | |
| 1073 | 4.34 (VO) | 0.5 (Y) | 3.2 (VY) | 457 (Y or O) | mated | |
| 1074 | 11.50 (Y) | 0.4 (VY) | 5.1 (Y or O) | 151 (VY) | unmated | |
| 1075 | 4.87 (VO) | 0.1 (VY) | 1.8 (VY) | 179 (VY) | mated | |
| 1076 | 7.58 (O) | 0.1 (VY) | 1.1 (VY) | 35 (VY) | mated | |
| 3-September | 1359 | 10.35 (Y) | 0.5 (Y) | 2.9 (VY) | 94 (VY) | unmated |
| 1360 | 4.16 (VO) | 0.1 (VY) | 1.1 (VY) | 204 (Y or O) | mated | |
| 1361 | 8.63 (O) | 0.1 (VY) | 2.0 (VY) | 26(VY) | mated | |
| 1362 | 5.76 (O) | 0 (VY) | 1.3 (VY) | 46 (VY) | mated |
| Date of Collection | Individual No. | Testis Size (mm3) | Accessory Gland Weight (mg) | Simplex Weight (mg) | No. of ESB in the Duplex | Justice Mated or Not |
|---|---|---|---|---|---|---|
| 3-June | 246 | 7.54 (VO) | 0.48 (VY) | 4.8 (Y or O) | 99 (VY) | mated |
| 247 | 5.04 (VO) | 0.55 (Y) | 4.45 (Y or O) | 70 (VY) | mated | |
| 248 | 4.19 (VO) | 0.28 (VY) | 1.7 (VY) | 42 (VY) | mated | |
| 249 | 7.94 (VO) | 0.64 (Y) | 4.4 (Y or O) | 103 (VY) | mated | |
| 250 | 7.25 (VO) | 0.28 (VY) | 1.7 (VY) | 35 (VY) | mated | |
| 20-August | 944 | 5.95 (VO) | 0.4 (VY) | 1.05 (VY) | 147 (VY) | mated |
| 946 | 6.81 (VO) | 0.6 (Y) | 3.5 (VY) | 78 (VY) | mated | |
| 947 | 5.62 (VO) | 0.5 (Y) | 3.3 (VY) | 78 (VY) | mated | |
| 948 | 7.46 VO) | 0.5 (Y) | 2.2 (VY) | 93 (VY) | mated | |
| 949 | 5.74 (VO) | 0.4 (VY) | 1.6 (VY) | 91 (VY) | mated | |
| 4-September | 1134 | 8.56 (O) | 0.1 (VY) | 1.5 (VY) | 95 (VY) | mated |
| 1135 | 5.52 (VO) | 0.5 (Y) | 5.0 (Y or O) | 272 (Y) | mated | |
| 1136 | 5.10 (VO) | 0.1 (VY) | 2.2 (VY) | 93 (VY) | mated | |
| 1137 | 5.22 (VO) | 0.5 (Y) | 2.8 (VY) | 506(O) | mated | |
| 1138 | 5.16 (VO) | 0.5 (Y) | 2.95 (VY) | 269 (Y) | mated | |
| 1139 | 4.21 (VO) | 0.9 (O) | 4.7 (Y or O) | 165 (VY) | mated | |
| 1144 | 4.18 (VO) | 0.1 (VY) | 1.4 (VY) | 79 (VY) | mated | |
| 1146 | 5.13 (VO) | 0.3 (VY) | 2.45 (Y or O) | 263 (Y) | mated | |
| 1147 | 3.44 (VO) | 0.4 (VY) | 2.5 (Y or O) | 188 (VY) | mated | |
| 18-September | 1402 | 3.43 (VO) | 0.2 (VY) | 1.9 (VY) | 277 (Y) | mated |
| 1412 | 2.47 (VO) | 0.4 (VY) | 1.85 (VY) | 446 (Y) | mated | |
| 1424 | 2.03 (VO) | 0.1 (VY) | 1.9 (VY) | 500 (Y) | mated | |
| 1426 | 4.21 (VO) | 0 (VY) | 1.6 (VY) | 195(VY) | mated | |
| 1427 | 3.97 (VO) | 0.8 (O) | 4.4 (Y or O) | 383 (Y) | mated | |
| 1428 | 3.52 (VO) | 0 (VY) | 0.6 (VY) | 9 (VY) | mated | |
| 1429 | 3.40 (VO) | 0.2 (VY) | 1.1 (VY) | 236 (Y) | mated | |
| 1430 | 2.18 (VO) | 0.3 (VY) | 1.3 (VY) | 195 (VY) | mated | |
| 1431 | 3.47 (VO) | 0.15 (VY) | 1.2 (VY) | 222 (Y) | mated |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hiroyoshi, S.; Reddy, G.V.P. Field and Laboratory Studies on the Ecology, Reproduction, and Adult Diapause of the Asian Comma Butterfly, Polygonia c-aureum L. (Lepidoptera: Nymphalidae). Insects 2018, 9, 169. https://doi.org/10.3390/insects9040169
Hiroyoshi S, Reddy GVP. Field and Laboratory Studies on the Ecology, Reproduction, and Adult Diapause of the Asian Comma Butterfly, Polygonia c-aureum L. (Lepidoptera: Nymphalidae). Insects. 2018; 9(4):169. https://doi.org/10.3390/insects9040169
Chicago/Turabian StyleHiroyoshi, Satoshi, and Gadi V. P. Reddy. 2018. "Field and Laboratory Studies on the Ecology, Reproduction, and Adult Diapause of the Asian Comma Butterfly, Polygonia c-aureum L. (Lepidoptera: Nymphalidae)" Insects 9, no. 4: 169. https://doi.org/10.3390/insects9040169