
Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies


Abstract
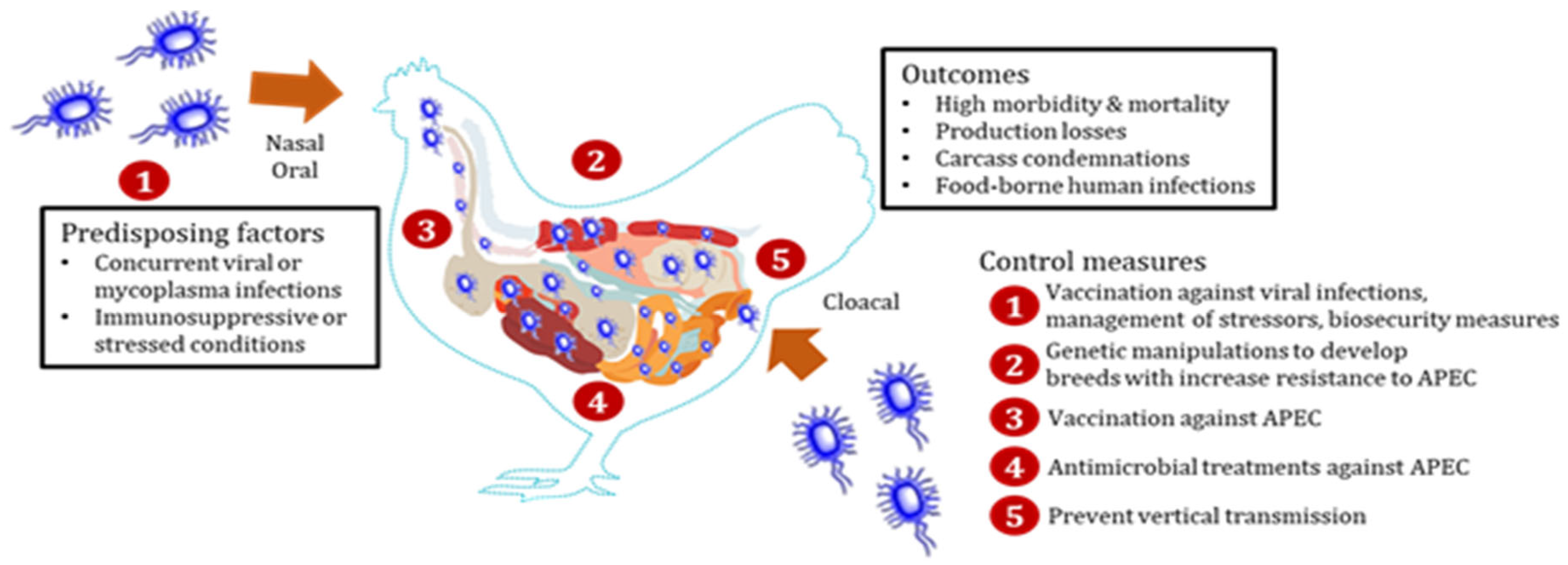
:1. Introduction
2. Virulence and Pathogenesis Factors
2.1. Adhesins
2.2. Invasins
2.3. Iron Acquisition Systems
2.4. Protectins
2.5. Toxins
2.6. Other Virulence and Pathogenesis Factors
2.6.1. Quorum-Sensing (QS) System
2.6.2. Secretion Systems
2.6.3. Two-Component Systems
2.6.4. Transcriptional Regulators
2.6.5. Metabolism-Associated Genes
2.6.6. Miscellaneous
2.7. Genes Essential for Systemic Infections and Adaptation in Chickens
3. Zoonotic Potential
3.1. Genetic Similarity and Commonality in Virulence Genes
3.2. Ability to Cause Disease with Similar Clinical Manifestations
4. Control Strategies
4.1. Management and Biosecurity Measures
4.2. Antibiotics
4.3. Vaccines
4.4. Probiotics
4.5. Bacteriophages
4.6. New Alternative Approaches
4.6.1. Innate Immune Stimulants
4.6.2. APEC Virulence and Growth Inhibitors
4.6.3. Antimicrobial Peptides
5. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dho-Moulin, M.; Fairbrother, J.M. Avian pathogenic Escherichia coli (APEC). Vet. Res. 1999, 30, 299–316. [Google Scholar]
- Dziva, F.; Stevens, M.P. Colibacillosis in poultry: Unravelling the molecular basis of virulence of avian pathogenic Escherichia coli in their natural hosts. Avian Pathol. 2008, 37, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Guabiraba, R.; Schouler, C. Avian colibacillosis: Still many black holes. FEMS Microbiol. Lett. 2015, 362, fnv118. [Google Scholar] [CrossRef]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef] [Green Version]
- Ghunaim, H.; Abu-Madi, M.A.; Kariyawasam, S. Advances in vaccination against avian pathogenic Escherichia coli respiratory disease: Potentials and limitations. Vet. Microbiol. 2014, 172, 13–22. [Google Scholar] [CrossRef]
- de Brito, B.G.; Gaziri, L.C.J.; Vidotto, M.C. Virulence Factors and Clonal Relationships among Escherichia coli Strains Isolated from Broiler Chickens with Cellulitis. Infect. Immun. 2003, 71, 4175. [Google Scholar] [CrossRef] [Green Version]
- Lutful Kabir, S.M. Avian colibacillosis and salmonellosis: A closer look at epidemiology, pathogenesis, diagnosis, control and public health concerns. Int. J. Environ. Res. Public Health 2010, 7, 89–114. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of Minimal Predictors of Avian Pathogenic Escherichia coli Virulence for Use as a Rapid Diagnostic Tool. J. Clin. Microbiol. 2008, 46, 3987–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collingwood, C.; Kemmett, K.; Williams, N.; Wigley, P. Is the Concept of Avian Pathogenic Escherichia coli as a Single Pathotype Fundamentally Flawed? Front. Vet. Sci. 2014, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Palaniyandi, S.; Mitra, A.; Herren, C.D.; Zhu, X.; Mukhopadhyay, S. LuxS contributes to virulence in avian pathogenic Escherichia coli O78:K80:H9. Vet. Microbiol. 2013, 166, 567–575. [Google Scholar] [CrossRef]
- Ma, J.; Bao, Y.; Sun, M.; Dong, W.; Pan, Z.; Zhang, W.; Lu, C.; Yao, H. Two functional type VI secretion systems in avian pathogenic Escherichia coli are involved in different pathogenic pathways. Infect. Immun. 2014, 82, 3867–3879. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xu, X.; Liu, X.; Wang, D.; Liang, H.; Wu, X.; Tian, M.; Ding, C.; Wang, G.; Yu, S. Escherichia coli type III secretion system 2 regulator EtrA promotes virulence of avian pathogenic Escherichia coli. Microbiology 2017, 163, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yin, L.; Xue, M.; Wang, Z.; Song, X.; Shao, Y.; Liu, H.; Tu, J.; Qi, K. The transcriptional regulator PhoP mediates the tolC molecular mechanism on APEC biofilm formation and pathogenicity. Avian Pathol. 2020, 49, 211–220. [Google Scholar] [CrossRef]
- Zhuge, X.; Tang, F.; Zhu, H.; Mao, X.; Wang, S.; Wu, Z.; Lu, C.; Dai, J.; Fan, H. AutA and AutR, Two Novel Global Transcriptional Regulators, Facilitate Avian Pathogenic Escherichia coli Infection. Sci. Rep. 2016, 6, 25085. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, N.L.; Vande Vorde, J.A.; Baker, A.R.; Horn, F.; Li, G.; Logue, C.M.; Nolan, L.K. FNR Regulates the Expression of Important Virulence Factors Contributing to the Pathogenicity of Avian Pathogenic Escherichia coli. Front. Cell Infect. Microbiol. 2017, 7, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, P.R.; Herrero-Fresno, A.; Pors, S.E.; Ahmed, S.; Wang, D.; Thofner, I.; Antenucci, F.; Olsen, J.E. The membrane transporter PotE is required for virulence in avian pathogenic Escherichia coli (APEC). Vet. Microbiol. 2018, 216, 38–44. [Google Scholar] [CrossRef]
- De Paiva, J.B.; Leite, J.L.; da Silva, L.P.; Rojas, T.C.; de Pace, F.; Conceicao, R.A.; Sperandio, V.; da Silveira, W.D. Influence of the major nitrite transporter NirC on the virulence of a Swollen Head Syndrome avian pathogenic E. coli (APEC) strain. Vet. Microbiol. 2015, 175, 123–131. [Google Scholar] [CrossRef]
- Jiang, F.; An, C.; Bao, Y.; Zhao, X.; Jernigan, R.L.; Lithio, A.; Nettleton, D.; Li, L.; Wurtele, E.S.; Nolan, L.K.; et al. ArcA Controls Metabolism, Chemotaxis, and Motility Contributing to the Pathogenicity of Avian Pathogenic Escherichia coli. Infect. Immun. 2015, 83, 3545–3554. [Google Scholar] [CrossRef] [Green Version]
- Markland, S.M.; LeStrange, K.J.; Sharma, M.; Kniel, K.E. Old Friends in New Places: Exploring the Role of Extraintestinal E. coli in Intestinal Disease and Foodborne Illness. Zoonoses Public Health 2015, 62, 491–496. [Google Scholar] [CrossRef]
- Bélanger, L.; Garenaux, A.; Harel, J.; Boulianne, M.; Nadeau, E.; Dozois, C.M. Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli. FEMS Immunol. Med Microbiol. 2011, 62, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.M.; Stegger, M.; Aziz, M.; Johnson, T.J.; Waits, K.; Nordstrom, L.; Gauld, L.; Weaver, B.; Rolland, D.; Statham, S.; et al. Escherichia coli ST131-H22 as a Foodborne Uropathogen. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Tivendale, K.A.; Logue, C.M.; Kariyawasam, S.; Jordan, D.; Hussein, A.; Li, G.; Wannemuehler, Y.; Nolan, L.K. Avian-pathogenic Escherichia coli strains are similar to neonatal meningitis E. coli strains and are able to cause meningitis in the rat model of human disease. Infect. Immun. 2010, 78, 3412–3419. [Google Scholar] [CrossRef] [Green Version]
- Agunos, A.; Léger, D.; Carson, C. Review of antimicrobial therapy of selected bacterial diseases in broiler chickens in Canada. Can. Vet. J. 2012, 53, 1289–1300. [Google Scholar]
- Nhung, N.T.; Chansiripornchai, N.; Carrique-Mas, J.J. Antimicrobial Resistance in Bacterial Poultry Pathogens: A Review. Front. Vet. Sci. 2017, 4, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregersen, R.H.; Christensen, H.; Ewers, C.; Bisgaard, M. Impact of Escherichia coli vaccine on parent stock mortality, first week mortality of broilers and population diversity of E. coli in vaccinated flocks. Avian Pathol. 2010, 39, 287–295. [Google Scholar] [CrossRef]
- Wang, S.; Peng, Q.; Jia, H.M.; Zeng, X.F.; Zhu, J.L.; Hou, C.L.; Liu, X.T.; Yang, F.J.; Qiao, S.Y. Prevention of Escherichia coli infection in broiler chickens with Lactobacillus plantarum B1. Poult. Sci. 2017, 96, 2576–2586. [Google Scholar] [CrossRef] [PubMed]
- Kaikabo, A.A.; AbdulKarim, S.M.; Abas, F. Evaluation of the efficacy of chitosan nanoparticles loaded PhiKAZ14 bacteriophage in the biological control of colibacillosis in chickens. Poult. Sci. 2017, 96, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Allan, B.; Wheler, C.; Koster, W.; Sarfraz, M.; Potter, A.; Gerdts, V.; Dar, A. In Ovo Administration of Innate Immune Stimulants and Protection from Early Chick Mortalities due to Yolk Sac Infection. Avian Dis. 2018, 62, 316–321. [Google Scholar] [CrossRef]
- Peng, L.Y.; Yuan, M.; Cui, Z.Q.; Wu, Z.M.; Yu, Z.J.; Song, K.; Tang, B.; Fu, B.D. Rutin inhibits quorum sensing, biofilm formation and virulence genes in avian pathogenic Escherichia coli. Microb. Pathog. 2018, 119, 54–59. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial Peptides as Potential Alternatives to Antibiotics in Food Animal Industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Awad, A.M.; El-Shall, N.A.; Khalil, D.S.; Abd El-Hack, M.E.; Swelum, A.A.; Mahmoud, A.H.; Ebaid, H.; Komany, A.; Sammour, R.H.; Sedeik, M.E. Incidence, Pathotyping, and Antibiotic Susceptibility of Avian Pathogenic Escherichia coli among Diseased Broiler Chicks. Pathogens 2020, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, L.; Ge, Z.; Yuehua, L.; Yubin, G.; Rachid, K.; Mustapha, O.; Junwei, W.; Karine, O. Virulence traits of avian pathogenic (APEC) and fecal (AFEC) E. coli isolated from broiler chickens in Algeria. Trop. Anim. Health Prod. 2018, 50, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Mohsin, M.; Sajjad Ur, R.; Saleemi, M.K. Virulence-associated genes and antimicrobial resistance among avian pathogenic Escherichia coli from colibacillosis affected broilers in Pakistan. Trop. Anim. Health Prod. 2019, 51, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Maciel, J.F.; Matter, L.B.; Trindade, M.M.; Camillo, G.; Lovato, M.; de Avila Botton, S.; Castagna de Vargas, A. Virulence factors and antimicrobial susceptibility profile of extraintestinal Escherichia coli isolated from an avian colisepticemia outbreak. Microb. Pathog. 2017, 103, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Silveira, F.; Maluta, R.P.; Tiba, M.R.; de Paiva, J.B.; Guastalli, E.A.; da Silveira, W.D. Comparison between avian pathogenic (APEC) and avian faecal (AFEC) Escherichia coli isolated from different regions in Brazil. Vet. J. 2016, 217, 65–67. [Google Scholar] [CrossRef] [PubMed]
- De Paiva, J.B.; da Silva, L.P.; Casas, M.R.; Conceicao, R.A.; Nakazato, G.; de Pace, F.; Sperandio, V.; da Silveira, W.D. In vivo influence of in vitro up-regulated genes in the virulence of an APEC strain associated with swollen head syndrome. Avian Pathol. 2016, 45, 94–105. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cao, C.; Huan, H.; Zhang, L.; Mu, X.; Gao, Q.; Dong, X.; Gao, S.; Liu, X. Isolation, identification, and pathogenicity of O142 avian pathogenic Escherichia coli causing black proventriculus and septicemia in broiler breeders. Infect. Genet. Evol. 2015, 32, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Toufeer, M.; Narvaez Bravo, C.; Lai, V.; Rempel, H.; Manges, A.; Diarra, M.S. Characterization of Extraintestinal Pathogenic Escherichia coli isolated from retail poultry meats from Alberta, Canada. Int. J. Food Microbiol. 2014, 177, 49–56. [Google Scholar] [CrossRef]
- Delannoy, S.; Schouler, C.; Souillard, R.; Yousfi, L.; Le Devendec, L.; Lucas, C.; Bougeard, S.; Keita, A.; Fach, P.; Galliot, P.; et al. Diversity of Escherichia coli strains isolated from day-old broiler chicks, their environment and colibacillosis lesions in 80 flocks in France. Vet. Microbiol. 2021, 252, 108923. [Google Scholar] [CrossRef]
- Zhuge, X.; Zhou, Z.; Jiang, M.; Wang, Z.; Sun, Y.; Tang, F.; Xue, F.; Ren, J.; Dai, J. Chicken-source Escherichia coli within phylogroup F shares virulence genotypes and is closely related to extraintestinal pathogenic E. coli causing human infections. Transbound Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Ren, J.; Chen, L.; Zhuge, X.; Hu, L.; Li, D.; Tang, F.; Dai, J. The YfcO fimbriae gene enhances adherence and colonization abilities of avian pathogenic Escherichia coli in vivo and in vitro. Microb. Pathog. 2016, 100, 56–61. [Google Scholar] [CrossRef]
- Verma, R.; Rojas, T.C.; Maluta, R.P.; Leite, J.L.; da Silva, L.P.; Nakazato, G.; Dias da Silveira, W. Fimbria-Encoding Gene yadC Has a Pleiotropic Effect on Several Biological Characteristics and Plays a Role in Avian Pathogenic Escherichia coli Pathogenicity. Infect. Immun. 2016, 84, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuge, X.; Wang, S.; Fan, H.; Pan, Z.; Ren, J.; Yi, L.; Meng, Q.; Yang, X.; Lu, C.; Dai, J. Characterization and functional analysis of AatB, a novel autotransporter adhesin and virulence factor of avian pathogenic Escherichia coli. Infect. Immun. 2013, 81, 2437–2447. [Google Scholar] [CrossRef] [Green Version]
- Zhu-Ge, X.K.; Pan, Z.H.; Tang, F.; Mao, X.; Hu, L.; Wang, S.H.; Xu, B.; Lu, C.P.; Fan, H.J.; Dai, J.J. The effects of upaB deletion and the double/triple deletion of upaB, aatA, and aatB genes on pathogenicity of avian pathogenic Escherichia coli. Appl. Microbiol. Biotechnol. 2015, 99, 10639–10654. [Google Scholar] [CrossRef]
- Ali, A.; Kolenda, R.; Khan, M.M.; Weinreich, J.; Li, G.; Wieler, L.H.; Tedin, K.; Roggenbuck, D.; Schierack, P. Novel Avian Pathogenic Escherichia coli Genes Responsible for Adhesion to Chicken and Human Cell Lines. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Wang, S.; Niu, C.; Shi, Z.; Xia, Y.; Yaqoob, M.; Dai, J.; Lu, C. Effects of ibeA deletion on virulence and biofilm formation of avian pathogenic Escherichia coli. Infect. Immun. 2011, 79, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Shi, Z.; Xia, Y.; Li, H.; Kou, Y.; Bao, Y.; Dai, J.; Lu, C. IbeB is involved in the invasion and pathogenicity of avian pathogenic Escherichia coli. Vet. Microbiol. 2012, 159, 411–419. [Google Scholar] [CrossRef]
- Wang, S.; Bao, Y.; Meng, Q.; Xia, Y.; Zhao, Y.; Wang, Y.; Tang, F.; ZhuGe, X.; Yu, S.; Han, X.; et al. IbeR facilitates stress-resistance, invasion and pathogenicity of avian pathogenic Escherichia coli. PLoS ONE 2015, 10, e0119698. [Google Scholar] [CrossRef]
- Pilatti, L.; Boldrin de Paiva, J.; Rojas, T.C.; Leite, J.L.; Conceicao, R.A.; Nakazato, G.; Dias da Silveira, W. The virulence factor ychO has a pleiotropic action in an Avian Pathogenic Escherichia coli (APEC) strain. BMC Microbiol. 2016, 16, 35. [Google Scholar] [CrossRef] [Green Version]
- Thomrongsuwannakij, T.; Blackall, P.J.; Djordjevic, S.P.; Cummins, M.L.; Chansiripornchai, N. A comparison of virulence genes, antimicrobial resistance profiles and genetic diversity of avian pathogenic Escherichia coli (APEC) isolates from broilers and broiler breeders in Thailand and Australia. Avian Pathol. 2020, 49, 457–466. [Google Scholar] [CrossRef]
- Tu, J.; Xue, T.; Qi, K.; Shao, Y.; Huang, B.; Wang, X.; Zhou, X. The irp2 and fyuA genes in High Pathogenicity Islands are involved in the pathogenesis of infections caused by avian pathogenic Escherichia coli (APEC). Pol. J. Vet. Sci. 2016, 19, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Sun, Q.; Zhao, L. Virulence Factors and Antibiotic Resistance of Avian Pathogenic Escherichia coli in Eastern China. J. Vet. Res. 2019, 63, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Varga, C.; Brash, M.L.; Slavic, D.; Boerlin, P.; Ouckama, R.; Weis, A.; Petrik, M.; Philippe, C.; Barham, M.; Guerin, M.T. Evaluating Virulence-Associated Genes and Antimicrobial Resistance of Avian Pathogenic Escherichia coli Isolates from Broiler and Broiler Breeder Chickens in Ontario, Canada. Avian Dis. 2018, 62, 291–299. [Google Scholar] [CrossRef]
- Paixao, A.C.; Ferreira, A.C.; Fontes, M.; Themudo, P.; Albuquerque, T.; Soares, M.C.; Fevereiro, M.; Martins, L.; Correa de Sa, M.I. Detection of virulence-associated genes in pathogenic and commensal avian Escherichia coli isolates. Poult. Sci. 2016, 95, 1646–1652. [Google Scholar] [CrossRef]
- Sabri, M.; Caza, M.; Proulx, J.; Lymberopoulos, M.H.; Bree, A.; Moulin-Schouleur, M.; Curtiss, R., 3rd; Dozois, C.M. Contribution of the SitABCD, MntH, and FeoB metal transporters to the virulence of avian pathogenic Escherichia coli O78 strain chi7122. Infect. Immun. 2008, 76, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, M.; Mohsin, M.; Johnson, T.J.; Smith, E.A.; Johnson, A.; Umair, M.; Saleemi, M.K.; Sajjad Ur, R. Genomic landscape of multi-drug resistant avian pathogenic Escherichia coli recovered from broilers. Vet. Microbiol. 2020, 247, 108766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jiang, S.; Liu, Y.; Sun, Y.; Yu, P.; Gong, Q.; Zeng, H.; Li, Y.; Xue, F.; Zhuge, X.; et al. Identification of ireA, 0007, 0008, and 2235 as TonB-dependent receptors in the avian pathogenic Escherichia coli strain DE205B. Vet. Res. 2020, 51, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Dai, J.; Zhuge, X.; Wang, H.; Hu, L.; Ren, J.; Chen, L.; Li, D.; Tang, F. Iron-regulated gene ireA in avian pathogenic Escherichia coli participates in adhesion and stress-resistance. BMC Vet. Res. 2016, 12, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, X.; Gao, R.; Xiao, W.; Gao, Q.; Cao, C.; Xu, H.; Gao, S.; Liu, X. EntE, EntS and TolC synergistically contributed to the pathogenesis of APEC strain E058. Microb. Pathog. 2020, 141, 103990. [Google Scholar] [CrossRef]
- Hejair, H.M.A.; Ma, J.; Zhu, Y.; Sun, M.; Dong, W.; Zhang, Y.; Pan, Z.; Zhang, W.; Yao, H. Role of outer membrane protein T in pathogenicity of avian pathogenic Escherichia coli. Res. Vet. Sci. 2017, 115, 109–116. [Google Scholar] [CrossRef]
- Song, X.; Qiu, M.; Jiang, H.; Xue, M.; Hu, J.; Liu, H.; Zhou, X.; Tu, J.; Qi, K. ybjX mutation regulated avian pathogenic Escherichia coli pathogenicity though stress-resistance pathway. Avian Pathol. 2020, 49, 144–152. [Google Scholar] [CrossRef]
- Song, X.; Hou, M.; Tu, J.; Xue, M.; Shao, Y.; Jiang, H.; Liu, H.; Xue, T.; Wang, G.; Qi, K. Outer membrane proteins YbjX and PagP co-regulate motility in Escherichia coli via the bacterial chemotaxis pathway. Res. Vet. Sci. 2019, 125, 279–284. [Google Scholar] [CrossRef]
- Nielsen, D.W.; Ricker, N.; Barbieri, N.L.; Allen, H.K.; Nolan, L.K.; Logue, C.M. Outer membrane protein A (OmpA) of extraintestinal pathogenic Escherichia coli. BMC Res. Notes 2020, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Tu, C.; Wang, Y.; Qi, K.; Hu, J.; Wang, Z.; Mi, R.; Yan, H.; Chen, Z.; Han, X. The role of the wzy gene in lipopolysaccharide biosynthesis and pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 2019, 127, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Han, X.; Wang, S.; Meng, Q.; Zhang, Y.; Ding, C.; Yu, S. The waaL gene is involved in lipopolysaccharide synthesis and plays a role on the bacterial pathogenesis of avian pathogenic Escherichia coli. Vet. Microbiol. 2014, 172, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ling, J.; Gao, Q.; He, H.; Mu, X.; Yan, Z.; Gao, S.; Liu, X. Role of the lpxM lipid A biosynthesis pathway gene in pathogenicity of avian pathogenic Escherichia coli strain E058 in a chicken infection model. Vet. Microbiol. 2013, 166, 516–526. [Google Scholar] [CrossRef]
- Gao, Q.; Xia, L.; Wang, X.; Ye, Z.; Liu, J.; Gao, S. SodA Contributes to the Virulence of Avian Pathogenic Escherichia coli O2 Strain E058 in Experimentally Infected Chickens. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Murase, K.; Martin, P.; Porcheron, G.; Houle, S.; Helloin, E.; Penary, M.; Nougayrede, J.P.; Dozois, C.M.; Hayashi, T.; Oswald, E. HlyF Produced by Extraintestinal Pathogenic Escherichia coli Is a Virulence Factor That Regulates Outer Membrane Vesicle Biogenesis. J. Infect. Dis. 2016, 213, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wang, C.L.; Chang, S.K.; Tsai, Y.L.; Chou, C.H. Characterization of Escherichia coli Isolated from Day-old Chicken Fluff in Taiwanese Hatcheries. Avian Dis. 2019, 63, 9–16. [Google Scholar] [CrossRef]
- Ibrahim, R.A.; Cryer, T.L.; Lafi, S.Q.; Basha, E.A.; Good, L.; Tarazi, Y.H. Identification of Escherichia coli from broiler chickens in Jordan, their antimicrobial resistance, gene characterization and the associated risk factors. BMC Vet. Res. 2019, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Sgariglia, E.; Aconiti Mandolini, N.; Napoleoni, M.; Medici, L.; Fraticelli, R.; Conquista, M.; Gianfelici, P.; Staffolani, M.; Fisichella, S.; Capuccella, M.; et al. Antibiotic resistance pattern and virulence genesin avian pathogenic Escherichia coli (APEC) from different breeding systems. Vet. Ital. 2019, 55, 26–33. [Google Scholar] [CrossRef]
- Dou, X.; Gong, J.; Han, X.; Xu, M.; Shen, H.; Zhang, D.; Zhuang, L.; Liu, J.; Zou, J. Characterization of avian pathogenic Escherichia coli isolated in eastern China. Gene 2016, 576, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Yin, H.; Hu, J.; Miao, J.; Chen, Z.; Qi, K.; Wang, Z.; Gong, J.; Phouthapane, V.; Jiang, W.; et al. Lsr operon is associated with AI-2 transfer and pathogenicity in avian pathogenic Escherichia coli. Vet. Res. 2019, 50, 109. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.Q.; Wu, Z.M.; Fu, Y.X.; Xu, D.X.; Guo, X.; Shen, H.Q.; Wei, X.B.; Yi, P.F.; Fu, B.D. Autoinducer-2 of quorum sensing is involved in cell damage caused by avian pathogenic Escherichia coli. Microb. Pathog. 2016, 99, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Lv, X.; Lu, J.; Yu, S.; Jin, Y.; Hu, J.; Zuo, J.; Mi, R.; Huang, Y.; Qi, K.; et al. The role of the ptsI gene on AI-2 internalization and pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 2017, 113, 321–329. [Google Scholar] [CrossRef]
- Xu, D.; Zuo, J.; Chen, Z.; Lv, X.; Hu, J.; Wu, X.; Qi, K.; Mi, R.; Huang, Y.; Miao, J.; et al. Different activated methyl cycle pathways affect the pathogenicity of avian pathogenic Escherichia coli. Vet. Microbiol. 2017, 211, 160–168. [Google Scholar] [CrossRef]
- Xue, M.; Xiao, Y.; Fu, D.; Raheem, M.A.; Shao, Y.; Song, X.; Tu, J.; Xue, T.; Qi, K. Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli. Animals 2020, 10, 1658. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X.; Xu, X.; Yang, D.; Wang, D.; Han, X.; Shi, Y.; Tian, M.; Ding, C.; Peng, D.; et al. Escherichia coli Type III Secretion System 2 ATPase EivC Is Involved in the Motility and Virulence of Avian Pathogenic Escherichia coli. Front. Microbiol. 2016, 7, 1387. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Wang, D.; Xin, S.; Zhou, D.; Li, T.; Tian, M.; Qi, J.; Ding, C.; Wang, S.; Yu, S. The CpxR regulates type VI secretion system 2 expression and facilitates the interbacterial competition activity and virulence of avian pathogenic Escherichia coli. Vet. Res. 2019, 50, 40. [Google Scholar] [CrossRef] [Green Version]
- Matter, L.B.; Ares, M.A.; Abundes-Gallegos, J.; Cedillo, M.L.; Yanez, J.A.; Martinez-Laguna, Y.; De la Cruz, M.A.; Giron, J.A. The CpxRA stress response system regulates virulence features of avian pathogenic Escherichia coli. Environ. Microbiol. 2018, 20, 3363–3377. [Google Scholar] [CrossRef]
- Wang, S.; Dai, J.; Meng, Q.; Han, X.; Han, Y.; Zhao, Y.; Yang, D.; Ding, C.; Yu, S. DotU expression is highly induced during in vivo infection and responsible for virulence and Hcp1 secretion in avian pathogenic Escherichia coli. Front. Microbiol. 2014, 5, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Pace, F.; Boldrin de Paiva, J.; Nakazato, G.; Lancellotti, M.; Sircili, M.P.; Guedes Stehling, E.; Dias da Silveira, W.; Sperandio, V. Characterization of IcmF of the type VI secretion system in an avian pathogenic Escherichia coli (APEC) strain. Microbiology 2011, 157, 2954–2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Zhang, Q.; Wang, H.; Quan, G.; Zhang, D.; Ren, W.; Liao, Y.; Xia, P.; Zhu, G. The different roles of hcp1 and hcp2 of the type VI secretion system in Escherichia coli strain CE129. J. Basic Microbiol. 2018, 58, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Hou, M.; Jiang, H.; Shen, X.; Xue, M.; Shao, Y.; Wang, L.; He, Q.; Zheng, L.; Tu, J.; et al. Hcp2a of type VI secretion system contributes to IL8 and IL1β expression of chicken tracheal epithelium by affecting APEC colonization. Res. Vet. Sci. 2020, 132, 279–284. [Google Scholar] [CrossRef]
- Tu, J.; Huang, B.; Zhang, Y.; Xue, T.; Li, S.; Qi, K. Modulation of virulence genes by the two-component system PhoP-PhoQ in avian pathogenic Escherichia coli. Pol. J. Vet. Sci. 2016, 19, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Raheem, M.A.; Gu, Y.; Lu, H.; Song, X.; Tu, J.; Xue, T.; Qi, K. The KdpD/KdpE two-component system contributes to the motility and virulence of avian pathogenic Escherichia coli. Res. Vet. Sci. 2020, 131, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Su, S.; Li, X.; Wang, H.; Liu, J.; Gao, S. Transcriptional analysis of RstA/RstB in avian pathogenic Escherichia coli identifies its role in the regulation of hdeD-mediated virulence and survival in chicken macrophages. Vet. Microbiol. 2020, 241, 108555. [Google Scholar] [CrossRef]
- Herren, C.D.; Mitra, A.; Palaniyandi, S.K.; Coleman, A.; Elankumaran, S.; Mukhopadhyay, S. The BarA-UvrY two-component system regulates virulence in avian pathogenic Escherichia coli O78:K80:H9. Infect. Immun. 2006, 74, 4900–4909. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Li, Q.; Xue, M.; Wang, Z.; Tu, J.; Song, X.; Shao, Y.; Han, X.; Xue, T.; Liu, H.; et al. The role of the phoP transcriptional regulator on biofilm formation of avian pathogenic Escherichia coli. Avian Pathol. 2019, 48, 362–370. [Google Scholar] [CrossRef]
- Yu, L.; Wang, H.; Han, X.; Li, W.; Xue, M.; Qi, K.; Chen, X.; Ni, J.; Deng, R.; Shang, F.; et al. The two-component system, BasSR, is involved in the regulation of biofilm and virulence in avian pathogenic Escherichia coli. Avian Pathol. 2020, 49, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Ye, Z.; Wang, X.; Mu, X.; Gao, S.; Liu, X. RstA is required for the virulence of an avian pathogenic Escherichia coli O2 strain E058. Infect. Genet. Evol. 2015, 29, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, H.; Gurlebeck, D.; Gross, J.; Dreck, K.; Pannen, D.; Ewers, C.; Wieler, L.H.; Schnetz, K. YjjQ Represses Transcription of flhDC and Additional Loci in Escherichia coli. J. Bacteriol. 2015, 197, 2713–2720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Li, W.; Qi, K.; Wang, S.; Chen, X.; Ni, J.; Deng, R.; Shang, F.; Xue, T. McbR is involved in biofilm formation and H2O2 stress response in avian pathogenic Escherichia coli X40. Poult. Sci. 2019, 98, 4094–4103. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, H.; Wang, X.; Zhang, D.; Ye, Z.; Gao, S.; Liu, X. RfaH promotes the ability of the avian pathogenic Escherichia coli O2 strain E058 to cause avian colibacillosis. J. Bacteriol. 2013, 195, 2474–2480. [Google Scholar] [CrossRef] [Green Version]
- Zhuge, X.; Sun, Y.; Jiang, M.; Wang, J.; Tang, F.; Xue, F.; Ren, J.; Zhu, W.; Dai, J. Acetate metabolic requirement of avian pathogenic Escherichia coli promotes its intracellular proliferation within macrophage. Vet. Res. 2019, 50, 31. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Chen, Y.; Qian, X.; Liu, Y.; Ren, J.; Xue, F.; Sun, J.; Tang, F.; Dai, J. orf20 in prophage phiv142-3 contributes to the adhesion and colonization ability of avian pathogenic Escherichia coli strain DE142 by affecting the formation of flagella and I fimbriae. Vet. Microbiol. 2019, 235, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tang, F.; Xue, F.; Ren, J.; Liu, Y.; Yang, D.; Dai, J. Prophage phiv142-3 enhances the colonization and resistance to environmental stresses of avian pathogenic Escherichia coli. Vet. Microbiol. 2018, 218, 70–77. [Google Scholar] [CrossRef]
- Liu, Y.; Gong, Q.; Qian, X.; Li, D.; Zeng, H.; Li, Y.; Xue, F.; Ren, J.; Zhu Ge, X.; Tang, F.; et al. Prophage phiv205-1 facilitates biofilm formation and pathogenicity of avian pathogenic Escherichia coli strain DE205B. Vet. Microbiol. 2020, 247, 108752. [Google Scholar] [CrossRef]
- Hejair, H.M.A.; Zhu, Y.; Ma, J.; Zhang, Y.; Pan, Z.; Zhang, W.; Yao, H. Functional role of ompF and ompC porins in pathogenesis of avian pathogenic Escherichia coli. Microb. Pathog. 2017, 107, 29–37. [Google Scholar] [CrossRef]
- Verma, R.; Rojas, T.C.G.; Maluta, R.P.; Leite, J.L.; Nakazato, G.; de Silveira, W.D. Role of hypothetical protein YicS in the pathogenicity of Avian Pathogenic Escherichia coli in vivo and in vitro. Microbiol. Res. 2018, 214, 28–36. [Google Scholar] [CrossRef]
- Liu, H.; Chen, L.; Si, W.; Wang, C.; Zhu, F.; Li, G.; Liu, S. Physiology and pathogenicity of cpdB deleted mutant of avian pathogenic Escherichia coli. Res. Vet. Sci. 2017, 111, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Lamarche, M.G.; Dozois, C.M.; Daigle, F.; Caza, M.; Curtiss, R., 3rd; Dubreuil, J.D.; Harel, J. Inactivation of the pst system reduces the virulence of an avian pathogenic Escherichia coli O78 strain. Infect. Immun. 2005, 73, 4138–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, X.; Huan, H.; Xu, H.; Gao, Q.; Xiong, L.; Gao, R.; Gao, S.; Liu, X. The transfer-messenger RNA-small protein B system plays a role in avian pathogenic Escherichia coli pathogenicity. J. Bacteriol. 2013, 195, 5064–5071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderkelen, L.; Ons, E.; Van Herreweghe, J.M.; Callewaert, L.; Goddeeris, B.M.; Michiels, C.W. Role of lysozyme inhibitors in the virulence of avian pathogenic Escherichia coli. PLoS ONE 2012, 7, e45954. [Google Scholar] [CrossRef] [Green Version]
- Mbanga, J.; Nyararai, Y.O. Virulence gene profiles of avian pathogenic Escherichia coli isolated from chickens with colibacillosis in Bulawayo, Zimbabwe. Onderstepoort J. Vet. Res. 2015, 82, e1–e8. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, W.; Tanaka, H.; Taniguchi, Y.; Iimura, M.; Soga, E.; Kubo, R.; Matsuo, N.; Kawamura, K.; Arakawa, Y.; Nagano, Y.; et al. Acquisition of mcr-1 and Cocarriage of Virulence Genes in Avian Pathogenic Escherichia coli Isolates from Municipal Wastewater Influents in Japan. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Saha, O.; Hoque, M.N.; Islam, O.K.; Rahaman, M.M.; Sultana, M.; Hossain, M.A. Multidrug-Resistant Avian Pathogenic Escherichia coli Strains and Association of Their Virulence Genes in Bangladesh. Microorganisms 2020, 8, 1135. [Google Scholar] [CrossRef] [PubMed]
- Tuntufye, H.N.; Lebeer, S.; Gwakisa, P.S.; Goddeeris, B.M. Identification of Avian pathogenic Escherichia coli genes that are induced in vivo during infection in chickens. Appl. Environ. Microbiol. 2012, 78, 3343–3351. [Google Scholar] [CrossRef] [Green Version]
- Dozois, C.M.; Daigle, F.; Curtiss, R., 3rd. Identification of pathogen-specific and conserved genes expressed in vivo by an avian pathogenic Escherichia coli strain. Proc. Natl. Acad. Sci. USA 2003, 100, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, X.; Nolan, L.K.; Zhang, W.; Li, G. Identification of Host Adaptation Genes in Extraintestinal Pathogenic Escherichia coli during Infection in Different Hosts. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef]
- Forsyth, V.S.; Himpsl, S.D.; Smith, S.N.; Sarkissian, C.A.; Mike, L.A.; Stocki, J.A.; Sintsova, A.; Alteri, C.J.; Mobley, H.L.T. Optimization of an Experimental Vaccine To Prevent Escherichia coli Urinary Tract Infection. mBio 2020, 11, e00555-20. [Google Scholar] [CrossRef]
- Linciano, P.; Cavalloro, V.; Martino, E.; Kirchmair, J.; Listro, R.; Rossi, D.; Collina, S. Tackling Antimicrobial Resistance with Small Molecules Targeting LsrK: Challenges and Opportunities. J. Med. Chem. 2020, 63, 15243–15257. [Google Scholar] [CrossRef] [PubMed]
- Murima, P.; McKinney, J.D.; Pethe, K. Targeting Bacterial Central Metabolism for Drug Development. Chem. Biol. 2014, 21, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.C.; Finlay, B.B. Targeting the type III secretion system to treat bacterial infections. Expert Opin. Ther. Targets 2014, 18, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, S.L.; Stegger, M.; Kudirkiene, E.; Lilje, B.; Poulsen, L.L.; Ronco, T.; Pires Dos Santos, T.; Kiil, K.; Bisgaard, M.; Pedersen, K.; et al. Diversity and Population Overlap between Avian and Human Escherichia coli Belonging to Sequence Type 95. mSphere 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafi, S.; Rahimi, M.; Nikousefat, Z. Extra-intestinal pathogenic Escherichia coli from human and avian origin: Detection of the most common virulence-encoding genes. Vet. Res. Forum 2019, 10, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Zhuge, X.; Jiang, M.; Tang, F.; Sun, Y.; Ji, Y.; Xue, F.; Ren, J.; Zhu, W.; Dai, J. Avian-source mcr-1-positive Escherichia coli is phylogenetically diverse and shares virulence characteristics with E. coli causing human extra-intestinal infections. Vet. Microbiol. 2019, 239, 108483. [Google Scholar] [CrossRef]
- Cunha, M.P.V.; Saidenberg, A.B.; Moreno, A.M.; Ferreira, A.J.P.; Vieira, M.A.M.; Gomes, T.A.T.; Knobl, T. Pandemic extra-intestinal pathogenic Escherichia coli (ExPEC) clonal group O6-B2-ST73 as a cause of avian colibacillosis in Brazil. PLoS ONE 2017, 12, e0178970. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Porter, S.B.; Johnston, B.; Thuras, P.; Clock, S.; Crupain, M.; Rangan, U. Extraintestinal Pathogenic and Antimicrobial-Resistant Escherichia coli, Including Sequence Type 131 (ST131), from Retail Chicken Breasts in the United States in 2013. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, N.M.; Johnson, J.R.; Johnston, B.; Curtiss, R., 3rd; Mellata, M. Zoonotic potential of Escherichia coli isolates from retail chicken meat products and eggs. Appl. Environ. Microbiol. 2015, 81, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, L.; Spangholm, D.J.; Pedersen, K.; Jensen, L.B.; Emborg, H.D.; Agerso, Y.; Aarestrup, F.M.; Hammerum, A.M.; Frimodt-Moller, N. Broiler chickens, broiler chicken meat, pigs and pork as sources of ExPEC related virulence genes and resistance in Escherichia coli isolates from community-dwelling humans and UTI patients. Int. J. Food Microbiol. 2010, 142, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Li, G.; Wilking, H.; Kiessling, S.; Alt, K.; Antao, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Zhu Ge, X.; Jiang, J.; Pan, Z.; Hu, L.; Wang, S.; Wang, H.; Leung, F.C.; Dai, J.; Fan, H. Comparative genomic analysis shows that avian pathogenic Escherichia coli isolate IMT5155 (O2:K1:H5; ST complex 95, ST140) shares close relationship with ST95 APEC O1:K1 and human ExPEC O18:K1 strains. PLoS ONE 2014, 9, e112048. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.; Johnson, S.J.; Stell, A.L.; Doetkott, C.; Johnson, J.R.; Kim, K.S.; Spanjaard, L.; Nolan, L.K. Comparison of extraintestinal pathogenic Escherichia coli strains from human and avian sources reveals a mixed subset representing potential zoonotic pathogens. Appl. Environ. Microbiol. 2008, 74, 7043–7050. [Google Scholar] [CrossRef] [Green Version]
- Mora, A.; Lopez, C.; Dabhi, G.; Blanco, M.; Blanco, J.E.; Alonso, M.P.; Herrera, A.; Mamani, R.; Bonacorsi, S.; Moulin-Schouleur, M.; et al. Extraintestinal pathogenic Escherichia coli O1:K1:H7/NM from human and avian origin: Detection of clonal groups B2 ST95 and D ST59 with different host distribution. BMC Microbiol. 2009, 9, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulin-Schouleur, M.; Reperant, M.; Laurent, S.; Bree, A.; Mignon-Grasteau, S.; Germon, P.; Rasschaert, D.; Schouler, C. Extraintestinal pathogenic Escherichia coli strains of avian and human origin: Link between phylogenetic relationships and common virulence patterns. J. Clin. Microbiol. 2007, 45, 3366–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Kariyawasam, S.; Wannemuehler, Y.; Mangiamele, P.; Johnson, S.J.; Doetkott, C.; Skyberg, J.A.; Lynne, A.M.; Johnson, J.R.; Nolan, L.K. The genome sequence of avian pathogenic Escherichia coli strain O1:K1:H7 shares strong similarities with human extraintestinal pathogenic E. coli genomes. J. Bacteriol. 2007, 189, 3228–3236. [Google Scholar] [CrossRef] [Green Version]
- Moulin-Schouleur, M.; Schouler, C.; Tailliez, P.; Kao, M.R.; Bree, A.; Germon, P.; Oswald, E.; Mainil, J.; Blanco, M.; Blanco, J. Common virulence factors and genetic relationships between O18:K1:H7 Escherichia coli isolates of human and avian origin. J. Clin. Microbiol. 2006, 44, 3484–3492. [Google Scholar] [CrossRef] [Green Version]
- Maluta, R.P.; Logue, C.M.; Casas, M.R.; Meng, T.; Guastalli, E.A.; Rojas, T.C.; Montelli, A.C.; Sadatsune, T.; de Carvalho Ramos, M.; Nolan, L.K.; et al. Overlapped sequence types (STs) and serogroups of avian pathogenic (APEC) and human extra-intestinal pathogenic (ExPEC) Escherichia coli isolated in Brazil. PLoS ONE 2014, 9, e105016. [Google Scholar] [CrossRef]
- Danzeisen, J.L.; Wannemuehler, Y.; Nolan, L.K.; Johnson, T.J. Comparison of multilocus sequence analysis and virulence genotyping of Escherichia coli from live birds, retail poultry meat, and human extraintestinal infection. Avian Dis. 2013, 57, 104–108. [Google Scholar] [CrossRef]
- Bergeron, C.R.; Prussing, C.; Boerlin, P.; Daignault, D.; Dutil, L.; Reid-Smith, R.J.; Zhanel, G.G.; Manges, A.R. Chicken as reservoir for extraintestinal pathogenic Escherichia coli in humans, Canada. Emerg. Infect. Dis. 2012, 18, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.; Viso, S.; Lopez, C.; Alonso, M.P.; Garcia-Garrote, F.; Dabhi, G.; Mamani, R.; Herrera, A.; Marzoa, J.; Blanco, M.; et al. Poultry as reservoir for extraintestinal pathogenic Escherichia coli O45:K1:H7-B2-ST95 in humans. Vet. Microbiol. 2013, 167, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Peigne, C.; Bidet, P.; Mahjoub-Messai, F.; Plainvert, C.; Barbe, V.; Medigue, C.; Frapy, E.; Nassif, X.; Denamur, E.; Bingen, E.; et al. The plasmid of Escherichia coli strain S88 (O45:K1:H7) that causes neonatal meningitis is closely related to avian pathogenic E. coli plasmids and is associated with high-level bacteremia in a neonatal rat meningitis model. Infect. Immun. 2009, 77, 2272–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giufre, M.; Graziani, C.; Accogli, M.; Luzzi, I.; Busani, L.; Cerquetti, M. Escherichia coli of human and avian origin: Detection of clonal groups associated with fluoroquinolone and multidrug resistance in Italy. J. Antimicrob. Chemother. 2012, 67, 860–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Fakhr, M.K.; Nolan, L.K. Comparison of Escherichia coli isolates implicated in human urinary tract infection and avian colibacillosis. Microbiology 2005, 151, 2097–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellata, M.; Johnson, J.R.; Curtiss, R., 3rd. Escherichia coli isolates from commercial chicken meat and eggs cause sepsis, meningitis and urinary tract infection in rodent models of human infections. Zoonoses Public Health 2018, 65, 103–113. [Google Scholar] [CrossRef]
- Stromberg, Z.R.; Johnson, J.R.; Fairbrother, J.M.; Kilbourne, J.; Van Goor, A.; Curtiss, R.R.; Mellata, M. Evaluation of Escherichia coli isolates from healthy chickens to determine their potential risk to poultry and human health. PLoS ONE 2017, 12, e0180599. [Google Scholar] [CrossRef] [Green Version]
- Nandanwar, N.; Janssen, T.; Kuhl, M.; Ahmed, N.; Ewers, C.; Wieler, L.H. Extraintestinal pathogenic Escherichia coli (ExPEC) of human and avian origin belonging to sequence type complex 95 (STC95) portray indistinguishable virulence features. Int. J. Med. Microbiol. 2014, 304, 835–842. [Google Scholar] [CrossRef]
- Krishnan, S.; Chang, A.C.; Hodges, J.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Nicholson, B.A.; Nolan, L.K.; Prasadarao, N.V. Serotype O18 avian pathogenic and neonatal meningitis Escherichia coli strains employ similar pathogenic strategies for the onset of meningitis. Virulence 2015, 6, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.; Johansen, A.E.; Thofner, I.; Christensen, J.P.; Pors, S.E.; Fresno, A.H.; Moller-Jensen, J.; Olsen, J.E. Infectious potential of human derived uropathogenic Escherichia coli UTI89 in the reproductive tract of laying hens. Vet. Microbiol. 2019, 239, 108445. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, D.; Ye, Z.; Zhu, X.; Yang, W.; Dong, L.; Gao, S.; Liu, X. Virulence traits and pathogenicity of uropathogenic Escherichia coli isolates with common and uncommon O serotypes. Microb. Pathog. 2017, 104, 217–224. [Google Scholar] [CrossRef]
- Meena, P.R.; Yadav, P.; Hemlata, H.; Tejavath, K.K.; Singh, A.P. Poultry-origin extraintestinal Escherichia coli strains carrying the traits associated with urinary tract infection, sepsis, meningitis and avian colibacillosis in India. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, S.; Huan, H.; Xu, X.; Zhu, X.; Yang, W.; Gao, Q.; Liu, X. Comparison of virulence factors and expression of specific genes between uropathogenic Escherichia coli and avian pathogenic E. coli in a murine urinary tract infection model and a chicken challenge model. Microbiology 2009, 155, 1634–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, H.; Bachmeier, J.; Bisgaard, M. New strategies to prevent and control avian pathogenic Escherichia coli (APEC). Avian Pathol. 2020, 1–30. [Google Scholar] [CrossRef]
- Dhaouadi, S.; Soufi, L.; Hamza, A.; Fedida, D.; Zied, C.; Awadhi, E.; Mtibaa, M.; Hassen, B.; Cherif, A.; Torres, C.; et al. Co-occurrence of mcr-1 mediated colistin resistance and beta-lactamases encoding genes in Multidrug-resistant Escherichia coli from broiler chickens with colibacillosis in Tunisia. J. Glob. Antimicrob. Resist. 2020. [Google Scholar] [CrossRef]
- Kim, Y.B.; Yoon, M.Y.; Ha, J.S.; Seo, K.W.; Noh, E.B.; Son, S.H.; Lee, Y.J. Molecular characterization of avian pathogenic Escherichia coli from broiler chickens with colibacillosis. Poult. Sci. 2020, 99, 1088–1095. [Google Scholar] [CrossRef]
- Meguenni, N.; Chanteloup, N.; Tourtereau, A.; Ahmed, C.A.; Bounar-Kechih, S.; Schouler, C. Virulence and antibiotic resistance profile of avian Escherichia coli strains isolated from colibacillosis lesions in central of Algeria. Vet. World 2019, 12, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Sarba, E.J.; Kelbesa, K.A.; Bayu, M.D.; Gebremedhin, E.Z.; Borena, B.M.; Teshale, A. Identification and antimicrobial susceptibility profile of Escherichia coli isolated from backyard chicken in and around ambo, Central Ethiopia. BMC Vet. Res. 2019, 15, 85. [Google Scholar] [CrossRef] [PubMed]
- Kurnia, R.S.; Indrawati, A.; Mayasari, N.; Priadi, A. Molecular detection of genes encoding resistance to tetracycline and determination of plasmid-mediated resistance to quinolones in avian pathogenic Escherichia coli in Sukabumi, Indonesia. Vet. World 2018, 11, 1581–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amer, M.M.; Mekky, H.M.; Amer, A.M.; Fedawy, H.S. Antimicrobial resistance genes in pathogenic Escherichia coli isolated from diseased broiler chickens in Egypt and their relationship with the phenotypic resistance characteristics. Vet. World 2018, 11, 1082–1088. [Google Scholar] [CrossRef]
- Gao, J.; Duan, X.; Li, X.; Cao, H.; Wang, Y.; Zheng, S.J. Emerging of a highly pathogenic and multi-drug resistant strain of Escherichia coli causing an outbreak of colibacillosis in chickens. Infect. Genet. Evol. 2018, 65, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Vounba, P.; Kane, Y.; Ndiaye, C.; Arsenault, J.; Fairbrother, J.M.; Bada Alambedji, R. Molecular Characterization of Escherichia coli Isolated from Chickens with Colibacillosis in Senegal. Foodborne Pathog. Dis. 2018, 15, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Hoepers, P.G.; Silva, P.L.; Rossi, D.A.; Valadares Junior, E.C.; Ferreira, B.C.; Zuffo, J.P.; Koerich, P.K.; Fonseca, B.B. The association between extended spectrum beta-lactamase (ESBL) and ampicillin C (AmpC) beta-lactamase genes with multidrug resistance in Escherichia coli isolates recovered from turkeys in Brazil. Br. Poult. Sci. 2018, 59, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Subedi, M.; Luitel, H.; Devkota, B.; Bhattarai, R.K.; Phuyal, S.; Panthi, P.; Shrestha, A.; Chaudhary, D.K. Antibiotic resistance pattern and virulence genes content in avian pathogenic Escherichia coli (APEC) from broiler chickens in Chitwan, Nepal. BMC Vet. Res. 2018, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Ehsan, I.; Sajjad Ur, R.; Saleemi, M.K.; Javed, M.R.; Mohsin, M. Detection of the colistin resistance gene mcr-1 in avian pathogenic Escherichia coli in Pakistan. J. Glob. Antimicrob. Resist. 2017, 11, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Younis, G.; Awad, A.; Mohamed, N. Phenotypic and genotypic characterization of antimicrobial susceptibility of avian pathogenic Escherichia coli isolated from broiler chickens. Vet. World 2017, 10, 1167–1172. [Google Scholar] [CrossRef] [Green Version]
- Yassin, A.K.; Gong, J.; Kelly, P.; Lu, G.; Guardabassi, L.; Wei, L.; Han, X.; Qiu, H.; Price, S.; Cheng, D.; et al. Antimicrobial resistance in clinical Escherichia coli isolates from poultry and livestock, China. PLoS ONE 2017, 12, e0185326. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, H.; Matsuoka, Y.; Nakagawa, E.; Murase, T. Characteristics of Escherichia coli isolated from broiler chickens with colibacillosis in commercial farms from a common hatchery. Poult. Sci. 2017, 96, 3717–3724. [Google Scholar] [CrossRef]
- Lima Barbieri, N.; Nielsen, D.W.; Wannemuehler, Y.; Cavender, T.; Hussein, A.; Yan, S.G.; Nolan, L.K.; Logue, C.M. mcr-1 identified in Avian Pathogenic Escherichia coli (APEC). PLoS ONE 2017, 12, e0172997. [Google Scholar] [CrossRef] [Green Version]
- Perreten, V.; Strauss, C.; Collaud, A.; Gerber, D. Colistin Resistance Gene mcr-1 in Avian-Pathogenic Escherichia coli in South Africa. Antimicrob Agents Chemother 2016, 60, 4414–4415. [Google Scholar] [CrossRef] [Green Version]
- Sola-Gines, M.; Cameron-Veas, K.; Badiola, I.; Dolz, R.; Majo, N.; Dahbi, G.; Viso, S.; Mora, A.; Blanco, J.; Piedra-Carrasco, N.; et al. Diversity of Multi-Drug Resistant Avian Pathogenic Escherichia coli (APEC) Causing Outbreaks of Colibacillosis in Broilers during 2012 in Spain. PLoS ONE 2015, 10, e0143191. [Google Scholar] [CrossRef] [Green Version]
- Cavicchio, L.; Dotto, G.; Giacomelli, M.; Giovanardi, D.; Grilli, G.; Franciosini, M.P.; Trocino, A.; Piccirillo, A. Class 1 and class 2 integrons in avian pathogenic Escherichia coli from poultry in Italy. Poult. Sci. 2015, 94, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Hornsey, M.; Betts, J.W.; Mehat, J.W.; Wareham, D.W.; van Vliet, A.H.M.; Woodward, M.J.; La Ragione, R.M. Characterization of a colistin-resistant Avian Pathogenic Escherichia coli ST69 isolate recovered from a broiler chicken in Germany. J. Med. Microbiol. 2019, 68, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Bourely, C.; Chauvin, C.; Jouy, E.; Cazeau, G.; Jarrige, N.; Leblond, A.; Gay, E. Comparative epidemiology of E. coli resistance to third-generation cephalosporins in diseased food-producing animals. Vet. Microbiol. 2018, 223, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Halfaoui, Z.; Menoueri, N.M.; Bendali, L.M. Serogrouping and antibiotic resistance of Escherichia coli isolated from broiler chicken with colibacillosis in center of Algeria. Vet. World 2017, 10, 830–835. [Google Scholar] [CrossRef] [Green Version]
- Ievy, S.; Islam, M.S.; Sobur, M.A.; Talukder, M.; Rahman, M.B.; Khan, M.F.R.; Rahman, M.T. Molecular Detection of Avian Pathogenic Escherichia coli (APEC) for the First Time in Layer Farms in Bangladesh and Their Antibiotic Resistance Patterns. Microorganisms 2020, 8, 1021. [Google Scholar] [CrossRef] [PubMed]
- Saidenberg, A.B.S.; Stegger, M.; Price, L.B.; Johannesen, T.B.; Aziz, M.; Cunha, M.P.V.; Moreno, A.M.; Knöbl, T. mcr-Positive Escherichia coli ST131-H22 from Poultry in Brazil. Emerg. Infect. Dis. 2020, 26, 1951–1954. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Wu, X.; Huo, S. Molecular characterization of multidrug-resistant avian pathogenic Escherichia coli isolated from septicemic broilers. Poult. Sci. 2015, 94, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Koutsianos, D.; Athanasiou, L.V.; Dimitriou, T.; Nikolaidis, M.; Tsadila, C.; Amoutzias, G.; Mossialos, D.; Koutoulis, K.C. Antibiotic Resistance Patterns and mcr-1 Detection in Avian Pathogenic Escherichia coli Isolates from Commercial Layer and Layer Breeder Flocks Demonstrating Colibacillosis in Greece. Microb. Drug Resist. 2020. [Google Scholar] [CrossRef]
- Bista, S.; Thapa Shrestha, U.; Dhungel, B.; Koirala, P.; Gompo, T.R.; Shrestha, N.; Adhikari, N.; Joshi, D.R.; Banjara, M.R.; Adhikari, B.; et al. Detection of Plasmid-Mediated Colistin Resistant mcr-1 Gene in Escherichia coli Isolated from Infected Chicken Livers in Nepal. Animals 2020, 10, 2060. [Google Scholar] [CrossRef]
- Temmerman, R.; Garmyn, A.; Antonissen, G.; Vanantwerpen, G.; Vanrobaeys, M.; Haesebrouck, F.; Devreese, M. Evaluation of Fluoroquinolone Resistance in Clinical Avian Pathogenic Escherichia coli Isolates from Flanders (Belgium). Antibiotics 2020, 9, 800. [Google Scholar] [CrossRef]
- Osman, K.M.; Kappell, A.D.; Elhadidy, M.; ElMougy, F.; El-Ghany, W.A.A.; Orabi, A.; Mubarak, A.S.; Dawoud, T.M.; Hemeg, H.A.; Moussa, I.M.I.; et al. Poultry hatcheries as potential reservoirs for antimicrobial-resistant Escherichia coli: A risk to public health and food safety. Sci. Rep. 2018, 8, 5859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- Redweik, G.A.J.; Stromberg, Z.R.; Van Goor, A.; Mellata, M. Protection against avian pathogenic Escherichia coli and Salmonella Kentucky exhibited in chickens given both probiotics and live Salmonella vaccine. Poult. Sci. 2020, 99, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, K.; Kong, Q.; Liu, Q. Immunization with outer membrane vesicles of avian pathogenic Escherichia coli O78 induces protective immunity in chickens. Vet. Microbiol. 2019, 236, 108367. [Google Scholar] [CrossRef]
- Hoseini Shahidi, R.; Hashemi Tabar, G.; Bassami, M.R.; Jamshidi, A.; Dehghani, H. The design and application of a bacterial ghost vaccine to evaluate immune response and defense against avian pathogenic Escherichia coli O2:K1 serotype. Res. Vet. Sci. 2019, 125, 153–161. [Google Scholar] [CrossRef]
- Hu, J.; Zuo, J.; Chen, Z.; Fu, L.; Lv, X.; Hu, S.; Shi, X.; Jing, Y.; Wang, Y.; Wang, Z.; et al. Use of a modified bacterial ghost lysis system for the construction of an inactivated avian pathogenic Escherichia coli vaccine candidate. Vet. Microbiol. 2019, 229, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi-Nik, H.; Bassami, M.R.; Mohri, M.; Rad, M.; Khan, M.I. Bacterial ghost of avian pathogenic E. coli (APEC) serotype O78:K80 as a homologous vaccine against avian colibacillosis. PLoS ONE 2018, 13, e0194888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Liu, Q.; Willias, S.; Liang, K.; Li, P.; Cheng, A.; Kong, Q. A bivalent vaccine derived from attenuated Salmonella expressing O-antigen polysaccharide provides protection against avian pathogenic Escherichia coli O1 and O2 infection. Vaccine 2018, 36, 1038–1046. [Google Scholar] [CrossRef]
- Van Goor, A.; Stromberg, Z.R.; Mellata, M. A recombinant multi-antigen vaccine with broad protection potential against avian pathogenic Escherichia coli. PLoS ONE 2017, 12, e0183929. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Chaudhari, A.A.; Oh, I.G.; Eo, S.K.; Park, S.Y.; Jawale, C.V. Immune responses to oral vaccination with Salmonella-delivered avian pathogenic Escherichia coli antigens and protective efficacy against colibacillosis. Can. J. Vet. Res. 2015, 79, 229–234. [Google Scholar]
- Sadeyen, J.R.; Wu, Z.; Davies, H.; van Diemen, P.M.; Milicic, A.; La Ragione, R.M.; Kaiser, P.; Stevens, M.P.; Dziva, F. Immune responses associated with homologous protection conferred by commercial vaccines for control of avian pathogenic Escherichia coli in turkeys. Vet. Res. 2015, 46, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, K.M.; Browning, G.F.; Noormohammadi, A.H.; Markham, P.; Marenda, M.S. Avian pathogenic Escherichia coli DeltatonB mutants are safe and protective live-attenuated vaccine candidates. Vet. Microbiol. 2014, 173, 289–298. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Woodward, M.J.; Kumar, M.; Rodenberg, J.; Fan, H.; Wales, A.D.; Karaca, K. Efficacy of a live attenuated Escherichia coli O78:K80 vaccine in chickens and turkeys. Avian Dis. 2013, 57, 273–279. [Google Scholar] [CrossRef]
- Nagano, T.; Kitahara, R.; Nagai, S. An attenuated mutant of avian pathogenic Escherichia coli serovar O78: A possible live vaccine strain for prevention of avian colibacillosis. Microbiol. Immunol. 2012, 56, 605–612. [Google Scholar] [CrossRef]
- Yaguchi, K.; Ohgitani, T.; Noro, T.; Kaneshige, T.; Shimizu, Y. Vaccination of chickens with liposomal inactivated avian pathogenic Escherichia coli (APEC) vaccine by eye drop or coarse spray administration. Avian Dis. 2009, 53, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Lynne, A.M.; Kariyawasam, S.; Wannemuehler, Y.; Johnson, T.J.; Johnson, S.J.; Sinha, A.S.; Lynne, D.K.; Moon, H.W.; Jordan, D.M.; Logue, C.M.; et al. Recombinant Iss as a potential vaccine for avian colibacillosis. Avian Dis. 2012, 56, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, T.Z.; Tabatabaei, S.; Karimi, V.; Fasaei, B.N.; Derakhshandeh, A.; Jahromi, A.O.N. Assessment of immunity against avian colibacillosis induced by an aroA mutant containing increased serum survival gene in broilers. Braz. J. Microbiol. 2012, 43, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Fernandes Filho, T.; Favaro, C., Jr.; Ingberman, M.; Beirao, B.C.; Inoue, A.; Gomes, L.; Caron, L.F. Effect of spray Escherichia coli vaccine on the immunity of poultry. Avian Dis. 2013, 57, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, Q.; Yi, J.; Liang, K.; Wei, Y.; Kong, Q. A biologically conjugated polysaccharide vaccine delivered by attenuated Salmonella Typhimurium provides protection against challenge of avian pathogenic Escherichia coli O1 infection. Pathog. Dis. 2017, 75. [Google Scholar] [CrossRef]
- Vandemaele, F.; Bleyen, N.; Abuaboud, O.; vanderMeer, E.; Jacobs, A.; Goddeeris, B.M. Immunization with the biologically active lectin domain of PapGII induces strong adhesion-inhibiting antibody responses but not protection against avian pathogenic Escherichia coli. Avian Pathol. 2006, 35, 238–249. [Google Scholar] [CrossRef]
- Vandemaele, F.; Ververken, C.; Bleyen, N.; Geys, J.; D’Hulst, C.; Addwebi, T.; van Empel, P.; Goddeeris, B.M. Immunization with the binding domain of FimH, the adhesin of type 1 fimbriae, does not protect chickens against avian pathogenic Escherichia coli. Avian Pathol. 2005, 34, 264–272. [Google Scholar] [CrossRef]
- Roland, K.; Karaca, K.; Sizemore, D. Expression of Escherichia coli antigens in Salmonella typhimurium as a vaccine to prevent airsacculitis in chickens. Avian Dis. 2004, 48, 595–605. [Google Scholar] [CrossRef]
- Lynne, A.M.; Foley, S.L.; Nolan, L.K. Immune response to recombinant Escherichia coli Iss protein in poultry. Avian Dis. 2006, 50, 273–276. [Google Scholar] [CrossRef] [Green Version]
- Kariyawasam, S.; Wilkie, B.N.; Gyles, C.L. Construction, characterization, and evaluation of the vaccine potential of three genetically defined mutants of avian pathogenic Escherichia coli. Avian Dis. 2004, 48, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, S.; Wilkie, B.N.; Hunter, D.B.; Gyles, C.L. Systemic and mucosal antibody responses to selected cell surface antigens of avian pathogenic Escherichia coli in experimentally infected chickens. Avian Dis. 2002, 46, 668–678. [Google Scholar] [CrossRef]
- Tuntufye, H.N.; Ons, E.; Pham, A.D.; Luyten, T.; Van Gerven, N.; Bleyen, N.; Goddeeris, B.M. Escherichia coli ghosts or live E. coli expressing the ferri-siderophore receptors FepA, FhuE, IroN and IutA do not protect broiler chickens against avian pathogenic E. coli (APEC). Vet. Microbiol. 2012, 159, 470–478. [Google Scholar] [CrossRef]
- Ma, S.T.; Ding, G.J.; Huang, X.W.; Wang, Z.W.; Wang, L.; Yu, M.L.; Shi, W.; Jiang, Y.P.; Tang, L.J.; Xu, Y.G.; et al. Immunogenicity in chickens with orally administered recombinant chicken-borne Lactobacillus saerimneri expressing FimA and OmpC antigen of O78 avian pathogenic Escherichia coli. J. Med. Microbiol. 2018, 67, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Amoako, K.K.; Prysliak, T.; Potter, A.A.; Collinson, S.K.; Kay, W.W.; Allan, B.J. Attenuation of an avian pathogenic Escherichia coli strain due to a mutation in the rpsL gene. Avian Dis. 2004, 48, 19–25. [Google Scholar] [CrossRef]
- Hu, R.; Li, J.; Zhao, Y.; Lin, H.; Liang, L.; Wang, M.; Liu, H.; Min, Y.; Gao, Y.; Yang, M. Exploiting bacterial outer membrane vesicles as a cross-protective vaccine candidate against avian pathogenic Escherichia coli (APEC). Microb. Cell Fact. 2020, 19, 119. [Google Scholar] [CrossRef]
- Koutsianos, D.; Gantelet, H.; Franzo, G.; Lecoupeur, M.; Thibault, E.; Cecchinato, M.; Koutoulis, K.C. An Assessment of the Level of Protection Against Colibacillosis Conferred by Several Autogenous and/or Commercial Vaccination Programs in Conventional Pullets upon Experimental Challenge. Vet. Sci. 2020, 7, 80. [Google Scholar] [CrossRef]
- Hu, R.; Liu, H.; Wang, M.; Li, J.; Lin, H.; Liang, M.; Gao, Y.; Yang, M. An OMV-Based Nanovaccine Confers Safety and Protection against Pathogenic Escherichia coli via Both Humoral and Predominantly Th1 Immune Responses in Poultry. Nanomaterials 2020, 10, 2293. [Google Scholar] [CrossRef]
- Soleymani, S.; Tavassoli, A.; Hashemi Tabar, G.; Kalidari, G.A.; Dehghani, H. Design, development, and evaluation of the efficacy of a nucleic acid-free version of a bacterial ghost candidate vaccine against avian pathogenic E. coli (APEC) O78:K80 serotype. Vet. Res. 2020, 51, 144. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, G.M.; ElZorkany, H.E.; Farroh, K.Y.; Abd El-Aziz, W.R.; Elshoky, H.A. Potential improvement of the immune response of chickens against E. coli vaccine by using two forms of chitosan nanoparticles. Int. J. Biol Macromol. 2020, 167, 395–404. [Google Scholar] [CrossRef]
- Cox, G.J.; Griffith, B.; Reed, M.; Sandstrom, J.D.; Peterson, M.P.; Emery, D.; Straub, D.E. A vaccine to prevent egg layer peritonitis in chickens. Avian Dis. 2020. [Google Scholar] [CrossRef]
- Huttner, A.; Gambillara, V. The development and early clinical testing of the ExPEC4V conjugate vaccine against uropathogenic Escherichia coli. Clin. Microbiol. Infect. 2018, 24, 1046–1050. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Wang, Y.; Yan, W.; Li, A.; Jiang, H.; Fang, J. Effects of Lactobacillus plantarum 15-1 and fructooligosaccharides on the response of broilers to pathogenic Escherichia coli O78 challenge. PLoS ONE 2019, 14, e0212079. [Google Scholar] [CrossRef] [Green Version]
- Tarabees, R.; Gafar, K.M.; El-Sayed, M.S.; Shehata, A.A.; Ahmed, M. Effects of Dietary Supplementation of Probiotic Mix and Prebiotic on Growth Performance, Cecal Microbiota Composition, and Protection Against Escherichia coli O78 in Broiler Chickens. Probiot. Antimicrob. Proteins 2019, 11, 981–989. [Google Scholar] [CrossRef]
- Tarabees, R.; El-Sayed, M.S.; Shehata, A.A.; Diab, M.S. Effects of the Probiotic Candidate E. faecalis-1, the Poulvac E. coli Vaccine, and their Combination on Growth Performance, Caecal Microbial Composition, Immune Response, and Protection against E. coli O78 Challenge in Broiler Chickens. Probiot. Antimicrob. Proteins 2020, 12, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Lin, T.-L.; Tsai, Y.-L.; Wu, T.-R.; Lai, W.-F.; Lu, C.-C.; Lai, H.-C. Next generation probiotics in disease amelioration. J. Food Drug Anal. 2019, 27, 615–622. [Google Scholar] [CrossRef]
- Naghizadeh, M.; Karimi Torshizi, M.A.; Rahimi, S.; Dalgaard, T.S. Synergistic effect of phage therapy using a cocktail rather than a single phage in the control of severe colibacillosis in quails. Poult. Sci. 2019, 98, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.; Sereno, R.; Azeredo, J. In vivo efficiency evaluation of a phage cocktail in controlling severe colibacillosis in confined conditions and experimental poultry houses. Vet. Microbiol. 2010, 146, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Huff, W.E.; Huff, G.R.; Rath, N.C.; Balog, J.M.; Donoghue, A.M. Alternatives to antibiotics: Utilization of bacteriophage to treat colibacillosis and prevent foodborne pathogens. Poult. Sci. 2005, 84, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, X.; Shi, W.; Qian, Z.; Wang, Y. Sensitization of avian pathogenic Escherichia coli to amoxicillin in vitro and in vivo in the presence of surfactin. PLoS ONE 2019, 14, e0222413. [Google Scholar] [CrossRef] [PubMed]
- Goonewardene, K.; Ahmed, K.A.; Gunawardana, T.; Popowich, S.; Kurukulasuriya, S.; Karunarathna, R.; Gupta, A.; Ayalew, L.E.; Lockerbie, B.; Foldvari, M.; et al. Mucosal delivery of CpG-ODN mimicking bacterial DNA via the intrapulmonary route induces systemic antimicrobial immune responses in neonatal chicks. Sci. Rep. 2020, 10, 5343. [Google Scholar] [CrossRef]
- Peng, L.Y.; Yuan, M.; Song, K.; Yu, J.L.; Li, J.H.; Huang, J.N.; Yi, P.F.; Fu, B.D.; Shen, H.Q. Baicalin alleviated APEC-induced acute lung injury in chicken by inhibiting NF-kappaB pathway activation. Int. Immunopharmacol. 2019, 72, 467–472. [Google Scholar] [CrossRef]
- Cuperus, T.; van Dijk, A.; Matthijs, M.G.; Veldhuizen, E.J.; Haagsman, H.P. Protective effect of in ovo treatment with the chicken cathelicidin analog D-CATH-2 against avian pathogenic E. coli. Sci. Rep. 2016, 6, 26622. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, L.Y.; Wu, S.C.; Xia, F.; Fu, Y.X.; Wu, Y.L.; Leng, C.Q.; Yi, P.F.; Shen, H.Q.; Wei, X.B.; et al. Andrographolide interferes quorum sensing to reduce cell damage caused by avian pathogenic Escherichia coli. Vet. Microbiol. 2014, 174, 496–503. [Google Scholar] [CrossRef]
- Kathayat, D.; Helmy, Y.A.; Deblais, L.; Rajashekara, G. Novel small molecules affecting cell membrane as potential therapeutics for avian pathogenic Escherichia coli. Sci. Rep. 2018, 8, 15329. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Deblais, L.; Kassem, I.I.; Kathayat, D.; Rajashekara, G. Novel small molecule modulators of quorum sensing in avian pathogenic Escherichia coli (APEC). Virulence 2018, 9, 1640–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totsika, M. Disarming pathogens: Benefits and challenges of antimicrobials that target bacterial virulence instead of growth and viability. Future Med. Chem. 2017, 9, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Belete, T.M. Novel targets to develop new antibacterial agents and novel alternatives to antibacterial agents. Hum. Microbiome J. 2019, 11, 100052. [Google Scholar] [CrossRef]
- Kathayat, D.; Antony, L.; Deblais, L.; Helmy, Y.A.; Scaria, J.; Rajashekara, G. Small Molecule Adjuvants Potentiate Colistin Activity and Attenuate Resistance Development in Escherichia coli by Affecting pmrAB System. Infect. Drug Resist. 2020, 13, 2205–2222. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M. Antimicrobial peptide, cLF36, affects performance and intestinal morphology, microflora, junctional proteins, and immune cells in broilers challenged with E. coli. Sci. Rep. 2019, 9, 14176. [Google Scholar] [CrossRef]


{kind=link}
| Virulence Factors | Genes/Proteins Involved | Role in Pathogenesis/Infection | Reference |
|---|---|---|---|
| Adhesins | fimH, fimC, papA, papC, papEF, papG I, papG II, papGIII, felA, sfa/sfaS, afaIBC, focGE, lpfA, lpf0141, lpf0154, flgE, crl, csg, bmaE, tsh, mat/ecpA, hra/hrlA/hek, iha, yqiG, kii | Adhesion, colonization, biofilm formation, motility, intracellular survival | [32,33,34,35,36,37,38,39,41] |
| yfc O | Adhesion, colonization, resistance to environmental stresses | [42] | |
| yad C | Adhesion, intracellular survival, motility | [43] | |
| aat A, aatB, upaB | Adhesion, colonization, biofilm formation | [44,45] | |
| fdtA, rluD, yjhB, ecpR, fdeC | Adhesion | [46] | |
| Invasins | ibeA, ibeB, tia, gimB | Invasion, resistance to oxidative stress, colonization, proliferation, biofilm formation | [35,47,48] |
| IbeR | Invasion, resistance to serum and environmental stresses, expression of virulence genes | [49] | |
| ych O | Motility, adhesion, invasion, biofilm formation, expression of membrane proteins and metabolism genes | [50] | |
| Iron acquisition systems | iutA, iucC, iucD, aerJ, iucA, iucB, iroBCDEN, fyuA, sitABCD, mntH, feoB, irp2, ireA, eitABCD, fepC, chuA, bfr | Iron and manganese uptake from the host, adhesion, invasion, colonization, persistence, expression of virulence genes, resistance to environmental stresses | [36,37,51,52,53,54,55,56,57] |
| entE, entS, tolC | Invasion, colonization, persistence | [60] | |
| Protectins | iss, traT, ompT, kpsMT(K1), kpsMT(II), kpsMT(III), neuC, neuS, neuD, kfiC-K5, betA | Protect from serum bactericidal activity and phagocytosis, adhesion, invasion, intracellular survival, colonization, proliferation | [36,41,51,53,61] |
| YbjX, PagP | Resistance to serum and environmental stresses, invasion, intracellular survival | [62,63] | |
| OmpA | Intracellular survival | [64] | |
| wzy | Adhesion, invasion, intracellular survival, colonization | [65] | |
| waa L | Motility, resistance to phagocytosis and environmental stresses, adhesion, invasion, biofilm formation | [65] | |
| sod A | Protect against ROS-mediated host defenses, biofilm formation | [68] | |
| lpx M | Invasion, intracellular survival, colonization, regulation of expression of cytokine genes and nitric oxide production | [67] | |
| Toxins | hlyF, hlyA, hlyE, cdtB, cdtS, vat, sat, stx2f, astA, pic, EAST-1, espC, ace4/35 | Cell lysis and damage, induce host cell vacuolization, colonization, motility, biofilm formation, agglutination, formation of outer membrane vesicles | [35,38,39,41,51,53,69,70,71,72,73] |
| Other virulence and pathogenesis factors | |||
| Quorum-sensing system (AI-2) | LuxS, LsrABCDFGK, ptsI, Pfs | Motility, biofilm formation, adherence, invasion, colonization, intracellular survival, persistence, expression of virulence genes, cell damage | [10,74,75,76,77] |
| Secretion systems | DotU, CpxRA, IcmF, Hcp, ClpV, VrgG (Type VI) | Interbacterial competition, adhesion, invasion, intracellular survival, colonization, motility, biofilm formation, production of type 1 fimbriae, resistance to serum bactericidal activity, modulation of intracellular host responses (IL-18, IL-1β) | [11,80,81,82,83,84,85] |
| EtrA, YqeI, EivC (Type III) | Motility, intracellular survival, resistance to phagocytosis and serum bactericidal activity, proliferation, expression of fimbriae genes, downregulation of pro-inflammatory cytokines | [12,78,79] | |
| Two-component systems | PhoPQ, tolC | Biofilm formation, motility, adhesion, invasion, intracellular survival, systemic infection, expression of virulence genes and genes associated with flagellar assembly, ABC transporters, quorum sensing, and bacterial chemotaxis | [13,86,90] |
| BasSR | Biofilm formation, APEC virulence and colonization in vivo | [91] | |
| KdpDE | Expression of flagella-related genes, flagellum formation, motility and resistance to serum bactericidal activity | [87] | |
| RstAB, hdeD | Iron acquisition, acid resistance, intracellular survival, colonization | [88,92] | |
| BarA-UvrY | Adhesion, invasion, persistence, intracellular survival, resistance to serum bactericidal activity and oxidative stress, regulation of exopolysaccharide production and type 1 and P fimbriae | [89] | |
| Transcriptional regulators | AutA/AutR | Expression of K1 capsule and acid resistance systems, adaptive lifestyle change | [14] |
| FNR | Adhesion, invasion, expression of type 1 fimbriae and type VII secretion system, resistance to oxidative stress | [15] | |
| YjjQ | Flagellar motility | [93] | |
| McbR | Biofilm formation, response to H2O2 | [94] | |
| tyrR | Invasion, motility, intracellular survival | [37] | |
| RfaH | Invasion, intracellular survival, resistance to serum bactericidal activity | [95] | |
| Metabolism-associated genes | acs -yjcH-actP | Intracellular survival, proliferation, colonization, production of pro-inflammatory cytokines and nitric oxide | [96] |
| PotE, PotF | Colonization, adhesion | [16] | |
| NirC | Adhesion, colonization | [17] | |
| ArcA | Chemotaxis, motility | [18] | |
| Miscellaneous | OmpF, OmpC | Adhesion, invasion, colonization, proliferation | [100] |
| Prophage phiv142-3 (orf20) and phiv205-1 | Resistance to serum and environmental stresses, adhesion, invasion, intracellular survival, colonization, biofilm formation, formation of flagella and I fimbriae | [97,98,99] | |
| YicS | Motility, biofilm formation, invasion | [101] | |
| cpd B | Colonization | [102] | |
| pst B | Resistance to serum bactericidal activity and oxidative stress, colonization | [103] | |
| tmRNA-SmpB | Colonization, persistence, replication, intracellular survival | [104] | |
| mli C | Resistance to serum bactericidal activity | [105] | |
| malX, frz, cvaABC, cvi, cba, cib/cibI, cbi, cma, eaeA, sopB, yfcV, gad, mchBCF, mcmA, bor, air, eilA, celB, pabB, capU, cif, tir, tccp, nleB, iaL, cjrC, mig-14p | Unknown/not clearly known functions | [34,39,40,41,53,54,57,106,107,108] | |
| Genes essential for systemic infections and adaptation | metH, lysA, pntA, purL, serS, ybjE, ycdK (rutC), wcaJ, gspL, sdsR, irp2, eitD, ylbE, yjiY, tkt1, pilN, pilQ, tsh, hpb, TcfD, Z5222, waaO, waaY, iutA, iucA, iucD, iroC, ColE2, traK, traG, traT, SopA, psiA, hkaG, hkbV, hkbQ, Z3370, Int, CC0532, TM0427, YPO3000, rhsH, RSp0733, bioABFCD, rnfA, rfnE, gene encoding endonuclease III, creABCD, yehD, potF, flgE, tyrR, bfr | Systemic APEC infections and adaptation | [37,109,110,111] |
| Study Location | Study Samples | Resistance Reported | Resistance Genes Reported | Reference |
|---|---|---|---|---|
| Belgium | Broilers | EFX | gyrA, parC, parE, PMQR | [172] |
| Nepal | Chickens | CLT, TTC, CFX, CXT, IPM | mcr-1 | [171] |
| Greece | Layers | CLT | mcr-1 | [170] |
| Bangladesh | Chickens | APC, DXC, TTC, NFN, CFX, NDA, CXT, IPM, GNC, CMC, SFN, AZT, PMB | [108] | |
| Brazil | Chickens | CLT | mcr-5, mcr-9 | [168] |
| Bangladesh | Layers | APC, TTC, CMC, EHC, EFX, CFX, SPM, CLT, GNC | [167] | |
| Thailand and Australia | Broilers and broiler breeders | AXC, CFU, CMC, EFX, FFC, GNC, NDA, TMP-SFM, TTC | gyrA, parC | [51] |
| Tunisia | Chicken’s fresh carcasses | NDA, FMQ, EFX, DXC, TTC, FFC, TMP-SFM, SPM, AXC, AXC-CVA; CFT, CFZ, ATN, CFP, CLT | mcr-1, blaCTX-M-1, blaTEM,blaSHV, tetA, dfraI, floR, cmlA, aadA, strA, strB, sul1 | [146] |
| Japan | Municipal wastewater Influents | CLT | mcr-1 | [107] |
| Egypt | Broilers | APC, AXC-CVA, TTC, CLT, DXC, SMC, FFC, CFT, CFX | [32] | |
| Korea | Broilers | APC, NDA, TTC, CPT, CPX | blaCTX-M-1, blaCTX-M-15,blaTEM-1,aac(3)-II, qnrA, qnrS | [147] |
| Germany | Broiler | CLT | mcr-1 | [164] |
| Algeria | Broilers | NDA, AXC, APC, TCC, PPA, TMP-SFM | [148] | |
| China | Chickens | APC, CFT, CMC, GNC, KMC, SPM, TMP-SFM, NDA, TTC | [53] | |
| Taiwan | Day-old hatchery chicks | APC, AXC, CPX, FFC, TMP-SFM | [70] | |
| Jordan | Broilers | TMP-SFM, FFC, AXC, DXC, SCC | int1, sul1, sul2, tetA, blaTEM | [71] |
| Italy | Broilers/turkey /goose/guinea fowl/duck/ pigeon/layers /capon | APC, NDA, AXC-CVA, CFT, CFX, CFN, CFZ, CMC, EFX, GNC, KMC, NDA, SPM, TTC, FMQ, TMP-SFM | [72] | |
| France | Broilers/ducks /turkeys/layers | CFU | [165] | |
| Ethiopia | Backyard chickens | APC, CFT, CXC, CRX, TTC | [149] | |
| Pakistan | Broilers | APC, TTC, CFX | [34] | |
| Indonesia | Chickens | TTC, OTC, CFX, CFX, NFX | tetA, tetB, qnrA, qnrB, qnrS | [150] |
| Canada | Broilers /Breeders | TTC, APC, SCC, GNC, CFU, KMC, APC, TMP-SFM | [54] | |
| Egypt | Broilers | OTC, KMC, APC, CDC, SPM, EFX, CMC, CFT, GNC, EHC, OXC, TMP-SFM | CITM, ere, aac(3)-(IV), tetA, tetB, dfr(A1), aad (A1) | [151] |
| China | Layer | CRX, TMC, MMC, CFN, SPM, APC | strA, strB, blaCMY-2, blaCTX-M-19, blaTEM-1B, fosA, mphA, floR, sul2, tetA, tetB | [152] |
| Senegal | Chickens | TTC, SSX, APC, TMP-SFM, SPM, CFU, CTX, CFX | gyrB, parC, blaCTX-M,blaOXA-1, blaTEM,blaSHV, tetA, dhrfVII | [153] |
| Algeria | Broilers | TTC, TMP-SFM, EFX, NDA, APC, DXC, CMC, AXC-CVA, FMQ | [166] | |
| Brazil | Turkeys | EHC, APC, TTC, OTC, LCC, SCC, TMP-SFM | blaCTX-M-2,blaCTX-M-8, blaCTX-M-8/25, blaTEM,blaSHV, blaCMY-2 | [154] |
| Nepal | Broilers | APC, CMX, DXC, CLT | [155] | |
| Egypt | Broilers | PNC, CFP, NFX, CMC | sul1, blaTEM,blaSHV,tetA, aadA | [157] |
| Pakistan | Broilers | CLT | mcr-1 | [156] |
| China | Chickens | TTC, NDA, TMP-SFM, APC, AKC, ATN, CFZ, CFT, CMC, CFX | [158] | |
| Japan | Broilers | APC, CFT, OTC, SPM, NDA, EFX, TMP, KMC, GNC, CMC | blaCTX-M-2,blaCTX-M-14, blaCTX-M-65, blaTEM-1,blaSHV-12, blaCMY-2, aacA4, aadA2, aadA5, catB3, dfrA1, dfrA17, parC, aac(6′)-Ib-cr, oqxAB | [159] |
| China/ Egypt | Chickens | CLT | mcr-1 | [160] |
| Brazil | Chickens | APC, TTC, GNC, NMC, EFX, NFX, TMP-SFM | [35] | |
| South Africa | Broilers | CLT | mcr-1 | [161] |
| Spain | Broilers | CFT, CFZ, XCT, NDA, CFX, TTC, KMC, SPM, TMP-SFM, GNC, APC, FFC | blaCTX-M-14, blaSHV-2, blaCMY-2, qnrA, aac(6′)-Ib-cr | [162] |
| China | Chickens | TTC, NDA, APC, SFZ, SPM, TMP, CMX | [73] | |
| Italy | Broilers/layers /turkeys | SPM, TMP | aadA dfrA | [163] |
| China | Broilers | APC, KMC, CFX, LFX, SPM, GNC, OFX, NFX, CTX | blaCTX-M-15, blaOXA-30, blaTEM, blaSHV-2, blaCMY-2, qnrA, qnrB, qnrS, strA, strB, aph(3′)-II1, aac(3)-IIa, aac(6′)-Ib, ant(3″)-Ia | [169] |
| Vaccine Description | Challenge | Route | Main Findings | Reference |
|---|---|---|---|---|
| Live-attenuated APEC O78 ΔaroA (Poulvac® E. coli) and autogenous (O18, O78, and O111) vaccine (alone and combination) | Layers (O78, O18, and O111) | Spray, I/M | -Combination of live- attenuated and autogenous vaccine significantly reduce the mortality, lesion scores, and APEC load -Live-attenuated vaccine alone does not provide significant protection | [202] |
| Live-attenuated Salmonella-delivered vaccine secreting APEC antigens (PapA, PapG, IutA, ClpG) | Layers (JOL718) | Oral | -High IgA, IFN- γ, IL-2, and IL-6 levels -Complete protection against JOL718 challenge -Less lesions in liver | [182] |
| Live-attenuated APEC O78 ΔaroA vaccine (Poulvac® E. coli) and formalin-inactivated APEC O78 bacterin | Turkeys (EC34195nal®, O78:K80) | Spray and S/C, respectively | -2 log10 CFU/g reduction in APEC load in spleen, liver, and kidney -No lesions -Induce transcription of IL-13 and TGF-β4 -Repress transcription of IFN- γ -Modulate CXCLi2 expression -Elevate IgA and IgY levels -Higher ability of splenocytes to proliferate | [183] |
| Live-attenuated APEC O78 ΔaroA vaccine (Poulvac® E. coli) | Broilers | Spray | -High CD4+TCRVβ1+ cells -High naïve and memory CD8 cells -Stimulate high state of immunocompetence -Stimulate production of IgA | [190] |
| Live-attenuated ΔtonB and ΔtonB/Δfur vaccine (E956) | Layers (E956) | Spray | -Less lesions In airsacs, heart, and liver | [184] |
| Siderophore receptor and porin (SRP®) APEC vaccine for egg layer peritonitis | Layers (O1, O2, and O78) | I/V, I/VAG, I/T, I/P | -Complete protection against mortality - Substantially reduce APEC colonization and lesions | [206] |
| Live-attenuated O78:K80 ΔaroA vaccine | Chickens and turkeys (O78 and X) | Spray or spray followed by oral booster | -Significant reduction in mortality -homologous and heterologous protection | [185] |
| Live-attenuated O78:K80 ΔaroA vaccine | Broilers (χ1378 and 02:K12) | Aerosol | -High weight gain -No antibody responses -No homologous and heterologous protection | [189] |
| Live-attenuated APEC O78:J29—Δcrp vaccine (AESN1331) | Layers (O78:J46) | Spray, eye drop, and in ovo | -Significant reduction in mortality -Less lesion scores in heart and liver | [186] |
| Live-attenuated S. typhimurium vaccine expressing O78 LPS and E. coli type 1 fimbriae | Layers (χ7122, χ7252, and χ7096) | Oral and spray | -Lower lesion scores in airsacs -No heterologous protection | [194] |
| ΔgalE, ΔpurA, and ΔaroA (APECO78:EC99) vaccine | Broilers (EC99 and EC317) | Spray | -High IgY level -Less lesion scores and APEC load -No heterologous protection | [196] |
| Live E. coli vaccine with rspL mutation (EC844) | Broilers (EC317) | Aerosol and oral | -Lower APEC lesions with three doses | [200] |
| Recombinant Salmonella vaccine strain S740 (pSS28) containing APEC O1 and APEC O2 O-antigens | Layers (O1 C24-2, O2 CE37) | Oral followed by I/M booster | -High IgG and IgA levels -73.33% and 66.67% protection against O2 CE37 and O1 C24-2 challenge, respectively -High opsonophagocytosis and serum bactericidal activity -Less lesions in liver and spleen | [180] |
| Recombinant antigen (rAg) vaccine containing ExPEC proteins (OmpA, OmpT, TraT, EtsC) | Layers (O2) | S/C | -Higher IgY, IL-1β, IL-6, IL-18, IFN-γ, IL-4, IFN-β, and IL-8 levels -Increased serum bactericidal activity against multiple APEC strains -Less lesions in airsacs, liver, and heart -Less APEC load in heart and liver | [181] |
| Recombinant attenuated S. Typhimurium vaccine (RASV) χ9373 | Layers χ7122 (O78:K80) | Oral | -High IgY level -Lower signs of airsacculitis -Lower lesion scores in heart and liver -Lower APEC load in blood -Increase blood and serum bactericidal activity against multiple APEC strains | [175] |
| Recombinant Salmonella vaccine strain S740 containing APEC O1 O-antigen (pSS27) | Layers (O1:C24-2) | Oral followed by and I/M booster | -High IgG and IgA levels ->50% protection against O1:C24-2 challenge | [191] |
| Recombinant iss vaccine | Broilers (O1, O2, and χ7122) | I/M | -High IgA and IgG levels -Less lesions in heart, liver, and airsacs -Heterologous protection | [188] |
| Recombinant iss vaccine | Layers (O2 and O78) | S/C | -High humoral response -Lower APEC lesions -Heterologous protection | [195] |
| E. coli BL21:D3 expressing FepA, FhuE, IroN, and IutA antigens | Broilers (CH2) | I/N | -High IgG level -No protection against CH2 challenge | [198] |
| Lactobacillus saerimineri expressing O78 FimC and OmpC antigens | Layers (O78) | Oral | -High IgG and IgA levels -Lower mortality | [199] |
| APEC O78, O1, and O2 outer membrane vesicles (OMVs) | Broilers (O78, O1, and O2) | I/M | -High IgG level -High IFN-γ, IL-17, and IL-10 levels ->90% protective efficacy -Reduce bacterial burden in liver and lung -No effect on growth performance | [201] |
| APEC O78 OMVs | Layers (O78) | I/M | -High IgG level -High opsonophagocytosis and serum bactericidal activity -Complete protection against O78 challenge | [176] |
| APEC O2 OMVs | Broilers (O2) | I/M | -Protective against homologous infection -Enhance specific (IgY) antibody response -Elicit IFN-γ mediated Th1 responses | [203] |
| Bacterial ghost vaccine (O2:K1) constructed using PhiX 174 lysis gene E | Broilers (O2:K1) | S/C, spray | -Less lesion scores in airsacs, liver, and heart -High IgG and sIgA levels -High IFN-γ | [177] |
| Bacterial ghost vaccine (O2: DE17ΔluxSΔaroA) constructed using PhiX 174 lysis gene E | Layers (O2: DE17) | S/C | -Complete protection against O2:DE17 challenge -Less protection against O2: CE35 challenge -No lesions in liver, heart, spleen, and lung -Higher IgG, IFN-γ, and TNF-α levels | [178] |
| Bacterial ghost vaccine (O78:K80) constructed using PhiX 174 lysis gene E | Broilers (O78:K80) | S/C, spray | -Reduce lesions in airsacs, liver, and heart -High IgY, IgA, and IFN-γ levels | [179] |
| Bacterial ghost vaccine (O78:K80) containing E-lysis and S nuclease genes | Broilers (O78:K80) | S/C, spray | -Lower lesion scores -High antibody (IgY and IgA) and IFN-γ levels -Increase proinflammatory (IL-6, IL-1 β, and TNFSF15) cytokines production | [204] |
| Chitosan nanoparticles containing APEC O1 and O78 outer membrane proteins (OMPs) and flagellar antigens | Broilers (O1 and O78) | S/C | -High antibody titer ->80% protection against O1 and O78 challenges | [205] |
| PapGII196 vaccine | Broilers (CH2) | I/M | -High IgG level -No protection against CH2 challenge | [192] |
| FimH (FimH156) vaccine | Broilers (CH2) | I/M and I/N | -High IgG and IgA level -No protection against CH2 challenge | [193] |
| APEC O78:EC99 cell surface antigens (FimA, PapG, IutA, and LPS) | Broilers (EC99) | I/N | -High IgA, IgG, and IgM levels -Less mortality | [197] |
| Liposomal inactivated APEC O78:KAI-2 vaccine | Layers (APEC O78:PDI386) | Eye drop or spray | -High IgG and IgA levels -Less mortality, APEC load, and lesions | [187] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathogens 2021, 10, 467. https://doi.org/10.3390/pathogens10040467
Kathayat D, Lokesh D, Ranjit S, Rajashekara G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathogens. 2021; 10(4):467. https://doi.org/10.3390/pathogens10040467
Chicago/Turabian StyleKathayat, Dipak, Dhanashree Lokesh, Sochina Ranjit, and Gireesh Rajashekara. 2021. "Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies" Pathogens 10, no. 4: 467. https://doi.org/10.3390/pathogens10040467