
Regulation and Functional Significance of 5-Hydroxymethylcytosine in Cancer
by
Vittal Rangan Arvinden
, 
Arunagiri Kuha Deva Magendhra Rao
,
Thangarajan Rajkumar
and
Samson Mani
* Department of Molecular Oncology, Cancer Institute (WIA), 38 Sardar Patel Road, Chennai 600036, India
*
Author to whom correspondence should be addressed.
Epigenomes 2017, 1(3), 19; https://doi.org/10.3390/epigenomes1030019
Submission received: 22 September 2017
/
Revised: 6 November 2017
/
Accepted: 16 November 2017
/
Published: 23 November 2017
Abstract
:Epigenetic modes of gene regulation are important for physiological conditions and its aberrant changes can lead to disease like cancer. 5-hydroxymethylcytosine (5hmC) is an oxidized form of 5-methylcytosine (5mC) catalyzed by Ten Eleven Translocation (TET) enzymes. 5hmC is considered to be a demethylation intermediate and is emerging as a stable and functional base modification. The global loss of 5hmC level is commonly observed in cancers and tumorigenic germline mutations in IDH, SDH and FH are found to be inhibiting TET activity. Although a global loss of 5hmC is characteristic in cancers, locus-specific 5hmC gain implicates selective gene expression control. The definitive role of 5hmC as a tumor suppressing or promoting modification can be deduced by identifying locus-specific 5hmC modification in different types of cancer. Determining the genes carrying 5hmC modifications and its selective variation will open up new therapeutic targets. This review outlines the role of global and locus-specific changes of 5hmC in cancers and the possible mechanisms underlying such changes. We have described major cellular factors that influence 5hmC levels and highlighted the significance of 5hmC in tumor micro environmental condition like hypoxia.
1. Introduction
Cytosine methylation (5mC) plays a vital role in development, differentiation and gene expression modulation and its mechanism of reversal were previously thought to be mediated by AID/APOBEC or by passive removal through the nuclear exclusion of DNA methyl transferase 1 (DNMT1) during replication [1,2]. 5-hydroxymethyl cytosine (5hmC) is considered as the sixth base of the genome as well as the second most significant DNA modification after 5mC [3,4]. The discovery of the role of Ten Eleven Translocation enzymes (TET) in the conversion of 5mC to 5hmC and their ability to further oxidise 5hmC → 5-formylcytosine (5fC) → 5-carboxycytosine (5caC) led to the understanding of active demethylation pathway in mammalian genome [5,6,7,8]. The global levels of 5hmC vary among tissues with highest levels in the brain followed by the liver whereas the same is low in the heart as well as placental tissues [9,10]. Though 5mC variants have a definitive function in the demethylation pathway, distinct binding proteins of each 5mC variant make it worthwhile to consider them as individual modifications [11,12]. As an individual base modification 5hmC maintenance over replication is necessary, but DNMT1 poorly methylates hemi hydroxymethylated DNA in vitro that invokes loss of maintenance of 5hmC during cell divisions [13,14]. However, 5hmC levels are not significantly reduced after S-phase, making them a stable DNA modification and supports its maintenance in vivo after cell division [15]. 5hmC levels are dramatically reduced in cancer tissues compared to adjacent normal tissue [16,17]. Mutations in genes like isocitrate dehydrogenase (IDH1/2), succinate dehydrogenase (SDH) and fumarate hydratase (FH); which control 5hmC genomic levels by regulating TET activity are reported in various cancers [18,19,20]. Though mutational (IDH, SDH and FH) imbalance in methylation machinery results in the loss of 5hmC and trigger carcinogenesis, locus-specific changes of 5hmC between tumor and normal tissues is also important for cancer progression.
In this review, we focus on the role of 5-hydroxymethylcytosine in cancer, cellular factors regulating its levels, mechanism of locus-specific 5hmC changes, influence of environmental factors like hypoxia, reactive oxygen species (ROS), heavy metals and carcinogens disturbing the balance between methylcytosine and hydroxymethylcytosine.
2. Interplay between 5-Hydroxymethylcytosine and Cancer
The global loss of 5hmC in cancerous cells portrays it as a crucial pre-transcriptional gene regulatory mechanism. Invariably reduced levels of 5hmC have been reported in tumors of the lung, colon, brain, breast, liver, prostate, kidney, melanoma, etc, compared to its respective normal tissues [21,22,23]. This loss of 5hmC in genomic context may lead to gene body hypermethylation as observed in kidney cancer and can hypermethylate promoters of tumor suppressor genes (TSG) that are frequently observed in cancers [24]. Reduced 5hmC is a predictive marker for poor prognosis and survival for multiple cancers like breast, astrocytoma, laryngeal squamous cell carcinoma, renal, oesophageal, gastric and hepatocellular carcinoma (HCC) [24,25,26,27,28,29,30]. Moreover, mutations in TET2 are frequently observed with decrease in 5hmC in hematological cancers, additionally mutations of 5hmC regulating enzymes like IDH1/2, SDH and FH were also observed [19,20,31,32]. Mutations in IDH1/2 (R132H/C/S and R172X) cause accumulation of 2-hydroxyglutarate (2-HG) by gaining neomorphic activity to convert 2-oxoglutarate (2-OG) to 2-HG [18]. Similarly, a mutation in SDH and FH would increase the cellular concentration of succinate and fumarate. These three metabolites succinate, fumarate and 2-HG are competitive inhibitors of 2-OG dependent dioxygenases like the TET enzymes and hence reduce global 5hmC levels in genomic DNA [33,34]. Notably, IDH mutations have frequently been reported in acute myeloid luekemia (AML), glioblastoma, melanoma and cancers of colon and lung [32]. Mutations in SDH are mostly reported in hereditary paraganglioma/phaeochromocytoma syndrome (HPGL/PCC), pancreatic neuroendocrine tumors and renal and gastrointestinal stromal tumors [19]. While mutations in FH have been reported in tumors of uterine and cutaneous leiomyomas, leiomyosarcoma and renal cell carcinoma (Table 1 describes different types of cancer that are reported with IDH, SDH and FH mutations) [20].
Despite frequently encountered global loss in cancers, locus-specific differential hydroxymethylation modulates expression of critical genes that promote survival and progression (Table 2). Several studies that have identified such loci describe the basis of 5hmC mediated gene regulation and also identify potential and novel regions that can be involved in regulation across the genome. Loss of 5hmC has been observed in Homeobox A Cluster (HOXA) genes, mir-200 and Leucine Zipper Tumor Suppressor 1 (LZTS1), leading to hypermethylation in those regions and promoting breast tumor progression and metastasis and in contrast, 5hmC gain at regions of (Tumor Necrosis Factor-Alpha) TNFA enable cell survival and stemness in breast tumor initiating cells (BTICs) under hypoxia [40,41,42,43]. Superoxide Dismutase 3 (SOD3), which is frequently downregulated in squamous cell carcinomas and adenocarcinomas, is also less expressed in lung (A549), breast (ZR75-1) and prostate (PC3) cancer cell lines as it incurs loss of 5hmC at its promoter region [44]. In colorectal cancers, genes differentially hydroxymethylated between tumor and paired normal samples showed high correlation with their mRNA expression [45]. In addition, Dickkopf-related protein 3 (DDK3) was hypermethylated in colon cancer, causing to an active WNT signaling mediated tumor growth in mice [46].
Increased global levels of 5hmC are also observed in certain types of cancers under certain conditions. In MLL rearranged leukemias, TET1 is upregulated due to transcriptional activation by MLL fusion protein that in turn causes global 5hmC gain [47]. Also, a TET1 knockout mouse had better survival illustrating oncogenic role of TET1 in MLL rearranged leukemias in contrast to its tumor suppressive activity. Additionally, Kroeze et al. showed an independent association of high levels of 5hmC with poor survival in AML [48]. The global increase of 5hmC in proneural glioblastoma promotes expression of oncogenes by increased TET1 expression [49]. High levels of 5hmC due to high TET1 & 3 expression in hypoxic BTIC predicts poor disease-free and overall survival in breast cancer [41]. Interestingly, Wang et al. showed a significant correlation between high 5hmC and cervical nodal metastasis in oral squamous cell carcinoma (OSCC) that is associated with poor overall survival [50]. This brings forth the view that the 5hmC levels can be variable within tumor sub-populations and can promote disease progression. Similarly, post-prostatectomy elevation in 5hmC of ERG-negative prostate cancer predicts biochemical recurrence of the disease [51].
Several studies have associated histone modifications with 5hmC content in cancer. H3K9me3 repressive histone mark was found with enriched 5hmC in renal cell carcinoma [53]. The enhancer regions of genes regulating oncogenic pathway marked with H3K4me1 gained 5hmC and significantly increased the respective gene expression in pancreatic cancer [54]. Increased 5hmC levels were observed in activating histone marks H3K27ac, H3K4me1 and H3K9me1 but were reduced in regions with H3K9me3 in cell-free circulating DNA of gastric, colorectal, pancreatic, liver and thyroid cancer [55]. Though a global loss of 5hmC and H3K27me3 marks are widely reported in cancers, a group of tumor cells in multiple cancers (prostate, breast, colon and pancreatic adenocarcinoma) showed high H3K27me3 modification and low 5hmC, whereas in normal tissues, both marks were strongly correlated [56].
Demethylating agents like 5-azacytidine, 2-deoxy 5-azacytidine (decitabine) are nucleoside analogs of cytidine, which are incorporated into DNA during S-phase that inhibit DNMT1 activity leading to global hypomethylation. These demethylating agents are commonly used in treatment of MDS, AML and other hematological cancers [57]. Decitabine increases 5hmC at hemimethylated regions by TET enzymes, while synergistic action of vitamin C and decitabine decreases proliferation and increases apoptosis upon triggering viral defense response by upregulating endogenous retroviral transcripts. Synergistic drug effect is reduced upon TET2 knockdown in A2780 cells, indicating a key function of 5hmC along with treatment with demethylating agents [58,59]. Targeted demethylation of cancer epigenome is a budding area of interest with the use of CRISPR-dcas9 TET1CD and specific guide RNA to demethylate target regions that increase target gene expression [60]. The use of CRISPR-dcas9 TET1CD with specific guide RNA upregulates BRCA1 by demethylation reducing proliferation and viability in HeLa and MCF7 cells [61].
Though loss of 5hmC is a frequent event in cancer, locus-specific changes are dynamic in different conditions. And hence it is reasonable to argue that deregulation of 5hmC in the genomic landscape can be a prime cause of cancer and also facilitates cancer progression. More findings are required to address on 5hmC locus specificity that will enable possible therapeutic aspects of 5hmC in cancer.
3. Effect of Cellular Factors on 5-Hydroxymethylcytosine
The 5hmC content of the genome depends primarily on expression and activity of TET, governed by various cofactors and inhibitors. Cofactors and co-substrates like 2-OG, Fe2+ and molecular oxygen (O2) are necessary to generate 5hmC (Figure 1). Mice injected with glucose, glutamate and glutamine had increased 2-OG and enhanced 5hmC levels in liver tissue especially at the gck (glucokinase) promoter [62]. PSAT1 (Phosphoserine Aminotransferase1) that produces high 2-OG, increases 5hmC, governing self-renewal and differentiation in mouse embryonic stem cells (mESC) [63]. Fe2+ is also an important modulator of TET determining global 5hmC levels. Chelation of Fe2+ by thiosemicarbazone (TSC24) reduced 5hmC and impaired murine development [64]. The secondary messenger—cyclic Adenylate MonoPhosphate (cAMP) was observed to regulate intracellular Fe2+ concentration to increase 5hmC, enabling cAMP controlled gene expression [65]. Post-translational modification of TET enzymes significantly determines their activity and 5hmC levels in cells. Acetylation, O-linked-N-acetylglucosamine (O-GlcNAc), phosphorylation, Poly-Adenosine Ribosylation (PARylation) and monoubiquitination are few known post-translational modifications of TET (Table 3). Acetylation of TET2 at K110 by p300 increases its stability by evading proteasomal degradation and more interestingly binds with DNMT1 and reduces hypermethylation under oxidative stress [66]. Monoubiquitination of TET1, 2 & 3 in K1212, K1299, K983 respectively by Cullin-RING ubiquitin ligase 4 (CRL4VprBP)E3 Ubiquitin ligase enhances the chromatin binding and increases 5hmC levels [67]. Similarly, covalent PARylation of TET1 has been shown to influence its activity and increase 5hmC while non-covalent binding reduces 5hmC by reducing DNA binding of TET1 [68]. TET enzymes are known to interact with the enzyme OGT (O-GlcNAc transferase) that plays a role in chromatin modulation, O-GlcNAcylation is possible in all three TET enzymes but O-GlcNAcylation of TET3 causes TET3 nuclear export and thus reduces 5hmC levels in the genome [69]. Phosphorylation of TET enzymes competes for site occupancy for O-GlcNAcylation hence may have opposing effect on TET enzymes [70].
MicroRNAs (miRNA) are short non-coding RNAs (ncRNAs) that repress gene expression by directly binding to the 3′UTR of target messenger RNA (mRNA) followed by RNA Induced Silencing Complex (RISC) mediated degradation. MicroRNAs targeting TET reduce its expression and therefore be considered as important factors in modulating 5hmC content. Few miRNAs like miR-26a can bind to TET1, 2 & 3 whereas miR-29b specifically targets TET1 and reduces 5hmC mediated demethylation during embryogenesis [71,72]. Overexpression of miRNAs against TET could be one of the possible mechanisms for reduced TET expression that is frequently observed in cancers. miR-22 that targets all three TET and reduces 5hmC levels especially in the loci of mir-200 favors cancer progression and metastasis in breast cancer [42]. Cheng et al reported more than 30 miRNAs that inhibit TET2 regulating hematopoiesis of wild-type AML [73]. TET1 targeting miRNAs like miR-520b, miR-494 in HCC and miR-191 are elevated in intrahepatic cholangiocarcinoma favoring proliferation, migration and invasion [74,75,76]. Besides, IDH and SDH-targeting miRNAs are also found to be upregulated in multiple cancers. Increased miR-181a in ovarian and liver cancer targets IDH1 [77]. Similarly, miR-183, miR-210 and miR-31 are found to target IDH2, SDHC and SDHD respectively (Table 4) [78,79,80]. However, a direct link between miRNA against IDH, SDH and FH and associating 5hmC levels are not yet reported. More studies on miRNA targeting IDH and SDH are warranted to elaborate 5hmC regulation in cancer.
4. Hypoxia-Induced 5-Hydroxymethylcytosine Changes Enable Cancer Progression
Hypoxia promotes tumor progression by favoring angiogenesis, epithelial-mesenchymal transition (EMT) and metastasis. Tumor hypoxia is observed to increase TET expression through Hypoxia Inducible Factor (HIF)–mediated transcriptional activation [41,84]. Apparently, TET enzymes assist hypoxia response program (HRP) by oxidizing repressive cytosine methylation of hypoxia-inducible genes (HIGs), since silencing TET1 reduced the 5hmC marks of HIGs in neuroblastoma cells [85]. Global 5hmC levels were found to be decreased in cancer cell lines like SHEP, SK-N-AS, N2A, H358 and Hep3B but surprisingly were increased in dedifferentiating N-type Neuroblastoma cell lines (SK-N-Be2c, SH-SY5Y, NBL-WN, La1-55n) and breast cancer cell line (4T1) [85,86]. Hence the role of 5hmC in determining cancer cell fate (metastatic/dedifferentiating), TET activity and levels of other 5mC variants under hypoxic conditions would help us understand and exploit epigenetic modulation for therapeutic, prognostic/ diagnostic applications.
Metabolites play substantial role in modulating 5hmC levels in tumor hypoxia. IDH which converts 2-OG to isocitrate essentially for lipid synthesis for cell growth and division [87]. It will be interesting to know if limiting levels of 2-OG can cause an eventual decrease in 5hmC. Intriguingly, gliomas with IDH1/2 mutation show reduced angiogenesis and low expression of HIG mRNA, adding evidence to prove the significant role of 5hmC in hypoxia [88]. In paraganglioma, a common cancer prevalent in hypoxic environments like high altitudes, SDH mutation is linked to poor survival suggesting possible involvement of 5hmC imbalance [89]. Increased cellular concentrations of oncometabolites like fumarate can also stabilize HIF1A, invoking changes in the pattern of gene expression which distinctly shows an additional layer of regulation by 5hmC and TET under hypoxic conditions [90].
Comprehensively, hypoxic condition increases TET expression and specifically marks HIG with 5hmC to facilitate HRP. TETs also function as transcriptional co-activators for some HIGs and different metabolites play a significant role in influencing HIGs expression patterns.
5. Heavy Metals Deregulate 5-Hydroxymethylcytosine
Heavy metals such as lead, mercury (Hg), arsenite (Ar3+), chromium (Cr) and nickel (Ni) are known to cause various types of cancers, but the underlying mechanism of carcinogenesis is largely unknown. DNA binding proteins possess zinc finger motif to facilitate its binding. The functions of metal binding proteins are altered by heavy metals, affecting mainly the zinc finger proteins [91,92]. TET contains highly conserved zinc finger motifs that are critical for both catalytic activity and binding to CpG islands of DNA. Heavy metals may therefore be speculated to hinder 5hmC balance by inhibiting zinc finger motif of TET and modulating TCA cycle enzymes.
Arsenite has been implicated to cause cancers of skin, lung and bladder [93]. Arsenite induces aberrant DNA methylation loss by depriving S-Adenosyl Methionine (SAM) induced epigenetic reprogramming [94]. Recently, it has been reported that Ar3+ can potentially inhibit TET activity and reduce the levels of 5hmC, which could be another underlying mechanism of Ar3+ induced carcinogenesis [95]. Similarly, Ni has been shown to cause hypermethylation in TSGs like P53 of Chinese hamster cell line by facilitating the increased binding and activity of DNMT1 [96]. Alternatively, Yin et al showed that Ni can inhibit the activity of TET enzymes by displacing Fe(II), a possible mode of hypermethylating TSG [97].
Cadmium (Cd) upon acute exposure reduces methylation but prolonged exposure hypermethylates the TSG regions like Ras Association Domain Family Member 1 (RASSF1) and p16 by DNMT overexpression. However, the effect of Cd in carcinogenesis has not been studied in relation to DNA hydroxymethylation, which would enable a better understanding of Cd exposed carcinogenesis [98]. A similar effect of heavy metals on IDH, SDH and FH inhibition may throw light on alternate 5hmC deregulation mode of carcinogenesis.
6. Oxidative Stress on 5-Hydroxymethylcytosine
Increased ROS levels in cancer cells are often observed with cancer metastasis and drug resistance [99]. Recently, 5-Fluro Uracil induced ROS was found to elevate TET1 expression that demethylates Nrf2 gene promotors enabling resistance and survival in colorectal cancer cells, SNU-C5R [52]. Though ROS can increase 5mC oxidation via 5hmC which would eventually demethylate and the 8-oxo-guanosine formed by oxidative damage in DNA impairs DNMT activity leading to global hypomethylation. Alternately, expression and activity of DNMT, increased by ROS, may account for regional hypermethylation and both the phenomena are commonly witnessed in various cancers [100]. It would be hence interesting to understand the role of ROS mediated 5mC/5hmC imbalance in carcinogenic events.
7. Carcinogens Deregulate 5-Hydroxymethylcytosine
Potent carcinogens have notable effect on hydroxymethylation of cytosine. Phenobarbital a non-genotoxic carcinogen was found to enrich 5hmC levels of Chemokine receptor (Cxcr7) and WNT1 inducible signaling pathway protein 1 (Wisp1), whereas loss of N-myc downstream regulated 1 (Ndrg1) that are previously linked to malignant transformation and poor prognosis in HCC [101]. Adult mice exposed to diethylstilbestrol non-steroidal estrogen resulted with uterine and vaginal cancer shows a long-term effect with prolonged 5hmC alteration by reduced TET1 expression [102]. Similarly, rats induced with genotoxic carcinogens like riddelliine and aristolochic acid showed a decrease in 5hmC levels and accompanied TET2 down-regulation in liver at pre-neoplastic transformation state [103]. Contrastingly, hydroquinone is shown to increase 5hmC levels and TET1 expression via increase in ROS production [104]. The role of these carcinogens in DNA damage and mutagenesis has been well documented but their role in altering 5hmC levels are to be elaborated more in cancer context.
8. Nutrients Enable 5-Hydroxymethylcytosine Regulation
Nutrients that potentially prevent pathogenesis are also crucial components of epigenetic maintenance and modifications [105]. Vitamin C (ascorbate) is well known and highly correlated with reduced risk of cancer [105,106]. The antioxidant property of vitamin C reduces ROS stress in cells but surprisingly increases activity of TET enzymes by aiding quick exchange of Fe2+ for subsequent catalysis [107,108]. Redox-active quinones are anti-oxidant rich natural products shown to increase the 5hmC levels in a TET-dependent manner [109]. Among trace elements, zinc is associated with reduced cancer risk and essential nutrient for immunity and anti-inflammatory processes [110,111]. Intriguingly, zinc can inhibit Oxoglutarate Dehydrogenase (OGDH) that converts 2-OG to succinyl CoA [112]. Although, there is no substantial evidence on OGDH inhibition and 5hmC increase, increase of SDH activity in order to clear succinate that may have an effect on 5hmC remains so far unexplained.
9. 5-Hydroxymethylcytosine Changes are Locus-Specific
Though a substantial loss of 5hmC was observed and linked to cancer, 5hmC changes are dynamic at particular conditions or upon receipt of certain stimulus enabling cellular response (Table 5). DNMT and TET can only recognize CpG and mCpG regions of DNA respectively, but the mechanism of locus-specific enrichment is poorly understood. Identifying such mechanisms would open up new avenues in therapeutic strategies. Possible mechanisms of locus-specific 5hmC changes are guided by factors like transcription factors (TF), TET binding proteins, Polycomb Repressive complex 2 (PRC2), histone modifications and post-translational modifications of TFs (Figure 2). TFs like HIF1, thyroid hormone nuclear receptors, retinoic acid receptor (RAR), etc., are shown to interact with TET1 and thus demethylate their target genes by 5hmC [84,113,114,115]. Apart from TFs, TET-interacting proteins can also recruit it to a particular locus. For instance, Growth Arrest And DNA Damage Inducible Alpha (GADD45A) is a demethylating complex protein that recruits TET to promoter of TCF21, a tumor suppressor found to be hypermethylated in cancer cell lines [116]. PRC2, which is involved in repression of gene expression through Enhancer of Zeste Homolog 2 (EZH2), specifically recruits TET1 by Suppressor of Zeste 12 Protein Homolog (SUZ12) to the H3K27 tri-methylated locus and enriches 5hmC. This enables a bivalent chromatin and poised for expression upon signal, which is highly observed in ESCs and tends to lose 5hmC marks upon PRC2 depletion in cancer cell lines [56,117]. Histone modifications like H3K4me1 and H3K27ac enriches 5hmC at enhancer regions correlated with gene expression and H3K36me3 modifications specifically gained 5hmC in intronic regions [118,119]. TET1 recognizes the post-translational modification PARylation of Peroxisome proliferator-activated receptor γ (PPARγ) and enriches 5hmC in the regulatory regions of PPARγ regulated genes and enables transcription by demethylation, similarly PARylation of TET1 stimulates its activity and may enable demethylation through PAR binding of TF by recruiting TET enzymes to their loci [120]. These findings suggest that specifying the locus of 5hmC modification itself is a complex process, which dynamically changes due to environmental/cellular stimuli.
10. Conclusions
This review elaborates the role of 5hmC in cancer with its regulation in genomic context by various factors. These factors are crucial as they can directly or indirectly determine 5hmC levels enabling carcinogenic events and cancer progression. Convincing evidence on loss of global 5hmC levels in cancers and identification of locus-specific changes of 5hmC in particular will lead to a better understanding of its selective mechanism in carcinogenesis and development of novel drug targets. Rather than a demethylation intermediate, 5hmC has emerged as a critical DNA modification agent involved in altered gene expression, DNA repair activity and maintenance of genomic integrity establishing its versatile role in cancer progression. Use of 5hmC in cancer treatment is at an early stage. Already available demethylating agents like decitabine are known to increase 5hmC levels and enable demethylation in hemimethylated regions. However, the action of these hypomethylating agents is non-specific from a genomic context. Drugs that can specifically demethylate particular loci in cancer would be of immense help. Exploitation of genome editing to target specific demethylation could be a therapeutic strategy of locus-specific demethylation in the future. Thus, identifying mechanisms of locus-specific 5hmC changes and loci that are differently hydroxymethylated in cancer will enable us to identify biomarkers and design improved therapeutic strategies.
Acknowledgments
Our work is supported by a grant (EMR/2015/001319) from Department of Science and Technology, India. We acknowledge Priya Ramanathan for critical comments and proofreading the manuscript.
Author Contributions
S.M. provided the theme and concept for the review; V.R.A. and A.K.D.M.R. wrote the manuscript. S.M. and T.R. evaluated the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, N.; Burns, D.M.; Blau, H.M. DNA demethylation dynamics. Cell 2011, 146, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Nestor, C.E.; Ottaviano, R.; Reddington, J.; Sproul, D.; Reinhardt, D.; Dunican, D.; Katz, E.; Dixon, M.J.; Harrison, D.J.; Meehan, R.R. Tissue type is a major modifier of the 5-hydroxymethylcytosine content of human genes. Genome Res. 2012, 22, 467–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münzel, M.; Globisch, D.; Carell, T. 5-hydroxymethylcytosine, the sixth base of the genome. Angew. Chem. Int. Ed. 2011, 50, 6460–6468. [Google Scholar]
- Pastor, W.A.; Aravind, L.; Rao, A. Tetonic shift: Biological roles of TET proteins in DNA demethylation and transcription. Nat. Rev. Mol. Cell Biol. 2013, 14, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Kriaucionis, S.; Tahiliani, M. Expanding the epigenetic landscape: Novel modifications of cytosine in genomic DNA. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Shen, L.; Dai, Q.; Wu, S.C.; Collins, L.B. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 2011, 333, 1300–1303. [Google Scholar] [CrossRef] [PubMed]
- Mohr, F.; Döhner, K.; Buske, C.; Rawat, V. Tet genes: New players in DNA demethylation and important determinants for stemness. Exp. Hematol. 2011, 39, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Kinney, S.; Chin, H.; Vaisvila, R.; Bitinaite, J.; Zheng, Y.; Estève, P.-O.; Feng, S.; Stroud, H.; Jacobsen, S.E.; Pradhan, S. Tissue-specific distribution and dynamic changes of 5-hydroxymethylcytosine in mammalian genomes. J. Biol. Chem. 2011, 286, 24685–24693. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, M. Distribution of 5-hydroxymethylcytosine in different human tissues. J. Nucleic Acids 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Iurlaro, M.; Ficz, G.; Oxley, D.; Raiber, E.-A.; Bachman, M.; Booth, M.J.; Andrews, S.; Balasubramanian, S.; Reik, W. A screen for hydroxymethylcytosine and formylcytosine binding proteins suggests functions in transcription and chromatin regulation. Genome Biol. 2013, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Spruijt, C.G.; Gnerlich, F.; Smits, A.H.; Pfaffeneder, T.; Jansen, P.; Bauer, C.; Münzel, M.; Wagner, M.; Müller, M.; Khan, F.; et al. Dynamic readers for 5-(hydroxy)methylcytosine and its oxidized derivatives. Cell 2013, 152, 1146–1159. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Liu, Y.; Upadhyay, A.K.; Chang, Y.; Howerton, S.B.; Vertino, P.M.; Zhang, X.; Cheng, X. Recognition and potential mechanisms for replication and erasure of cytosine hydroxymethylation. Nucleic Acids Res. 2012, 40, 4841–4849. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Lin, K.; Song, J.; Wang, Y. Effects of TET-induced oxidation products of 5-methylcytosine on Dnmt1- and DNMT3a-mediated cytosine methylation. Mol. BioSyst. 2014, 10, 1749–1752. [Google Scholar] [CrossRef] [PubMed]
- Bachman, M.; Uribe-Lewis, S.; Yang, X.; Williams, M.; Murrell, A.; Balasubramanian, S. 5-hydroxymethylcytosine is a predominantly stable DNA modification. Nat. Chem. 2014, 6, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, K.D.; Helin, K. Role of TET enzymes in DNA methylation, development, and cancer. Genes Dev. 2016, 30, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, G.P.; Xiong, W.; Hahn, M.A.; Jin, S.-G. The role of 5-hydroxymethylcytosine in human cancer. Cell Tissue Res. 2014, 356, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Losman, J.A.; Kaelin, W.G. What a difference a hydroxyl makes: Mutant IDH, (R)-2-hydroxyglutarate, and cancer. Genes Dev. 2013, 27, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Bardella, C.; Pollard, P.J.; Tomlinson, I. SDH mutations in cancer. Biochim. Biophys. Acta Bioenerg. 2011, 1807, 1432–1443. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Selak, M.A.; Gottlieb, E. Succinate dehydrogenase and fumarate hydratase: Linking mitochondrial dysfunction and cancer. Oncogene 2006, 25, 4675–4682. [Google Scholar] [CrossRef] [PubMed]
- Kroeze, L.I.; van der Reijden, B.A.; Jansen, J.H. 5-hydroxymethylcytosine: An epigenetic mark frequently deregulated in cancer. Biochim. Biophys. Acta Rev. Cancer 2015, 1855, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Lan, L. 5-hydroxymethylcytosine: A new insight into epigenetics in cancer. Cancer Biol. Ther. 2014, 15, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Mariani, C.J.; Madzo, J.; Moen, E.L.; Yesilkanal, A.; Godley, L.A. Alterations of 5-hydroxymethylcytosine in human cancers. Cancers 2013, 5, 786–814. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Guo, Z.; Ma, Q.; Xu, Z.; Zhou, Y.; Xu, Z.; Li, Z.; Liu, Y.; Ye, X.; et al. Loss of 5-hydroxymethylcytosine is linked to gene body hypermethylation in kidney cancer. Cell Res. 2016, 26, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liu, Y.; Zhang, Z.; Li, J.; Wan, Y.; Zhang, L.; Wang, Y.; Li, X.; Xu, Y.; Fu, X.; et al. 5-hydroxymethylcytosine loss is associated with poor prognosis for patients with WHO grade II diffuse astrocytomas. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Yu, Y.; Luo, M.; Zhang, Z.; Shi, S.; Feng, X.; Chen, Z.; He, J. Loss of 5-hydroxymethylcytosine is an independent unfavorable prognostic factor for esophageal squamous cell carcinoma. PLoS ONE 2016, 11, e0153100. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wu, K.; Ji, M.; Jin, W.; He, N.; Shi, B.; Hou, P. Decreased 5-hydroxymethylcytosine (5-hmC) is an independent poor prognostic factor in gastric cancer patients. J. Biomed. Nanotechnol. 2013, 9, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, L.; Chen, X.; Shen, J.; Shan, J.; Xu, Y.; Yang, Z.; Wu, L.; Xia, F.; Bie, P.; et al. Decrease of 5-hydroxymethylcytosine is associated with progression of hepatocellular carcinoma through downregulation of TET1. PLoS ONE 2013, 8, e62828. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.-W.; Li, G.-C.; Chen, C.-H.; Yeh, M.-H.; Huang, J.-S.; Tseng, H.-H.; Fu, T.-Y.; Liou, H.-H.; Pan, H.-W.; Huang, S.-F.; et al. Reduction of global 5-hydroxymethylcytosine is a poor prognostic factor in breast cancer patients, especially for an ER/PR-negative subtype. Breast Cancer Res. Treat. 2015, 153, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, K.; Shao, Y.; Sui, F.; Yang, Q.; Shi, B.; Hou, P.; Ji, M. Decreased 5-hydroxymethylcytosine (5-hmC) predicts poor prognosis in early-stage laryngeal squamous cell carcinoma. Am. J. Cancer Res. 2016, 6, 1089. [Google Scholar] [PubMed]
- Delhommeau, F.; Dupont, S.; Valle, V.; James, C.; Trannoy, S.; Massé, A.; Kosmider, O.; Couedic, J.-P.; Robert, F.; Alberdi, A.; et al. Mutation in TET2 in myeloid cancers. N. Engl. J. Med. 2009, 360, 2289–2301. [Google Scholar] [CrossRef] [PubMed]
- Reitman, Z.J.; Yan, H. Isocitrate dehydrogenase 1 and 2 mutations in cancer: Alterations at a crossroads of cellular metabolism. J. Natl. Cancer Inst. 2010, 102, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Sciacovelli, M.; Frezza, C. Oncometabolites: Unconventional triggers of oncogenic signalling cascades. Free Radic. Biol. Med. 2016, 100, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, S.; Gottlieb, E. Oncometabolites: Tailoring our genes. FEBS J. 2015, 282, 2796–2805. [Google Scholar] [CrossRef] [PubMed]
- Ghiam, A.F.; Cairns, R.A.; Thoms, J.; Pra, D.A.; Oncogene, A.-O. IDH mutation status in prostate cancer. Oncogene 2012, 31, 3826. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Dong, Q.; Zhang, C.; Kuan, P.F.; Liu, Y.; Oncogene, J.-W.R. Mutations in isocitrate dehydrogenase 1 and 2 occur frequently in intrahepatic cholangiocarcinomas and share hypermethylation targets with glioblastomas. Oncogene 2013, 32, 3091–3100. [Google Scholar] [CrossRef] [PubMed]
- Killian, J.K.; Kim, S.Y.; Miettinen, M.; Smith, C.; Merino, M.; Tsokos, M.; Quezado, M.; Smith, W.I.; Jahromi, M.S.; Xekouki, P.; et al. Succinate dehydrogenase mutation underlies global epigenomic divergence in gastrointestinal stromal tumor. Cancer Discov. 2013, 3, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, C.J.; Shuch, B.; Vocke, C.D.; Metwalli, A.R.; Bratslavsky, G.; Middelton, L.; Yang, Y.; Wei, M.H.; Pautler, S.E.; Peterson, J.; et al. Succinate dehydrogenase kidney cancer: An aggressive example of the warburg effect in cancer. J. Urol. 2012, 188, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, H.J.; Kiuru, M.; Ylisaukko-oja, S.K.; Salovaara, R.; Herva, R.; Koivisto, P.A.; Vierimaa, O.; Aittomäki, K.; Pukkala, E.; Launonen, V.; et al. Increased risk of cancer in patients with fumarate hydratase germline mutation. J. Med. Genet. 2006, 43, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Song, C.-X.; Huang, H.; Frankenberger, C.A.; Sankarasharma, D.; Gomes, S.; Chen, P.; Chen, J.; Chada, K.K.; He, C.; et al. HMGA2/TET1/HOXA9 signaling pathway regulates breast cancer growth and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 9920–9925. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-Z.; Chen, S.-F.; Nieh, S.; Benner, C.; Ger, L.-P.; Jan, C.-I.; Ma, L.; Chen, C.-H.; Hishida, T.; Chang, H.-T.; et al. Hypoxia drives breast tumor malignancy through a TET-TNFα-p38-MAPK signaling axis. Cancer Res. 2015, 75, 3912–3924. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Poliseno, L.; Song, M.; Ala, U.; Webster, K.; Ng, C.; Beringer, G.; Brikbak, N.J.; Yuan, X.; Cantley, L.C.; et al. microRNA-antagonism regulates breast cancer stemness and metastasis via TET-family-dependent chromatin remodeling. Cell 2013, 154, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Matthias, W.; Liou, W.; Pulverer, W.; Singer, C.F.; Rappaport-Fuerhauser, C.; Kandioler, D.; Egger, G.; Weinhäusel, A. Cytosine 5-hydroxymethylation of the LZTS1 gene is reduced in breast cancer. Transl. Oncol. 2013, 6, 715–721. [Google Scholar] [CrossRef]
- Kamiya, T.; Nakahara, R.; Mori, N.; Hara, H.; Adachi, T. Ten-eleven translocation 1 functions as a mediator of SOD3 expression in human lung cancer A549 cells. Free Radic. Res. 2017, 51, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Chapman, C.G.; Mariani, C.J.; Wu, F.; Meckel, K.; Butun, F.; Chuang, A.; Madzo, J.; Bissonnette, M.B.; Kwon, J.H.; Godley, L.A. Tet-catalyzed 5-hydroxymethylcytosine regulates gene expression in differentiating colonocytes and colon cancer. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Neri, F.; Dettori, D.; Incarnato, D.; Krepelova, A.; Rapelli, S.; Maldotti, M.; Parlato, C.; Paliogiannis, P.; Oliviero, S. TET1 is a tumour suppressor that inhibits colon cancer growth by derepressing inhibitors of the wnt pathway. Oncogene 2014, 34, 4168–4176. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Jiang, X.; Li, Z.; Li, Y.; Song, C.-X.; He, C.; Sun, M.; Chen, P.; Gurbuxani, S.; Wang, J.; et al. TET1 plays an essential oncogenic role in MLL-rearranged leukemia. Proc. Natl. Acad. Sci. USA 2013, 110, 11994–11999. [Google Scholar] [CrossRef] [PubMed]
- Kroeze, L.I.; Aslanyan, M.G.; van Rooij, A.; Koorenhof-Scheele, T.N.; Massop, M.; Carell, T.; Boezeman, J.B.; Marie, J.-P.; Halkes, C.J.M.; de Witte, T.; et al. Characterization of acute myeloid leukemia based on levels of global hydroxymethylation. Blood 2014, 124, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Takai, H.; Masuda, K.; Sato, T.; Sakaguchi, Y.; Suzuki, T.; Suzuki, T.; Koyama-Nasu, R.; Nasu-Nishimura, Y.; Katou, Y.; Ogawa, H.; et al. 5-Hydroxymethylcytosine plays a critical role in glioblastomagenesis by recruiting the CHTOP-methylosome complex. Cell Rep. 2014, 9, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, H.; Wang, Q.; Li, Z.; Zhu, Y.; Zhang, W.; Wang, Y.; Jiang, H.; Cheng, J. The level and clinical significance of 5-hydroxymethylcytosine in oral squamous cell carcinoma: An immunohistochemical study in 95 patients. Pathol. Res. Pract. 2017, 213, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Strand, S.H.; Hoyer, S.; Lynnerup, A.-S.; Haldrup, C.; Storebjerg, T.; Borre, M.; Orntoft, T.F.; Sorensen, K.D. High levels of 5-hydroxymethylcytosine (5hmC) is an adverse predictor of biochemical recurrence after prostatectomy in ERG-negative prostate cancer. Clin. Epigenet. 2015, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Piao, M.J.; Kim, K.C.; Kang, H.K.; Chang, W.Y.; Park, I.C.; Keum, Y.S.; Surh, Y.J.; Hyun, J.W. Epigenetic modification of Nrf2 in 5-fluorouracil-resistant colon cancer cells: Involvement of TET-dependent DNA demethylation. Cell Death Dis. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Lautenschlaeger, T.; Frankhouser, D.; Ye, Z.; Huebner, A.; Jin, V.; Yan, P.; Chakravarti, A. Abstract 2963: 5-hydroxymethylcytosine alterations at H3K9me3 marked genomic regions serve as potential biomarker for renal cell carcinoma patients. Cancer Res. 2015, 75. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Pradhan, K.; Campbell, N.; Mazdo, J.; Vasantkumar, A.; Maqbool, S.; Bhagat, T.D.; Gupta, S.; Suzuki, M.; Yu, Y.; et al. Altered hydroxymethylation is seen at regulatory regions in pancreatic cancer and regulates oncogenic pathways. Genome Res. 2017, 27, 1830–1842. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, X.; Lu, X.; You, L.; Song, Y.; Luo, Z.; Zhang, J.; Nie, J.; Zheng, W.; Xu, D.; et al. DNA 5-hydroxymethylcytosines from cell-free circulating DNA as diagnostic biomarkers for human cancers. bioRxiv 2017. [Google Scholar] [CrossRef]
- Haffner, M.; Pellakuru, L.; Ghosh, S.; Lotan, T.; Nelson, W.G.; Marzo, A.; Yegnasubramanian, S. Tight correlation of 5-hydroxymethylcytosine and polycomb marks in health and disease. Cell Cycle 2013, 12, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Issa, J.-P.J.; Baylin, S. Targeting the cancer epigenome for therapy. Nat. Rev. Genet. 2016, 17, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, B.; McGovern, A.; Cui, Y.; Choudhury, S.; Cho, I.-H.; Cooper, B.; Chevassut, T.; Lossie, A.C.; Irudayaraj, J. The hypomethylating agent decitabine causes a paradoxical increase in 5-hydroxymethylcytosine in human leukemia cells. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ohtani, H.; Zhou, W.; Ørskov, A.D.; Charlet, J.; Zhang, Y.W.; Shen, H.; Baylin, S.B.; Liang, G.; Grønbæk, K.; et al. Vitamin C increases viral mimicry induced by 5-aza-2′-deoxycytidine. Proc. Natl. Acad. Sci. USA 2016, 113, 10238–10244. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Tao, Y.; Gao, X.; Zhang, L.; Li, X.; Zou, W.; Ruan, K.; Wang, F.; Xu, G.-L.; Hu, R. A CRISPR-based approach for targeted DNA demethylation. Cell Discov. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.R.; Cui, Y.; Lubecka, K.; Stefanska, B.; Irudayaraj, J. CRISPR-dCas9 mediated TET1 targeting for selective DNA demethylation at BRCA1 promoter. Oncotarget 2016, 7, 46545–46556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.W.; Wang, Z.; Xie, W.; Cai, Y.; Xia, L.; Easwaran, H.; Luo, J.; Yen, R.-W.; Li, Y.; Baylin, S.B. Acetylation enhances TET2 function in protecting against abnormal DNA methylation during oxidative stress. Mol. Cell 2017, 65, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Lv, L.; Nakagawa, M.; Yu, Y.; Yu, C.; D’Alessio, A.C.; Nakayama, K.; Fan, H.-Y.; Chen, X.; Xiong, Y. CRL4(VprBP) E3 ligase promotes monoubiquitylation and chromatin binding of TET dioxygenases. Mol. Cell 2015, 57, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Ciccarone, F.; Valentini, E.; Zampieri, M.; Caiafa, P. 5mC-hydroxylase activity is influenced by the PARylation of TET1 enzyme. Oncotarget 2015, 6, 24333–24347. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, X.; Gao, W.; Li, P.; Hou, J.; Li, J.; Wong, J. Differential regulation of the ten-eleven translocation (TET) family of dioxygenases by O-linked β-N-acetylglucosamine transferase (OGT). J. Biol. Chem. 2014, 289, 5986–5996. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lin, H.; Xu, H.; Zhang, L.; Cheng, L.; Wen, B.; Shou, J.; Guan, K.; Xiong, Y.; Ye, D. TET-catalyzed 5-methylcytosine hydroxylation is dynamically regulated by metabolites. Cell Res. 2014, 24, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.-Y.; Kwak, S.; Lee, S.; Kim, H.; Lee, S.; Kim, J.-H.; Kim, Y.; Jeon, Y.; Chung, D.; Jin, X.; et al. Psat1-dependent fluctuations in α-ketoglutarate affect the timing of ESC differentiation. Cell Metab. 2016, 24, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-H.; Liang, S.; Guo, J.; Choi, J.-W.; Kim, N.-H.; Lu, W.-F.; Cui, X.-S. Analysis of ferrous on ten-eleven translocation activity and epigenetic modifications of early mouse embryos by fluorescence microscopy. Microsc. Microanal. 2016, 22, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Camarena, V.; Sant, D.W.; Huff, T.; Mustafi, S.; Muir, R.K.; Renslo, A.R.; Monje, P.; Wang, G. cAMP signaling regulates DNA demethylation by augmenting the intracellular labile ferrous iron pool. bioRxiv 2017. [Google Scholar] [CrossRef]
- Bauer, C.; Göbel, K.; Nagaraj, N.; Colantuoni, C.; Wang, M.; Müller, U.; Kremmer, E.; Rottach, A.; Leonhardt, H. Phosphorylation of TET proteins is regulated via O-glcnacylation by the O-linked N-acetylglucosamine transferase (OGT). J. Biol. Chem. 2015, 290, 4801–4812. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Ng, S.H.; Luk, A.C.; Liao, J.; Jiang, X.; Feng, B.; Lun Mak, K.K.; Rennert, O.M.; Chan, W.Y.; Lee, T.L. MicroRNA-29b/Tet1 regulatory axis epigenetically modulates mesendoderm differentiation in mouse embryonic stem cells. Nucleic Acids Res. 2015, 43, 7805–7822. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Jin, L.; Wang, X.; Luo, A.; Hu, J.; Zheng, X.; Tsark, W.M.; Riggs, A.D.; Ku, H.T.; Huang, W. MicroRNA-26a targets ten eleven translocation enzymes and is regulated during pancreatic cell differentiation. Proc. Natl. Acad. Sci. USA 2013, 110, 17892–17897. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Guo, S.; Chen, S.; Mastriano, S.J.; Liu, C.; D’Alessio, A.C.; Hysolli, E.; Guo, Y.; Yao, H.; Megyola, C.M.; et al. An extensive network of TET2-targeting microRNAs regulates malignant hematopoiesis. Cell Rep. 2013, 5, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lu, Z.; Gao, Y.; Ye, L.; Song, T.; Zhang, X. MiR-520b suppresses proliferation of hepatoma cells through targeting ten-eleven translocation 1 (TET1) mRNA. Biochem. Biophys. Res. Commun. 2015, 460, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Chuang, K.-H.H.; Whitney-Miller, C.L.; Chu, C.-Y.Y.; Zhou, Z.; Dokus, M.K.; Schmit, S.; Barry, C.T. MicroRNA-494 is a master epigenetic regulator of multiple invasion-suppressor microRNAs by targeting ten eleven translocation 1 in invasive human hepatocellular carcinoma tumors. Hepatology 2015, 62, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, Z.Q.; Yang, Z.R.; Tong, D.N.; Guan, J.; Shi, B.J.; Nie, J.; Ding, X.T.; Li, B.; Zhou, G.W.; et al. MicroRNA-191 acts as a tumor promoter by modulating the TET1-p53 pathway in intrahepatic cholangiocarcinoma. Hepatology 2017, 66, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.; Lee, C.; Joseph, P.; Marchini, S.; Baccarini, A.; Kolev, V.; Romualdi, C.; Fruscio, R.; Shah, H.; Wang, F.; et al. MicroRNA-181a has a critical role in ovarian cancer progression through the regulation of the epithelial-mesenchymal transition. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Puissegur, M.P.; Mazure, N.M.; Bertero, T.; Pradelli, L.; Grosso, S.; Robbe-Sermesant, K.; Maurin, T.; Lebrigand, K.; Cardinaud, B.; Hofman, V.; et al. miR-210 is overexpressed in late stages of lung cancer and mediates mitochondrial alterations associated with modulation of HIF-1 activity. Cell Death Differ. 2011, 18, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Sasayama, T.; Tanaka, K.; Nakamizo, S.; Nishihara, M.; Mizukawa, K.; Kohta, M.; Koyama, J.; Miyake, S.; Taniguchi, M.; et al. MicroRNA-183 upregulates HIF-1α By targeting isocitrate dehydrogenase 2 (IDH2) in glioma cells. J. Neurooncol. 2013, 111, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, M.D.; Boyd, K.L.; Moyo, T.; Mitra, R.; Duszynski, R.; Arrate, M.P.; Chen, X.; Zhao, Z.; Blackwell, T.S.; Andl, T.; et al. MicroRNA-31 initiates lung tumorigenesis and promotes mutant KRAS-driven lung cancer. J. Clin. Investig. 2016, 126, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Ito, K.; Ala, U.; Kats, L.; Webster, K.; Sun, S.; Jongen-Lavrencic, M.; Manova-Todorova, K.; Teruya-Feldstein, J.; Avigan, D.E.; et al. The oncogenic microRNA miR-22 targets the TET2 tumor suppressor to promote hematopoietic stem cell self-renewal and transformation. Cell Stem Cell 2013, 13, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yin, D.; Zhang, Y.; Yu, L.; Li, X.-D.; Zhou, Z.-J.; Zhou, S.-L.; Gao, D.-M.; Hu, J.; Jin, C.; et al. MicroRNA-29a induces loss of 5-hydroxymethylcytosine and promotes metastasis of hepatocellular carcinoma through a TET-SOCS1-MMP9 signaling axis. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, H.; Liu, Y.; Zhang, J.; Li, H.; Liu, W.; Cao, G.; Xv, P.; Zhang, J.; Lv, C.; et al. Mir-30a as potential therapeutics by targeting TET1 through regulation of drp-1 promoter hydroxymethylation in idiopathic pulmonary fibrosis. Int. J. Mol. Sci. 2017, 18, 633. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-P.; Chen, H.-F.; Chen, S.-Y.; Cheng, W.-C.; Wang, H.-W.; Shen, Z.-J.; Song, C.; Teng, S.-C.; He, C.; Wu, K.-J. TET1 regulates hypoxia-induced epithelial-mesenchymal transition by acting as a co-activator. Genome Biol. 2014, 15, 513. [Google Scholar] [CrossRef] [PubMed]
- Mariani, C.J.; Vasanthakumar, A.; Madzo, J.; Yesilkanal, A.; Bhagat, T.; Yu, Y.; Bhattacharyya, S.; Wenger, R.H.; Cohn, S.L.; Nanduri, J.; et al. TET1-mediated hydroxymethylation facilitates hypoxic gene induction in neuroblastoma. Cell Rep. 2014, 7, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Thienpont, B.; Steinbacher, J.; Zhao, H.; D’Anna, F.; Kuchnio, A.; Ploumakis, A.; Ghesquière, B.; Dyck, L.; Boeckx, B.; Schoonjans, L.; et al. Tumour hypoxia causes DNA hypermethylation by reducing TET activity. Nature 2016, 537, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Wise, D.R.; Ward, P.S.; Shay, J.E.S.; Cross, J.R.; Gruber, J.J.; Sachdeva, U.M.; Platt, J.M.; DeMatteo, R.G.; Simon, C.M.; Thompson, C.B. Hypoxia promotes isocitrate dehydrogenase-dependent carboxylation of α-ketoglutarate to citrate to support cell growth and viability. Proc. Natl. Acad. Sci. USA 2011, 108, 19611–19616. [Google Scholar] [CrossRef] [PubMed]
- Kickingereder, P.; Sahm, F.; Radbruch, A.; Wick, W.; Heiland, S.; von Deimling, A.; Bendszus, M.; Wiestler, B. IDH mutation status is associated with a distinct hypoxia/angiogenesis transcriptome signature which is non-invasively predictable with rCBV imaging in human glioma. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Her, Y.F.; Nelson-Holte, M.; Maher, L. Oxygen concentration controls epigenetic effects in models of familial paraganglioma. PLoS ONE 2015, 10, e0127471. [Google Scholar] [CrossRef] [PubMed]
- Laukka, T.; Mariani, C.J.; Ihantola, T.; Cao, J.Z.; Hokkanen, J.; Kaelin, W.G.; Godley, L.A.; Koivunen, P. Fumarate and succinate regulate expression of hypoxia-inducible genes via TET enzymes. J. Biol. Chem. 2015, 291, 4256–4265. [Google Scholar] [CrossRef] [PubMed]
- Tamás, M.J.; Sharma, S.K.; Ibstedt, S.; Jacobson, T.; Christen, P. Heavy metals and metalloids as a cause for protein misfolding and aggregation. Biomolecules 2014, 4, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, A. Zinc finger proteins as potential targets for toxic metal ions: Differential effects on structure and function. Antioxid. Redox Signal. 2001, 3, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.D.; Vucic, E.A.; Becker-Santos, D.D.; Gil, L.; Lam, W.L. Arsenic exposure and the induction of human cancers. J. Toxicol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Reichard, J.F.; Puga, A. Effects of arsenic exposure on DNA methylation and epigenetic gene regulation. Epigenomics 2010, 2, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jiang, J.; Li, L.; Amato, N.J.; Wang, Z.; Wang, Y. Arsenite targets the zinc finger domains of TET proteins and inhibits TET-mediated oxidation of 5-methylcytosine. Environ. Sci. Technol. 2015, 49, 11923–11931. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.W.; Klein, C.B.; Kargacin, B.; Salnikow, K.; Kitahara, J.; Dowjat, K.; Zhitkovich, A.; Christie, N.T.; Costa, M. Carcinogenic nickel silences gene expression by chromatin condensation and DNA methylation: A new model for epigenetic carcinogens. Mol. Cell. Biol. 1995, 15, 2547–2557. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Mo, J.; Dai, J.; Wang, H. Nickel(II) inhibits TET-mediated 5-methylcytosine oxidation by high affinity displacement of the cofactor iron(II). ACS Chem. Biol. 2017, 12, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- Arita, A.; Costa, M. Epigenetics in metal carcinogenesis: Nickel, arsenic, chromium and cadmium. Metall. Integr. Biomet. Sci. 2009, 1, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Waris, G.; Ahsan, H. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog. 2006, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Ni, X. Ros-mediated DNA methylation pattern alterations in carcinogenesis. Curr. Drug Targets 2015, 16, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.P.; Hunter, J.M.; Lempiäinen, H.; Müller, A.; Terranova, R.; Moggs, J.G.; Meehan, R.R. Dynamic changes in 5-hydroxymethylation signatures underpin early and late events in drug exposed liver. Nucleic Acids Res. 2013, 41, 5639–5654. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, W.N.; Chevalier, D.M.; Phelps, J.Y.; Cantor, A.M.; Padilla-Banks, E.; Newbold, R.R.; Archer, T.K.; Kinyamu, K.H.; Williams, C.J. Persistently altered epigenetic marks in the mouse uterus after neonatal estrogen exposure. Mol. Endocrinol. 2013, 27, 1666–1677. [Google Scholar] [CrossRef] [PubMed]
- Lian, C.; Xu, S.; Guo, W.; Yan, J.; Frank, M.; Liu, R.; Liu, C.; Chen, Y.; Murphy, G.F.; Chen, T. Decrease of 5-hydroxymethylcytosine in rat liver with subchronic exposure to genotoxic carcinogens riddelliine and aristolochic acid. Mol. Carcinog. 2015, 54, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Coulter, J.B.; O’Driscoll, C.M.; Bressler, J.P. Hydroquinone increases 5-hydroxymethylcytosine formation through ten eleven translocation 1 (TET1) 5-methylcytosine dioxygenase. J. Biol. Chem. 2013, 288, 28792–28800. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, S.P.; Burda, B.U.; Senger, C.A.; Lin, J.S.; Whitlock, E.P. Vitamin and mineral supplements in the primary prevention of cardiovascular disease and cancer: An updated systematic evidence review for the U.S. Preventive services task force. Ann. Intern. Med. 2013, 159, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Verrax, J.; Beck, R.; Dejeans, N.; Glorieux, C. Redox-active quinones and ascorbate: An innovative cancer therapy that exploits the vulnerability of cancer cells to oxidative stress. Anti-Cancer Agents Med. Chem. 2011, 11, 213–221. [Google Scholar] [CrossRef]
- Blaschke, K.; Ebata, K.T.; Karimi, M.M.; Zepeda-Martínez, J.A.; Goyal, P.; Mahapatra, S.; Tam, A.; Laird, D.J.; Hirst, M.; Rao, A.; et al. Vitamin C induces TET-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 2013, 500, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Minor, E.A.; Court, B.L.; Young, J.I.; Wang, G. Ascorbate induces ten-eleven translocation (TET) methylcytosine dioxygenase-mediated generation of 5-hydroxymethylcytosine. J. Biol. Chem. 2013, 288, 13669–13674. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Yang, Y.; Wang, X.; Chong, Z.; Yin, R.; Song, S.-H.; Zhao, C.; Li, C.; Huang, H.; Sun, B.-F.; et al. Redox-active quinones induces genome-wide DNA methylation changes by an iron-mediated and TET-dependent mechanism. Nucleic Acids Res. 2014, 42, 1593–1605. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Beck, F.W.J.; Snell, D.C.; Kucuk, O. Zinc in cancer prevention. Nutr. Cancer 2009, 61, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Zhao, Y.-P.; Jiang, Y.-G.; Wang, R.-W.; Ma, Z. Restoring the metabolic disturbance of zinc: May not only contribute to the prevention of esophageal squamous cell cancer. Med. Hypotheses 2008, 71, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.M.; Kristal, B.S.; Effron, M.S.; Shestopalov, A.I.; Ullucci, P.A.; Sheu, K.F.; Blass, J.P.; Cooper, A.J.L. Zn2+ inhibits α-ketoglutarate-stimulated mitochondrial respiration and the isolated α-ketoglutarate dehydrogenase complex. J. Biol. Chem. 2000, 275, 13441–13447. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.J.; Kim, M.R.; Chen, Y.S.; Yang, J.Y.; Oncogene, C.-C.J. Retinoic acid directs breast cancer cell state changes through regulation of TET2-pkcζ pathway. Oncogene 2017, 36, 3193–3206. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Kolendowski, B.; Isovic, M.; Reports, B.-K. Regulation of active DNA demethylation through RAR-mediated recruitment of a TET/TDG complex. Cell Rep. 2017, 19, 1685–1697. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Guyot, R.; Samarut, J.; Flamant, F.; Wong, J.; Gauthier, K. Methylcytosine dioxygenase TET3 interacts with thyroid hormone nuclear receptors and stabilizes their association to chromatin. Proc. Natl. Acad. Sci. USA 2017, 114, 8229–8234. [Google Scholar] [CrossRef] [PubMed]
- Arab, K.; Park, Y.; Lindroth, A.M.; Schäfer, A.; Oakes, C.; Weichenhan, D.; Lukanova, A.; Lundin, E.; Risch, A.; Meister, M.; et al. Long noncoding RNA tarid directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol. Cell 2014, 55, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Neri, F.; Incarnato, D.; Krepelova, A.; Rapelli, S.; Pagnani, A.; Zecchina, R.; Parlato, C.; Oliviero, S. Genome-wide analysis identifies a functional association of TET1 and polycomb repressive complex 2 in mouse embryonic stem cells. Genome Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Putiri, E.L.; Tiedemann, R.L.; Thompson, J.J.; Liu, C.; Ho, T.; Choi, J.-H.; Robertson, K.D. Distinct and overlapping control of 5-methylcytosine and 5-hydroxymethylcytosine by the TET proteins in human cancer cells. Genome Biol. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Szulwach, K.E.; Li, X.; Li, Y.; Song, C.X.; Han, J.W.; Kim, S.; Namburi, S.; Hermetz, K.; Kim, J.J.; Rudd, M.K.; et al. Integrating 5-hydroxymethylcytosine into the epigenomic landscape of human embryonic stem cells. PLoS Genet. 2011, 7, e1002154. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, K.; Shinoda, A.; Kano, F.; Sato, R.; Shirahige, K.; Murata, M. Pparγ-induced parylation promotes local DNA demethylation by production of 5-hydroxymethylcytosine. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Kafer, G.; Li, X.; Horii, T.; Suetake, I.; Tajima, S.; Hatada, I.; Carlton, P. 5-hydroxymethylcytosine marks sites of DNA damage and promotes genome stability. Cell Rep. 2016, 14, 1283–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
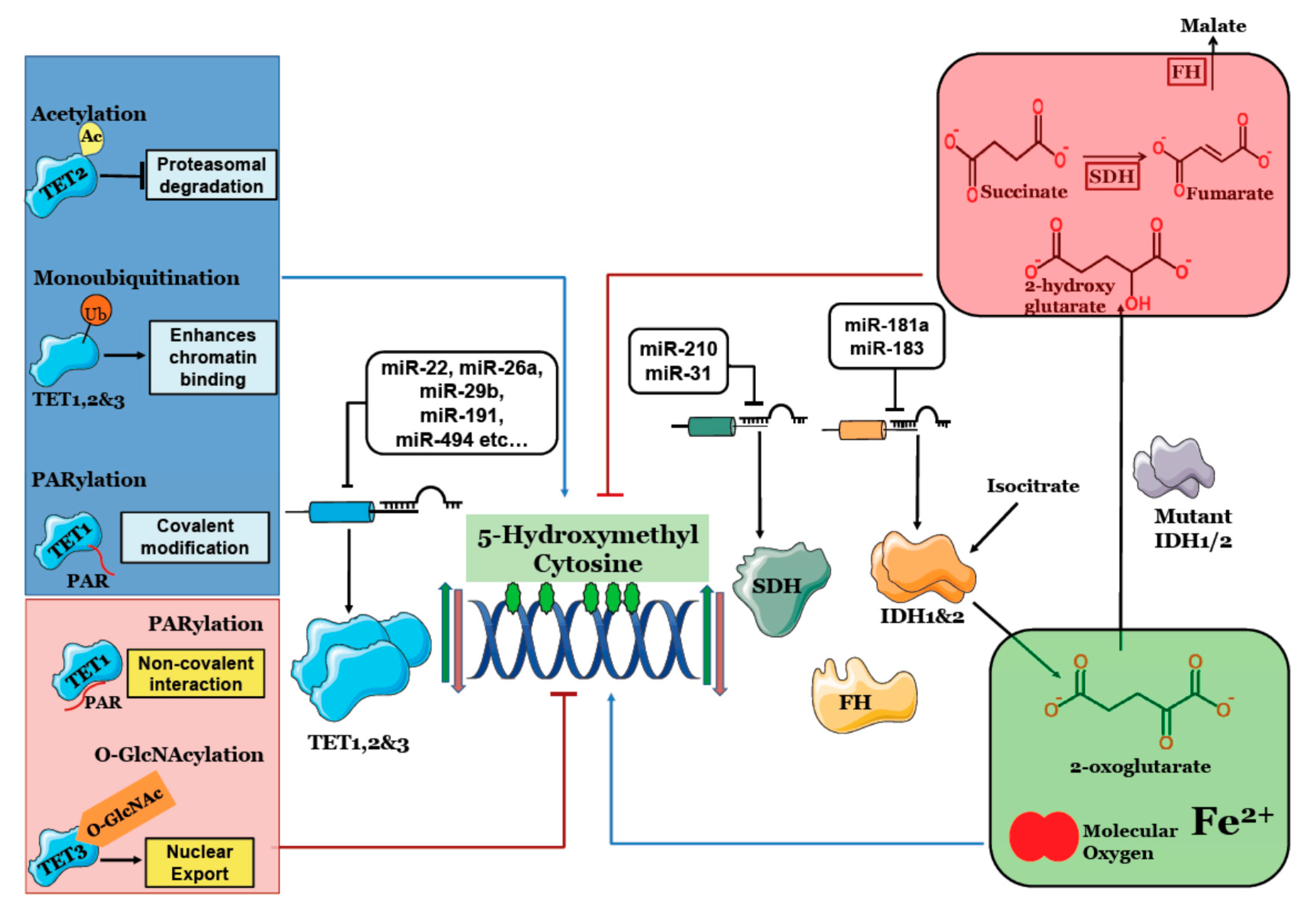
Figure 1.
Regulation of 5 Hydroxymethyl Cytosine: TET1, 2 & 3 (Ten Eleven Translocation) catalyze conversion of 5mC to 5hmC. IDH1 & 2 provides 2-oxoglutarate for 5hmC generation, Succinate Dehydrogenase (SDH) and Fumarate Hydratase (FH) clear succinate and fumarate that competitively inhibit TET enzymes. MicroRNAs targeting TET, IDH and SDH deregulate 5hmC content. Post-translational modifications like acetylation, monoubiquitination and covalent PARylation increase TET activity and 5hmC levels while O-GlcNAcylation and non-covalent PARylation reduce 5hmC. Mutant IDH1/2 produces 2-hydroxyglutarate that inhibits TET activity and significantly lowers 5hmC levels.
Figure 1.
Regulation of 5 Hydroxymethyl Cytosine: TET1, 2 & 3 (Ten Eleven Translocation) catalyze conversion of 5mC to 5hmC. IDH1 & 2 provides 2-oxoglutarate for 5hmC generation, Succinate Dehydrogenase (SDH) and Fumarate Hydratase (FH) clear succinate and fumarate that competitively inhibit TET enzymes. MicroRNAs targeting TET, IDH and SDH deregulate 5hmC content. Post-translational modifications like acetylation, monoubiquitination and covalent PARylation increase TET activity and 5hmC levels while O-GlcNAcylation and non-covalent PARylation reduce 5hmC. Mutant IDH1/2 produces 2-hydroxyglutarate that inhibits TET activity and significantly lowers 5hmC levels.
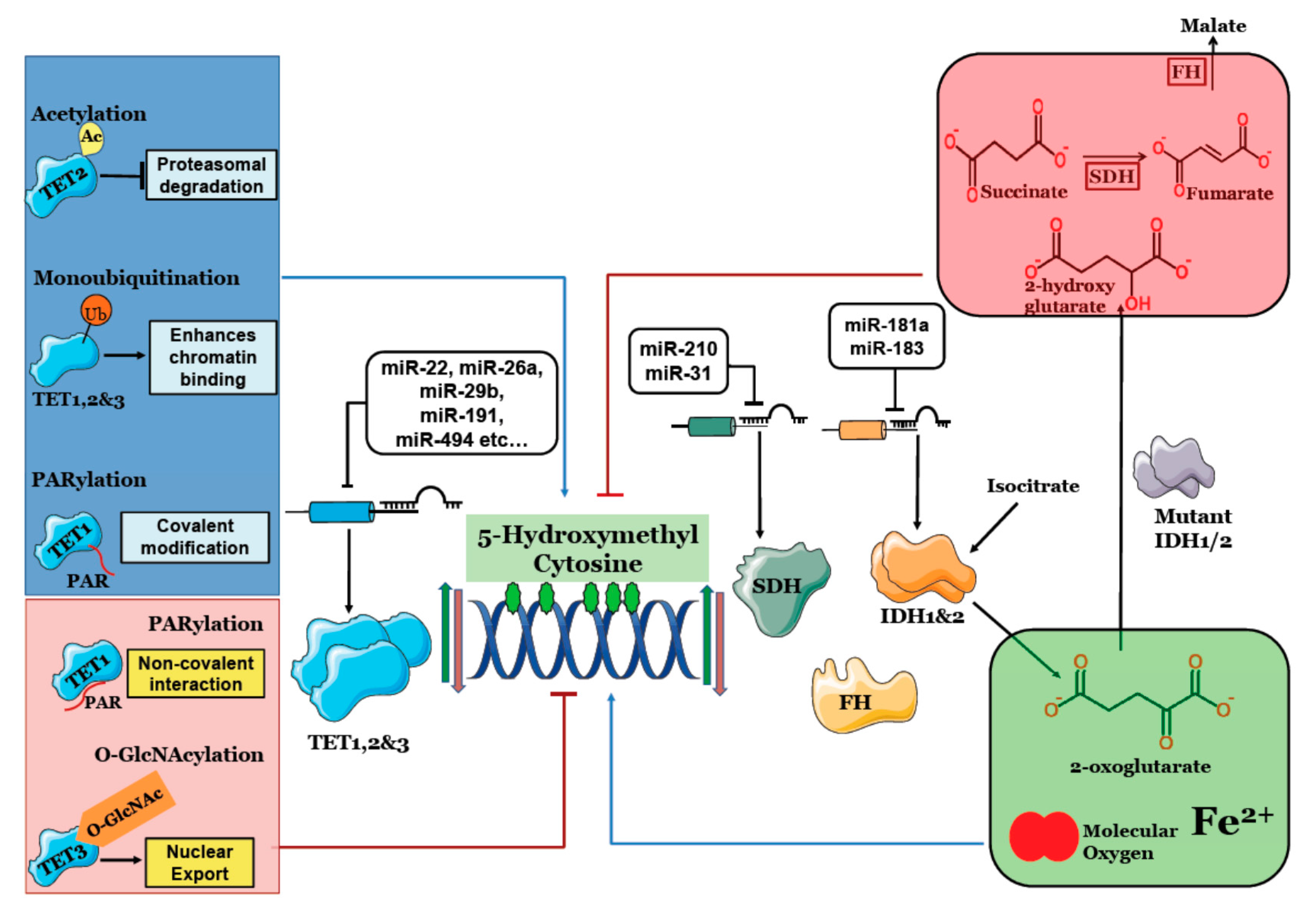
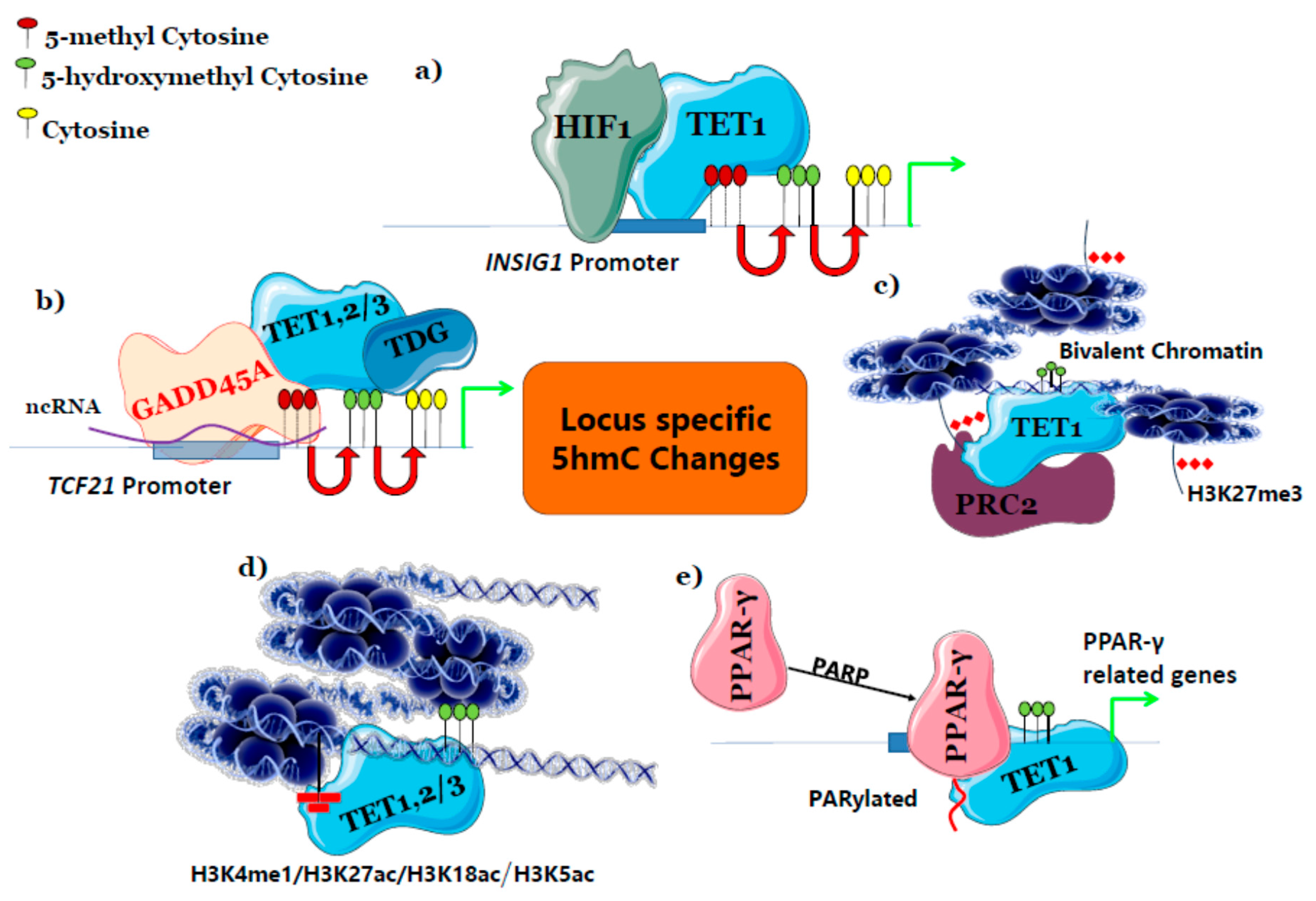
Figure 2.
Possible mechanisms governing locus-specific 5hmC changes. (a) Transcription Factors directly recruit TET and enriches 5hmC at their target genes, (b) TET binding protein indirectly specifies locus by connecting other DNA binding proteins and TET to enrich 5hmC, (c) PRC2 complex that represses gene expression enriches 5hmC by SUZ12 protein of PRC2 complex and makes it as bivalent chromatin, (d) histone modification co-localizes and enriches 5hmC at different regulatory regions, (e) post-translational modification of TF enables TET binding thereby recruiting to its target gene regions. INSIG1: Insulin Induced Gene 1; TDG: Thymine-DNA Glycosylase; TCF21: Transcription Factor 21; PARP: Poly ADP-ribose polymerase.
Figure 2.
Possible mechanisms governing locus-specific 5hmC changes. (a) Transcription Factors directly recruit TET and enriches 5hmC at their target genes, (b) TET binding protein indirectly specifies locus by connecting other DNA binding proteins and TET to enrich 5hmC, (c) PRC2 complex that represses gene expression enriches 5hmC by SUZ12 protein of PRC2 complex and makes it as bivalent chromatin, (d) histone modification co-localizes and enriches 5hmC at different regulatory regions, (e) post-translational modification of TF enables TET binding thereby recruiting to its target gene regions. INSIG1: Insulin Induced Gene 1; TDG: Thymine-DNA Glycosylase; TCF21: Transcription Factor 21; PARP: Poly ADP-ribose polymerase.

{kind=link}
{kind=link}
Table 1.
Isocitrate dehydrogenase(IDH), succinate dehydrogenase (SDH) and fumarate hydratase (FH) mutations in different cancers.
Table 1.
Isocitrate dehydrogenase(IDH), succinate dehydrogenase (SDH) and fumarate hydratase (FH) mutations in different cancers.
| Mutation | Cancer Types | 5hmC Levels | References |
|---|---|---|---|
| IDH1 | Grade II gliomas and secondary Glioblastoma, AML, Acute B lymphoblastic Leukemia, Paraganglioma, melanoma, Intrahepatic cholangiocarcinoma, cartilageous, thyroid, colon, Prostate and follicular cancer. | Lowered | [32,35,36] |
| IDH2 | Gliomas, AML, Paraganglioma, melanoma, cartilageous, Intrahepatic cholangiocarcinoma and colon cancer. | Lowered | [32,36] |
| SDHB/C/D | HPGL/PCC, pancreatic neuroendocrine tumours, renal tumorigenesis and gastric gastrointestinal stromal tumors. | Lowered | [19,37,38] |
| FH | leiomyomas of the skin and the uterus (fibroids, myomas), renal cell carcinoma. | Lowered | [20,39] |
Table 2.
Imbalance in 5-hydroxymethylcytosine (5hmC) levels in different cancer.
| Cancer Type | Locus/Region | Role or Effect | References |
|---|---|---|---|
| Breast cancer | Loss of 5hmC in HOXA | High Mobility Group AT-Hook 2 (HMGA2) promotes breast cancer cell invasion in part through inhibition of TET1-mediated demethylation and HOXA gene expression. | [40] |
| Breast cancer | Loss of 5hmC in miR200 promoter region | miR-22 overexpression targets TET1, 2 & 3 mRNA and inhibit demethylation of mir-200 and enable EMT, invasiveness and metastasis. | [42] |
| Breast cancer | Loss of 5hmC in LZTS1 gene | Decreases of 5hmC levels in LZTS1 region down-regulates its expression that enhance cancer progression and metastasis. | [43] |
| Breast cancer | Gain of 5hmC in TNFA gene | Increased TNFA expression is essential for BTIC properties by TNFα-p38-MAPK signalling. | [41] |
| Colon cancer | Loss of 5hmC or hypermethylation in DDK3 genes | Down-regulation of TET1 and DNA hydroxymethylation mediated by TET1 controlling the WNT signaling is a key player in tumor growth. | [46] |
| Colon cancer | The gain of 5hmC in Nuclear factor-erythroid 2-related factor 2 (Nrf2) favors resistance towards 5FU | 5-FU resistance in CRC involves the upregulation of Nrf2 and HO-1 expression via epigenetic modifications of DNA demethylation. | [52] |
| Colon cancer | Loss of 5hmC in CA2, FMN2, PDCD4, PKIB and SLC26A2. Gain of 5hmC in BMP7, NKD2, TESC and TGFBI | Though loss of 5hmC a hallmark of cancer, locus-specific changes also play a role in colorectal cancer. | [45] |
| Lung cancer | Loss of 5hmC in SOD3 gene | SOD3 which has been downregulated in A549, THP1 and other cancer cell lines has shown to be upregulated by overexpression of TET1CD which demethylates SOD3 promoter. | [44] |
Table 3.
Post-translational modification of Ten Eleven Translocation (TET) and its effect on 5hmC levels.
Table 3.
Post-translational modification of Ten Eleven Translocation (TET) and its effect on 5hmC levels.
| Post-Translational Modification | Regulatory Protein | Function | Effect on 5hmC | Cell Type/Model Cell Line | Reference |
|---|---|---|---|---|---|
| Acetylation of TET2 at K110 | p300 acetylates TET2 under oxidative stress, while HDAC1/2 deacetylates TET2 | Increases TET2 half-life and activity especially at hypermethylated sites under oxidative stress. | Increased | Ovarian and Colorectal cancer cell line (A2780 and HCT116) | [66] |
| Monoubiquitination | CRL4VprBP ubiquitylates TET1 (K1212),TET2 (K1299) and TET3 (K983) | Promotes TET DNA binding | Increased | MEF | [67] |
| PARylation (Covalent) | PARP-1 covalently links PAR to TET1 | Increased TET1 activity | Increased | HEK293 | [68] |
| PARylation (Non-Covalent) | Non-covalent binding of poly ADP-ribose to TET1 catalytic domain. | Decreases DNA binding | Decreased | HEK293 | [68] |
| O-GlcNAcylation | OGT O-GlcNAcylates TET3 | Enables nuclear export | Decreased | Not cell type specific | [69] |
HDAC—Histone Deacetylase, PARP—Poly ADP-ribose polymerase, PAR—Poly ADP-ribose.
Table 4.
TET, IDH and SDH targeting miRNAs and their functional effect.
| microRNA | Target | Role | Level of 5hmC | Reference |
|---|---|---|---|---|
| miR-181a | IDH1 | Increased miR-181a lowers IDH1 thereby lowers lipid synthesis and increases lipid oxidation enzymes | ND * | [77] |
| miR-183 | IDH2 | Lowers IDH2 and increases HIF1A and VEGF. upregulated in malignant gliomas | ND * | [79] |
| miR-210 | SDHD | Overexpression of miR-210 induced cell death and activates HIF1α in late stages of lung cancer | ND * | [78] |
| miR-31 | SDHA | Modulates mitochondrial metabolism that enables iPSC reprogramming. | ND * | [80] |
| miR-22 | TET2 | Increased in MDS and leukemia and increased expression linked to poor survival. | reduced | [81] |
| miR-125b, miR-29b, miR-29c, miR-101, miR-7, etc (more than 30) | TET2 | Increased in myeloid malignancies. | reduced | [73] |
| miR-26a | TET1, 2 & 3 | Promotes pancreatic cell differentiation. | reduced | [72] |
| miR-29b | TET1 | Increases sharply during embryonic body formation and favors mesendoderm lineage differentiation. | reduced | [71] |
| miR-22 | TET2 | TET2 reduction favors EMT and metastasis by 5hmC reduction in miR 200b promoter region in breast cancer | reduced | [42] |
| miR-29a | TET1 | Increased miR-29a reduces TET1 and enhances metastasis in HCC. | reduced | [82] |
| miR-191 | TET1 | Increased in Intrahepatic cholangiocarcinoma; enables proliferation, migration and invasion | reduced | [76] |
| miR-30a | TET1 | Increased in pulmonary fibrosis, as a potential therapeutic target. | reduced | [83] |
| miR-520b | TET1 | Decreased in HCC and enables proliferation | ND * | [74] |
| miR-494 | TET1 | Increased in HCC and enables invasion | reduced | [75] |
* ND-Not determined, EMT—Epithelial Mesenchymal Transition, iPSC—induced pluripotent stem cells.
Table 5.
Locus-specific 5hmC changes in different conditions.
| Conditions | 5hmC Locus | Functions | References |
|---|---|---|---|
| Hypoxia | Promotor, TSS and genic regions of hypoxia-inducible genes like Vascular Endothelial Growth Factor A (VEGFA), BCL2 Interacting Protein 3 (BINP3), Enolase 1 (ENO1). | Enables EMT, angiogenesis and metastasis | [84] |
| Phenobarbital | Proximal Promotor regions of Cyp and Gst genes at initial exposure. During prolonged exposure Wisp, Cxcr7 and loss in Ndrf2 gene | A short-term exposure facilitates gene expression of xenobiotics metabolism and prolonged exposure induces HCC. | [101] |
| Drug-induced ROS | Promotors of Nrf2 genes | Cell survival and drug resistance | [52] |
| Elevated plasma glucose | Promotor regions of Hexokinase in liver | Glucose metabolism | [62] |
| DNA damage by aphidicolin and micro-irradiation | Marks sites of DNA damage | Facilitates repair mechanism and genomic integrity | [121] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arvinden, V.R.; Deva Magendhra Rao, A.K.; Rajkumar, T.; Mani, S. Regulation and Functional Significance of 5-Hydroxymethylcytosine in Cancer. Epigenomes 2017, 1, 19. https://doi.org/10.3390/epigenomes1030019
AMA Style
Arvinden VR, Deva Magendhra Rao AK, Rajkumar T, Mani S. Regulation and Functional Significance of 5-Hydroxymethylcytosine in Cancer. Epigenomes. 2017; 1(3):19. https://doi.org/10.3390/epigenomes1030019
Chicago/Turabian StyleArvinden, Vittal Rangan, Arunagiri Kuha Deva Magendhra Rao, Thangarajan Rajkumar, and Samson Mani. 2017. "Regulation and Functional Significance of 5-Hydroxymethylcytosine in Cancer" Epigenomes 1, no. 3: 19. https://doi.org/10.3390/epigenomes1030019