
A Novel Transcription Factor VPA0041 Was Identified to Regulate the Swarming Motility in Vibrio parahaemolyticus
1
Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou University, Yangzhou 225009, China
2
Jiangsu Key Laboratory of Zoonosis, Yangzhou University, Yangzhou 225009, China
3
Key Laboratory of Prevention and Control of Biological Hazard Factors (Animal Origin) for Agri-Food Safety and Quality, Ministry of Agriculture of China, Yangzhou University, Yangzhou 225009, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pathogens 2022, 11(4), 453; https://doi.org/10.3390/pathogens11040453
Submission received: 22 March 2022
/
Revised: 7 April 2022
/
Accepted: 8 April 2022
/
Published: 10 April 2022
Abstract
:Vibrio parahaemolyticus can change their usual lifestyle of surviving in an aqueous environment attached to a host, wherein both swimming motility and swarming motility play important roles in lifestyle changes, respectively. VPA0041 is a novel transcription factor involved in regulating the swarming ability of V. parahaemolyticus. The deletion of the vpa0041 gene resulted in the loss of swarming motility in the brain heart infusion (BHI) agars, while the swimming motility was unaffected by VPA0041. Transmission electron microscope (TEM) assays showed that no flagellum was found around the bacterial cells. RNA-sequencing (RNA-Seq) analysis revealed that VPA0041 regulated 315 genes; 207 genes were up-regulated, and 108 genes were down-regulated. RNA-seq results indicated that the lateral flagellar genes were down-regulated by VPA0041, which was confirmed by real-time quantitative reverse transcription-polymerase chain reaction (qRT-PCR). Electrophoretic mobility shift assays (EMSA) demonstrated that VPA0041 directly bound to the promoters of vpa0264, vpa1548, and vpa1550 to regulate the expression of the lateral flagellar genes. Our results demonstrated that the transcription factor VPA0041 could directly regulate the expression of lateral flagellar genes to mediate the swarming motility in V. parahaemolyticus.
1. Introduction
Vibrio parahaemolyticus is a gram-negative bacterium found in various aqueous environments, including marine and coastal environments [1,2,3,4]. V. parahaemolyticus expresses two flagellar systems responsible for swimming motility and swarming motility, respectively [5]. Individual swimming behavior allows the bacteria to swim in an aqueous environment, while swarming is the movement of bacteria over solid surfaces or during host colonization [6]. Changes of environmental conditions are closely related to the disease outbreaks caused by bacterial pathogens. V. parahaemolyticus is a foodborne pathogen that can be influenced by changes of environment factors [7]. Understanding the factors that could influence the dissemination of pathogens in the environment is essential for controlling opportunistic pathogens.
The marine and freshwater environment is a valuable source for bacteria worldwide. V. parahaemolyticus has an intricate life cycle that depends on environmental conditions, and contains swimmer and swarmer cells for living in liquid and on solid environments, respectively [7]. The single polar flagellum is encoded by genes responsible for the swimming motility in liquid environments. When V. parahaemolyticus attaches to solid surfaces, the lateral flagellum is stimulated and responsible for the swarming ability [8,9,10,11,12]. A previous study showed swarm colonies could be a continuous source of cells released into the environment. The released cells could be spread in the liquid environment and attach to new surfaces, but not stay within the liquid environment [7]. The flagellum contributes to bacterial pathogenicity by touching surfaces to adhere to either normal or degenerating tissue cells and promoting the biofilm formation to help the bacterial colonize to host [13,14,15]. Understanding the adaptation of swarming motility is essential for understanding the spread and pathogenicity of V. parahaemolyticus in the environment.
Motility has been identified as an essential virulence for the survival and colonization of V. parhaemolyticus [12,16]. The expression of flagellar genes is tightly regulated by the transcription regulators and sigma factors. The expression of polar flagellar genes is continuous, whereas the lateral flagellar genes are induced by surface growth, iron limitation, calcium presence, and inhibition of polar flagellum [17,18,19]. CalR, which is restrained by calcium acting as a LysR-type transcription factor in V. parahaemolyticus, negatively regulates laf genes, resulting in inhibiting the swarming motility [19]. The T3SS1 AraC-type regulator ExsA can inhibit LafK, the master regulator of the lateral flagellar system in the negative-feedback loop regulated by calR in high calcium environments [19]. The quorum sensing master regulator OpaR can inhibit the expression of laf gene to controlling the swarming motility [20]. Besides, the transcription factors VPA1701, SwrZ/SwrT, and the two-component system histidine kinase PhoR can down-regulate the expression of lateral flagellar genes for swarming motility in V. parahaemolyticus [20,21,22].
Two hundred and thirty genes have been identified that contribute to the colonization of V. parahaemolyticus to the small intestine by Tn-seq [23]. Among these 230 genes, 22 transcriptional factor genes have been selected to construct the deletion mutant strains, and investigate their impact to the swimming and swarming motility [21]. We found that VPA0041 was involved in the regulation of swarming motility. Thus, we further investigated the molecular mechanism of VPA0041 in regulating the swarming motility of V. parahaemolyticus. RNA-seq was used to identify the regulon of VPA0041 protein, and the lateral flagellar system was down-regulated in the ∆vpa0041 mutant strains, which was confirmed by qRT-PCR. Our findings revealed the regulation mechanism of VPA0041, which could determine the swarming motility in V. parahaemolyticus.
2. Results
2.1. VPA0041 Regulated the Swarming Motility of V. parahaemolyticus
In this study, we investigated the regulation of swarming motility by VPA0041 in V. parahaemolyticus. The vpa0041 deletion mutant strain did not show a significant difference in swimming motility, while the swarming motility was significantly decreased compared to WT on the BHI plated with 1.5% agar (Figure 1A). The complemented strains vpa0041+ restored the swarming ability, which confirmed the role of VPA0041 in swarming motility in V. parahaemolyticus (Figure 1A). The TEM was used to observe the flagellum of V. parahaemolyticus strains, and no flagellum was observed in the Δvpa0041 strain, whereas several typical lateral flagellums were observed surrounding WT and vpa0041+ cells (Figure 1B). These results indicated that the deletion of VPA0041 could down-regulate the lateral flagellum assembly and the ability of swarming motility.
2.2. The Annotation of VPA0041 Protein in V. parahaemolyticus
The study showed that VPA0041 was recognized as a new transcription regulator of the swarming motility in V. parahaemolyticus. The vpa0041 gene was 897 bp in length encoding 299 amino acids belonging to the LysR family, which contained a helix-turn-helix (HTH) domain at the N-terminal and a co-factor binding domain at C-terminal. BlastP results from GenBank indicated that V. parahaemolyticus VPA0041 shared 95%, 96%, 87%, 78%, 87%, and 77% identities to N646-3496 in Vibrio alginolyticus, VIBHAR_06999 in Vibrio campbellii, VIC_002269 in Vibrio coralliilyticus, AOT11_01700 in Vibrio vulnificus, VISP3789_21993 in Vibrio splendidus, and VCA1020 in Vibrio cholerae, respectively (Figure 2). This result indicated that the VPA0041 could be highly conserved with primarily the DNA binding domain among the Vibrio species.
2.3. The Global Transcriptional Analysis of VPA0041 in V. parahaemolyticus
VPA0041 protein was identified as a transcription factor engaged in regulating the swarming motility in V. parahaemolyticus. RNA-seq was used to identify the regulon of VPA0041 and further elucidate the potential mechanisms of VPA0041-regulated swarming motility. Comparison of the RNA-seq data for the WT and Δvpa0041 incubated on the BHI agar plates showed that 315 genes were significantly regulated by VPA0041 (log2FC ≤ −1 or log2FC ≥ 1; p < 0.05), 207 genes were up-regulated and 108 genes down-regulated in Δvpa0041 compared to WT (Figure 3A).
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed to identify the specific pathways regulated by VPA0041. Upwards of 100 pathways were activated, and several pathways are shown in Figure 3B. A total of 18 genes were assigned to the two-component system KEGG pathway (12 genes up-regulated and 6 genes down-regulated). Twelve genes associated with the bacterial secretion system were regulated by VPA0041 (six genes up-regulated and six genes down-regulated), including the type III secretion system 1 and type VI secretion system 2. The metabolic pathway (64 genes up-regulated and 19 genes down-regulated) and the ABC transporters pathway (16 genes up-regulated and 10 genes down-regulated) are also identified in the regulon of VPA0041. Notably, 15 genes of the lateral flagellar system and three genes of bacterial chemotaxis were down-regulated in the Δvpa0041 compared with the WT, indicating that VPA0041 protein might act as a positive regulator for the swarming motility in V. parahaemolyticus. The transcript levels of polar flagellar genes were not influenced by VPA0041 protein. These results demonstrated that VPA0041 maybe play an essential role in the multiple metabolic and virulence pathways in V. parahaemolyticus.
2.4. VPA0041 Regulates the Expression of the Lateral Flagellar Genes
RNA-seq analysis indicated that the lateral flagellar genes were down-regulated in the regulon of VPA0041. Among the 38 lateral flagellar system genes, 15 genes were significantly decreased in the Δvpa0041 compared to the WT (Figure 4A). Furthermore, qRT-PCR showed that the expression of vpa0264, vpa1537, vpa1538, vpa1539, vpa1540, and vpa1548 were significantly down-regulated in Δvpa0041 compared with the WT; and the complemented strains vpa0041+ restored the transcript level of these genes to the WT (Figure 4B). These results indicated that VPA0041 could regulate the expression of lateral flagellar genes to mediate the swarming motility in V. parahaemolyticus.
2.5. VPA0041 Directly Bound to the Promoters of vpa0264, vpa1548, and vpa1550 to Activate the Lateral Flagellar System
Thirty-eight lateral flagellar system genes were divided into 2 clusters located in chromosome II of V. parahaemolyticus [21]. Bioinformation analysis showed that the lateral flagellar system might contain nine promoters, shown in Figure 4A. The vpa0264-vpa0274 cluster, vpa1548 gene, and vpa1550-vpa1557 cluster were significantly down-regulated in the Δvpa0041 strain. EMSA results showed that the VPA0041 protein was directly bound to the promoter regions of vpa0264, vpa1548, and vpa1550 in a concentration-dependent manner (Figure 5A–C). The negative probe remained unbound with the highest concentration of VPA0041 protein (Figure 5D).
The promoter region of vpa0264 was shown in the Figure 6A. Four different probes were used to analyze binding sites of VPA0041 in the promoter of vpa0264. The EMSA results indicated that VPA0041 could bind to the vpa0264 promoter, promoter-1 and promoter-2 but not promoter-3 (Figure 5A and Figure 6B). Then the MEME-Suit tool (http://meme-suite.org; accessed on 13 August 2021) was used to identify the binding motif of the VPA0041 protein based on the sequences of vpa0264, vpa1548, and vpa1550 promoters. As showed in the Figure 6C, the specific binding motif of the VPA0041 is a TA-rich region, and the VPA0041-binding site in the vpa0264 promoter was 5′-AAGTCATTGATAAATATAAA-3′ (Figure 6A). In addition, the predicted VPA0041-binding sites in the promoters of vpa1548 and vpa1550 were showed in the Figure S1A. Then we also used the EMSA to verify the binding site of VPA0041 in the promoter of vpa1548 and vpa1550. However, the VPA0041 protein still could bind to all of the different probes of vpa1548 or vpa1550 promoter (Figure S1B–E). Our current results indicated that VPA0041 protein could directly bind to the promoters of vpa0264, vpa1548, and vpa1550 to induce the expression of the lateral flagellar system genes and responsible for the swarming motility of V. parahaemolyticus.
3. Discussion
The expression of lateral flagellar genes has three tiers (classes I, II, and III) and is tightly regulated by LafK dependent on σ54 and σ28 in Vibrio parahaemolyticus and other Vibrios [19,24]. Apart from the regulation of LafK to the lateral flagellar genes, the other transcriptional factors independent of σ54 and σ28 have also been identified to control the swarming motility behavior of Vibrio parahaemolyticus, these transcriptional factors including VPA1701, SwrT/SwrZ, OpaR, ScrG/ScrC, ToxR, OxyR, CalR, and HU [19,20,21,25,26,27,28,29]. In this study, we identified a novel transcriptional factor, VPA0041, which regulated the swarming motility, and 15 flagellar assembly genes were down-regulated by VPA0041. EMSA confirmed that the VPA0041 protein could directly bind to the promoters region of vpa0264-vpa0272 cluster, vpa1548 gene, and vpa1550-vpa1557 cluster (Figure 5). The vpa0264-vpa0272 cluster could encode the FlgBCDEFGHIJ protein, which could form the rod, P/L ring, and hook of the flagellum [9]. The vpa1548 gene could encode the FliC, which could form the filament of the flagellum [30]. The vpa1550 and vpa1551 genes could encode the FliDS protein, forming the filament cap of the flagellum [9,30]. Notably, the swimming motility was not affected in Δvpa0041, but no polar flagellum was observed in Δvpa0041. This result was due to that the bacterial were cultured in BHI agar and collected to observe the lateral flagellum, while the polar flagellum could not be induced in this condition. These results demonstrated that the VPA0041 could down-regulated the expression of flgBCDEFGHIJ, fliC, and fliDS to block the generation of the lateral flagellum.
VPA0041 is a transcription factor belonging to the LysR family, which is an abundant prokaryote and contains a conserved HTH motif at the N-terminal and a co-factor binding domain at the C-terminal [31]. LysR-type transcriptional regulators are distributed among diverse bacteria, regulating various biological processes, such as virulence, metabolism, quorum sensing, and motility [32,33,34]. In this study, we identified a novel LysR family transcription factor, VPA0041, capable of regulating the swarming motility of V. parahaemolyticus (Figure 1). RNA-seq results indicated that the VPA0041 protein could regulate the expression of lateral flagellar system genes, and confirmed by qRT-PCR (Figure 4). Moreover, the expression levels of lateral flagellar genes in the complementary strain were higher than those of the WT (Figure 4B), but the swarming motility of the complementary strain was weaker than that of WT (Figure 1A). This result may be due to that the high expression of VPA0041 protein increased the transcript levels of the lateral flagellar genes in complementary strain. However, the mRNA of the lateral flagellar genes translated to protein may be limited in the bacterial and fewer proteins were expressed, leading to the swarming motility of complementary strain was weaker than that in WT. In addition, the VPA0041 protein could directly bind to the promoter region of the lateral flagellar system to regulate the expression of the lateral flagellar genes and mediate the swarming motility in V. parahaemolyticus (Figure 5).
Results showed that the LysR family transcription factor VPA0041 could down-regulate the swarming motility in V. parahaemolyticus. Previous studies showed that LysR-type protein could act as a transcription regulator regulating the expression of motility in various bacteria. In E. coli, the LysR-type transcription regulators LrhA and HdfR could actively regulate the expression of FlhDC, which was the master regulator of the flagellar system and responsible for motility [35,36]. RovM was identified to be homologous to LrhA and regulated the cell invasion, virulence, and motility in Yersinia pseudotuberculosis [37]. Conversely, HexA was identified as a LysR-type transcription factor, which could inhibit the expression of fliA and fliC, responsible for motility ability [38]. The LysR-type transcription factor activates or inhibits the expression of the flagellar system genes. In our study, the LysR family protein VPA0041 could directly bind the promoter region of vpa0264, vpa1548, and vpa1550; these genes were involved in the lateral flagellar system in V. parahaemolyticus. Besides, our results identified the specific binding site of VPA0041 protein was 5′-AAGTCATTGATAAATATAAA-3′ in the promoter vpa0264, which was a TA-rich region (Figure 6). The EMSA results shown that VPA0041 protein could bind to all DNA probes of vpa1548 or vpa1550 promoter (Figure S1D,E), indicating that more than one binding sites were existence in these promoters. There were multiple bands in the vpa1550 promoter-1 to -3, while only one band was found in the vpa1550 promoter-4 (Figure S1E). These results indicated that one binding site might located in the promoter-4 region which was consistent with binding site predicted by MEME, and another binding site may existence in the region of 50–100 bp relative to ATG. Another LysR family protein GcdR also shown multiple bands and identified two specific binding sites in the promoter, one of them was a TA-rich region [39].
Concerning the pathogenesis of V. parahaemolyticus, a wide variety of virulence factors, including thermostable direct hemolysin (TDH), TDH related hemolysin (TRH), adhesins, motility, biofilm, Type III secretion system, and Type VI secretion system, have been comprehensively studied [6,40,41,42,43]. Previously studies have demonstrated the relationship between the motility and pathogenicity in several bacterial species including V. parahaemolyticus, Vibrio alginolyticus, Acinetobacter baumannii, and Lysobacter [21,44,45,46,47,48,49]. Interestingly, our RNA-seq results showed that VPA0041 significantly regulated 315 genes on the BHI agar plate (Figure 3). The KEGG pathway analysis also indicated that several virulence-associated pathways were enhanced, including the T3SS1, T6SS1, and lateral flagellar system. Previous studies reported that the lateral flagellum and killing function of T6SS was induced on the agar plate [42,50], which was consistent with our RNA-seq results. T3SS1 is related to motility, biofilm formation, cytotoxicity and contributes to the survival of V. parahaemolyticus in the environment [51,52]. Additionally, these virulence-associated genes were also regulated by VPA0041, indicating that the VPA0041 might be a virulence regulator in V. parahaemolyticus (Figure 3). The molecular regulation mechanisms of VPA0041 to T3SS1 and T6SS1 need to be further elucidated.
In summary, this study demonstrated that VPA0041 was required for swarming motility and might be beneficial for the survival and pathogenesis of V. parahaemolyticus in the surface or tissue environment. This study provides evidence of the participation of the VPA0041 in regulation of the swarming motility of V. parahaemolyticus.
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Growth Conditions
All bacterial strains and plasmids were listed in Table 1, and the primers used in this study were listed in Table 2. V. parahaemolyticus RIMD2210633 WT and its derivatives, as well as E. coli, were incubated in Luria-Bertani (LB) at 37 °C while shaking at 220 rpm. When necessary, the carbenicillin (Carb, 50 μg/mL), chloramphenicol (Cm, 25 μg/mL), kanamycin (Km, 50 μg/mL), and isopropyl β-D-1-thiogalactopyranoside (IPTG, 1 mM) were added to the medium.
4.2. Construction of vpa0041 Mutant and Complemented Strains
The construction of vpa0041 deletion mutant and complemented strains were completed as previously described [57]. The primers for the construction of the vpa0041 mutant strain were list in Table 2. The PCR-amplified DNA products of up-stream (627 bp) and down-stream (651 bp) were generating to the overlap fragments (1278 bp) by overlap PCR. The overlapped DNA products were inserted into the pDM4 plasmid and transformed into E. coli SM10 λpir. Subsequently, the recombinant plasmid Δvpa0041::pDM4 was transformed into the WT by conjugation and selected in the LB agar with Carb and Cm. Finally, the second cross-over recombination was selected on the LB agar containing 10% sucrose. The mutant strain (∆vpa0041) was verified by PCR (vpa0041-out/in-F/R) and sequencing.
The ribosome binding site (RBS) and open reading frame (ORF) regions of the vpa0041 gene were amplified by PCR with specific primers (vpa0041-F/R) and cloned into an expression vector pMMB207. The recombinant plasmid (vpa0041::pMMB207) was then transformed into E. coli SM10λpir and conjugated into the vpa0041 mutant strain; PCR confirmed this with specific primers (pMMB207-F/R) and sequencing. The expression of VPA0041 was induced by adding 0.1 mM IPTG.
4.3. The Motility Analysis
WT, ∆vpa0041 and vpa0041+ were grown in LB medium for 12 h and diluted into new fresh LB medium until OD600 of 1.0. Bacterial suspensions measuring 2 μL were dropped onto LB plates with 0.3% agar at 37 °C for 12 h, and Brain Heart Infusion (BHI) plates with 1.5% agar at 30 °C for 24 h to observe the swimming motility and swarming motility, respectively. Three independent experiments were carried out for the motility assay.
4.4. The Transmission Electron Microscope Analysis
The WT ∆vpa0041 and vpa0041+ strains were incubated on swarming plates for 24 h. The bacteria were then gently washed by 0.01 M PBS into 2 mL Eppendorf tubes. Subsequently, the suspension was added dropwise onto grids for 5 min until the mesh was dry, then the dried samples were covered with 5% uranyl acetate for 30 min in the dry environment before observation by transmission electron microscopy (JEM 2100, Tokyo, Japan).
4.5. qRT-PCR Analysis of the Lateral Flagellar Genes
V. parahaemolyticus WT ∆vpa0041 and vpa0041+ strains were cultured on BHI plates for 18 h, and cells were scraped and harvested from plates, then the total RNA was extracted by the Bacterial Total RNA Extraction Kit (Shangon Biotech, Shanghai, China). The RNA was then treated with RNase free DNase I (Takara, Tsuruga, Japan) to remove gDNA, and the PrimeScript TM RT reagent Kit (Takara, Tsuruga, Japan) was used to generated cDNA. qRT-PCR was performed with SYRB ® Premix Ex Taq TM (Takara, Tsuruga, Japan) on the ABI StepOnePlus Real-Time PCR System. The reverses transcription reaction mix was performed with 20 μL volume including 10 μL SYBR Premix Ex Taq II, 2 μL cDNA, 1 μL each primer, 6 μL ddH2O. Six lateral flagellar genes of each sample were checked by 2−∆∆Ct, and the gyrB gene was used as a control. The primers were listed in Table 2.
4.6. RNA-Seq Analysis
V. parahaemolyticus WT and Δvpa0041 strains were cultured on BHI agar plates, and the total RNA was extracted by the Bacterial Total RNA Extraction Kit (Shangon Biotech, Shanghai, China). RNA samples were digested using DNase I (Promega, Madison, WI, USA) to remove gDNA. Three parallel RNA of each strain were sequenced by the Illumina HiSeq 2000 platform (GENEWIZ, Suzhou, China). The subsequent procedures and statistical analysis were carried out as previously described [22]. In brief, the clean data were aligned to the reference genome of V. parahaemolyticus RIMD2210633 using Rockhopper [58,59]. The significant difference expressed genes were defined as the value of log2 foldchange ≥ 1 or ≤ −1, and p < 0.05. The differentially expressed genes were annotated to the KEGG pathway database [60]. Raw sequencing reads were deposited in the European Nucleotide Archive database under accession number PRJEB39385.
4.7. Purification of VPA0041 Protein
The ORF of vpa0041 gene was amplified with primers vpa0041-his-F/R and cloned into the pET30a with BamHI and XhoI. The recombined plasmid vpa0041::pET30a was transformed into E. coli BL21 (DE3), and verified by PCR with the primers pET30a-F/R. The vpa0041::pET30a/BL21 strain was cultured in LB and the expression of VPA0041 protein was induced by IPTG. The bacteria were cultured in 16 °C with 120 rpm for 12 h, before bacterial pellets were collected, washed twice with the binding buffer (0.5 M NaCl, 20 mM Tris-HCl, 5 mM imidazole, pH 7.9) and lysed by sonicate. VPA0041 protein was purified by His Bind Purification Kit (Novagen, Darmstadt, Germany). 12% SDS-PAGE was used to verify the purity of VPA0041 protein (Figure S2).
4.8. Electrophoretic Mobility Shift Assay
The EMSA analysis was performed as previously described [61]. Briefly, the promoter regions of the lateral flagellar genes (vpa0264, vpa1548, vpa1550, and the negative DNA) were amplified with 5′-FAM primers (Table 2). The reaction mix was implemented in 20 μL containing 10 ng DNA probes, 1 μg poly(dI:dC), 4 μL 5 × binding buffer (10 mM NaCl, 0.1 mM DTT, 0.1 mM EDTA, 10 mM Tris, pH 7.4) and an increasing amount of VPA0041-his protein. After incubation at 25 °C for 30 min, the reaction mixes were separated by 6% native PAGE gels for 2 h at 100 V. The gel was scanned by Typhoon FLA 9500 (GE Healthcare, Uppsala, Sweden).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens11040453/s1. Figure S1: Identify the specific binding sites of VPA0041 protein. (A) The promoter regions of the VPA0041 directly regulated genes, vpa0264, vpa1548, and vpa1550. The conserved VPA0041-binding site is indicated by the blue underline. The translation start site is labeled with the red. (B,C) The promoter regions of vpa1548 (B) and vpa1550 (C). (D,E) EMSA analysis of VPA0041 binding sites with the various truncations; Figure S2: SDS-PAGE of VPA0041 protein. 12% acrylamide gel was used to verify the purity of VPA0041 protein. M represent for marker, 1 represent for VPA0041 protein.
Author Contributions
Conceptualization, D.G. and H.M.; methodology, M.L. and H.M.; software, M.L. and Y.L.; validation, H.M. and Y.L.; formal analysis, M.L. and H.M.; investigation, M.L. and H.M.; resources, D.G.; data curation, D.G.; writing—original draft preparation, H.M.; writing—review and editing, M.L. and D.G.; visualization, M.L. and H.M.; supervision, D.G.; project administration, D.G.; funding acquisition, D.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, 31700122 and China Postdoctoral Science Foundation, 2018M632388.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw sequencing reads of RNA-seq were deposited in the European Nucleotide Archive database under accession number PRJEB39385.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Austin, B. Vibrios as causal agents of zoonoses. Vet. Microbiol. 2010, 140, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.N.; Bowers, J.C.; Griffitt, K.J.; Molina, V.; Clostio, R.W.; Pei, S. Ecology of Vibrio parahaemolyticus and Vibrio vulnificus in the coastal and estuarine waters of Louisiana, Maryland, Mississippi, and Washington (United States). Appl. Environ. Microbiol. 2012, 78, 7249–7257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Zhang, X.; Fan, H.; Li, Y.; Hu, Q.; Yang, R.; Cui, Y. Genetic diversity, virulence factors and X farm-to-table spread pattern of Vibrio parahaemolyticus food-associated isolates. Food Microbiol. 2019, 84, 103270. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Dong, S.; Yan, Y.; Zhan, L.; Zhang, J.; Chen, J.; Zhang, Z.; Zhang, Y.; Mei, L. Prevalence and Population Analysis of Vibrio parahaemolyticus Isolated from Freshwater Fish in Zhejiang Province, China. Foodborne Pathog. Dis. 2021, 18, 139–146. [Google Scholar] [CrossRef]
- Sar, N.; MrCarter, L.; Simon, M.; Silverman, M. Chemotactic control of the two flagellar systems of Vibrio parahaemolyticus. J. Bacteriol. 1990, 172, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Harshey, R.M. Bacterial motility on a surface: Many ways to a common goal. Annu. Rev. Microbiol. 2003, 57, 249–273. [Google Scholar] [CrossRef]
- Freitas, C.; Glatter, T.; Ringgaard, S. The release of a distinct cell type from swarm colonies facilitates dissemination of Vibrio parahaemolyticus in the environment. ISME J. 2020, 14, 230–244. [Google Scholar] [CrossRef]
- McCarter, L.; Silverman, M. Surface-induced swarmer cell differentiation of Vibrio parahaemolyticus. Mol. Microbiol. 1990, 4, 1057–1062. [Google Scholar] [CrossRef]
- McCarter, L.L. Dual flagellar systems enable motility under different circumstances. J. Mol. Microbiol. Biotechnol. 2004, 7, 18–29. [Google Scholar] [CrossRef]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of Vibrio cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- McCarter, L.L. Bacterial acrobatics on a surface: Swirling packs, collisions, and reversals during swarming. J. Bacteriol. 2010, 192, 3246–3248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gode-Potratz, C.J.; Kustusch, R.J.; Breheny, P.J.; Weiss, D.S.; McCarter, L.L. Surface sensing in Vibrio parahaemolyticus triggers a programme of gene expression that promotes colonization and virulence. Mol. Microbiol. 2011, 79, 240–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merino, S.; Shaw, J.G.; Tomás, J.M. Bacterial lateral flagella: An inducible flagella system. FEMS. Microbiol. Lett. 2016, 263, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [Green Version]
- Murray, T.S.; Ledizet, M.; Kazmierczak, B. Swarming motility, secretion of type 3 effectors and biofilm formation phenotypes exhibited within a large cohort of Pseudomonas aeruginosa clinical isolates. J. Med. Microbiol. 2010, 59, 511–520. [Google Scholar] [CrossRef]
- Khan, F.; Tabassum, N.; Anand, R.; Kim, Y.M. Motility of Vibrio spp.: Regulation and controlling strategies. Appl. Microbiol. Biotechnol. 2020, 104, 8187–8208. [Google Scholar] [CrossRef]
- McCarter, L.; Silverman, M. Iron regulation of swarmer cell differentiation of Vibrio parahaemolyticus. J. Bacteriol. 1989, 171, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Kawagishi, I.; Imagawa, M.; Imae, Y.; McCarter, L.L.; Homma, M. The sodium-driven polar flagellar motor of marine Vibrio as the mechanosensor that regulates lateral flagellar expression. Mol. Microbiol. 1996, 20, 693–699. [Google Scholar] [CrossRef]
- Gode-Potratz, C.J.; Chodur, M.D.; McCarter, L.L. Calcium and iron regulate swarming and type III secretion in Vibrio parahaemolyticus. J. Bacteriol. 2010, 192, 6025–6038. [Google Scholar] [CrossRef] [Green Version]
- Jaques, S.; McCarter, L.L. Three new regulators of swarming in Vibrio parahaemolyticus. J. Bacteriol. 2006, 188, 2625–2635. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.; Meng, H.; Li, Y.; Ge, H.; Jiao, X. A GntR family transcription factor (VPA1701) for swarming motility and colonization of Vibrio parahaemolyticus. Pathogens 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, H.; Gu, D.; Lu, X.; Zhou, X.; Xia, X. Transcriptomic analysis of PhoR reveals its role in regulation of swarming motility and T3SS expression in Vibrio parahaemolyticus. Microbiol. Res. 2020, 235, 126448. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, T.P.; Chao, M.C.; Abel, S.; Blondel, C.J.; Abel Zur Wiesch, P.; Zhou, X.; Davis, B.M.; Waldor, M.K. Genetic analysis of Vibrio parahaemolyticus intestinal colonization. Proc. Natl. Acad. Sci. USA 2016, 113, 6283–6288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Zhang, S.; Ren, W.; Gong, X.; Long, H.; Zhang, X.; Cai, X.; Huang, A.; Xie, Z. Roles of rpoN in biofilm formation of Vibrio alginolyticus HN08155 at different cell densities. Microbiol. Res. 2021, 247, 126728. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; McCarter, L.L. ScrG, a GGDEF-EAL protein, participates in regulating swarming and sticking in Vibrio parahaemolyticus. J. Bacteriol. 2007, 189, 4094–4107. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, R.B.; Antunes, L.C.; Greenberg, E.P.; McCarter, L.L. Vibrio parahaemolyticus ScrC modulates cyclic dimeric GMP regulation of gene expression relevant to growth on surfaces. J. Bacteriol. 2008, 190, 851–860. [Google Scholar] [CrossRef] [Green Version]
- Phan, N.Q.; Uebanso, T.; Shimohata, T.; Nakahashi, M.; Mawatari, K.; Takahashi, A. DNA-binding protein HU coordinates pathogenicity in Vibrio parahaemolyticus. J. Bacteriol. 2015, 197, 2958–2964. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.; Fen, S.; Yu, S.; Wong, H. Influence of oxyR on growth, biofilm formation, and mobility of Vibrio parahaemolyticus. Appl. Environ. Microbiol. 2016, 82, 788–796. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Qiu, Y.; Tang, H.; Hu, L.; Yang, W.; Zhu, X.; Huang, X.; Wang, T.; Zhang, Y. ToxR is required for biofilm formation and motility of Vibrio parahaemolyticus. Biomed. Environ. Sci. 2018, 31, 848–850. [Google Scholar]
- Majander, F.; Korhonen, T.K.; Westerlund-Wikström, B. Simultaneous display of multiple foreign peptides in the FliD capping and FliC filament proteins of the Escherichia coli flagellum. Appl. Environ. Microbiol. 2005, 71, 4263–4268. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, S.E.; Oyston, P.C.F. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.M.; Ikawa, Y.; Tsuge, S. GamR, the LysR-type galactose metabolism regulator, regulates hrp gene expression via transcriptional activation of two key hrp regulators, HrpG and HrpX, in Xanthomonas oryzae pv. oryzae. Appl. Environ. Microbiol. 2016, 82, 3947–3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Wang, X.; Mao, Q.; Xu, R.; Zhou, X.; Ma, Y.; Liu, Q.; Zhang, Y.; Wang, Q. VqsA, a novel LysR-type transcriptional regulator, coordinates quorum sensing (QS) and is controlled by QS to regulate virulence in the pathogen Vibrio alginolyticus. Appl. Environ. Microbiol. 2018, 84, e00444-18. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, B.; Yang, P.; Wang, T.; Chang, Z.; Wang, J.; Wang, Q.; Li, W.; Wu, J.; Huang, D.; et al. LysR-type transcriptional regulator OvrB encoded in O island 9 drives enterohemorrhagic Escherichia coli O157:H7 virulence. Virulence 2019, 10, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, M.; Park, C. H-NS-Dependent regulation of flagellar synthesis is mediated by a LysR family protein. J. Bacteriol. 2000, 182, 4670–4672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehnen, D.; Blumer, C.; Polen, T.; Wackwitz, B.; Wendisch, V.F.; Unden, G. LrhA as a new transcriptional key regulator of flagella, motility and chemotaxis genes in Escherichia coli. Mol. Microbiol. 2002, 45, 521–532. [Google Scholar] [CrossRef]
- Heroven, A.K.; Dersch, P. RovM, a novel LysR-type regulator of the virulence activator gene rovA, controls cell invasion, virulence and motility of Yersinia pseudotuberculosis. Mol. Microbiol. 2006, 62, 1469–1483. [Google Scholar] [CrossRef]
- Harris, S.J.; Shih, Y.L.; Bentley, S.D.; Salmond, G.P. The hexA gene of Erwinia carotovora encodes a LysR homologue and regulates motility and the expression of multiple virulence determinants. Mol. Microbiol. 1998, 28, 705–717. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Kang, Z.; Guo, X.; Guo, S.; Xiao, D.; Liu, Y.; Ma, C.; Gao, C.; Xu, P. Regulation of glutarate catapolism by GntR family regulator CsiR and LysR family regulator GcdR in Pseudomonas putida KT2440. mBio 2019, 10, e01570-19. [Google Scholar] [CrossRef] [Green Version]
- Erwin, D.P.; Nydam, D.S.; Call, D.R. Vibrio parahaemolyticus ExsE is requisite for initial adhesion and subsequent type III secretion system1-dependent autophagy in HeLa cells. Microbiology 2012, 158, 2303–2314. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Zhang, Y. Structure, function and regulation of the thermostable direct hemolysin (TDH) in pandemic Vibrio parahaemolyticus. Microb. Pathog. 2018, 123, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yaakov, R.; Salomon, D. The regulatory network of Vibrio parahaemolyticus type VI secretion system 1. Environ. Microbiol. 2019, 21, 2248–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, S.; Okada, R.; Tandhavanant, S.; Hiyoshi, H.; Gotoh, K.; Iida, T.; Kodama, T. Export of a Vibrio parahaemolyticus toxin by the Sec and type III secretion machineries in tandem. Nat. Microbiol. 2019, 4, 781–788. [Google Scholar] [CrossRef]
- Liu, M.; Zhu, X.; Zhang, C.; Zhao, Z. LuxQ-LuxU-LuxO pathway regulates biofilm formation by Vibrio parahaemolyticus. Microbiol. Res. 2021, 250, 126791. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Tang, H.; Qiu, Y.; Yang, W.; Yang, H.; Zhou, D.; Huang, X.; Hu, L.; Zhang, Y. Quorum sensing regulates the transcription of lateral flagellar genes in Vibrio parahaemolyticus. Future Microbiol. 2019, 14, 1043–1053. [Google Scholar] [CrossRef]
- Yin, W.; Zhang, N.; Xu, H.; Gong, X.; Long, H.; Ren, W.; Zhang, X.; Cai, X.; Huang, A.; Xie, Z. Stress adaptation and virulence in Vibrio alginolyticus is mediated by two (p)ppGpp synthetase genes, relA and spoT. Microbiol. Res. 2021, 253, 126883. [Google Scholar] [CrossRef]
- Mea, H.J.; Yong, P.V.C.; Wong, E.H. An overview of Acinetobacter baumannii pathogenesis: Motility, adherence and biofilm formation. Microbiol. Res. 2021, 247, 126722. [Google Scholar] [CrossRef]
- Gautam, L.K.; Sharma, P.; Capalash, N. Attenuation of Acinetobacter baumannii virulence by inhibition of polyphosphate kinase 1 with repurposed drugs. Microbiol. Res. 2021, 242, 126627. [Google Scholar] [CrossRef]
- Brescia, F.; Marchetti-Deschmann, M.; Musetti, R.; Perazzolli, M.; Pertot, I.; Puopolo, G. The rhizosphere signature on the cell motility, biofilm formation and secondary metabolite production of a plant-associated Lysobacter strain. Microbiol. Res. 2020, 234, 126424. [Google Scholar] [CrossRef]
- Salomon, D.; Gonzalez, H.; Updegraff, B.L.; Orth, K. Vibrio parahaemolyticus type VI secretion system 1 is activated in marine conditions to target bacteria, and is differentially regulated from system 2. PLoS ONE 2013, 8, e61086. [Google Scholar] [CrossRef]
- Calder, T.; de Souza Santos, M.; Attah, V.; Klimko, J.; Fernandez, J.; Salomon, D.; Krachler, A.M.; Orth, K. Structural and regulatory mutations in Vibrio parahaemolyticus type III secretion system display variable effects in virulence. FEMS Microbiol. Lett. 2014, 361, 107–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, I.T.; Tulman, E.R.; Geary, S.J.; Zhou, X. A gatekeeper protein contributes to T3SS2 function via interaction with an ATPase in Vibrio parahaemolyticus. Microbiol. Res. 2021, 252, 126857. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Millet, Y.A.; Chao, M.C.; Sasabe, J.; Davis, B.M.; Waldor, M.K. A genome-wide screen reveals that the Vibrio cholerae phosphoenolpyruvate phosphotransferase system modulates virulence gene expression. Infect. Immun. 2015, 83, 3381–3395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Konkel, M.E.; Call, D.R. Vp1659 is a Vibrio parahaemolyticus type III secretion system 1 protein that contributes to translocation of effector proteins needed to induce cytolysis, autophagy, and disruption of actin structure in HeLa cells. J. Bacteriol. 2010, 192, 3491–3502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Wang, S.; Yu, F.; Zhang, L.; Qi, G.; Liu, Y.; Gao, S.; Kan, B. Construction and evaluation of a safe, live, oral Vibrio cholerae vaccine candidate, IEM108. Infect. Immun. 2003, 71, 5498–5504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, V.M.; Bäckman, A.; Bagdasarian, M. A series of wide-host range low-copy-number vectors that allow direct screening for recombinants. Gene 1991, 97, 39–47. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lauritz, J.; Jass, J.; Milton, D.L. A ToxR homolog from Vibrio anguillarum serotype O1 regulates its own production, bile resistance, and biofilm formation. J. Bacteriol. 2002, 184, 1630–1639. [Google Scholar] [CrossRef] [Green Version]
- Tjaden, B. De novo assembly of bacterial transcriptomes from RNA-seq data. Genome Biol. 2015, 16, 1. [Google Scholar] [CrossRef] [Green Version]
- McClure, R.; Balasubramanian, D.; Sun, Y.; Bobrovskyy, M.; Sumby, P.; Genco, C.A.; Vanderpool, C.K.; Tjaden, B. Computational analysis of bacterial RNA-seq data. Nucleic Acids Res. 2013, 41, e140. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Gu, D.; Guo, M.; Yang, M.; Zhang, Y.; Zhou, X.; Wang, Q. A σE-mediated temperature gauge controls a switch from LuxR-mediated virulence gene expression to thermal stress adaptation in Vibrio alginolyticus. PLoS Pathog. 2016, 12, e1005645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
VPA0041 regulates the swarming motility of V. parahaemolyticus. (A) The Vibrio parahaemolyticus WT, ∆vpa0041 and vpa0041+ were incubated in swimming plates (LB + 0.3% Agar) at 37 °C and swarming plates (BHI + 1.5% Agar) at 30 °C. (B) TEM micrographs show the generation of lateral flagellum in V. parahaemolyticus WT, ∆vpa0041, and vpa0041+.
Figure 1.
VPA0041 regulates the swarming motility of V. parahaemolyticus. (A) The Vibrio parahaemolyticus WT, ∆vpa0041 and vpa0041+ were incubated in swimming plates (LB + 0.3% Agar) at 37 °C and swarming plates (BHI + 1.5% Agar) at 30 °C. (B) TEM micrographs show the generation of lateral flagellum in V. parahaemolyticus WT, ∆vpa0041, and vpa0041+.

Figure 2.
Multiple amino acid sequence alignments of VPA0041 proteins in Vibrios. Red rectangular frames indicate the homology of VPA0041 protein between V. parahaemolyticus and other Vibrios. The graphs show differences, and dots indicate missing residues.
Figure 2.
Multiple amino acid sequence alignments of VPA0041 proteins in Vibrios. Red rectangular frames indicate the homology of VPA0041 protein between V. parahaemolyticus and other Vibrios. The graphs show differences, and dots indicate missing residues.
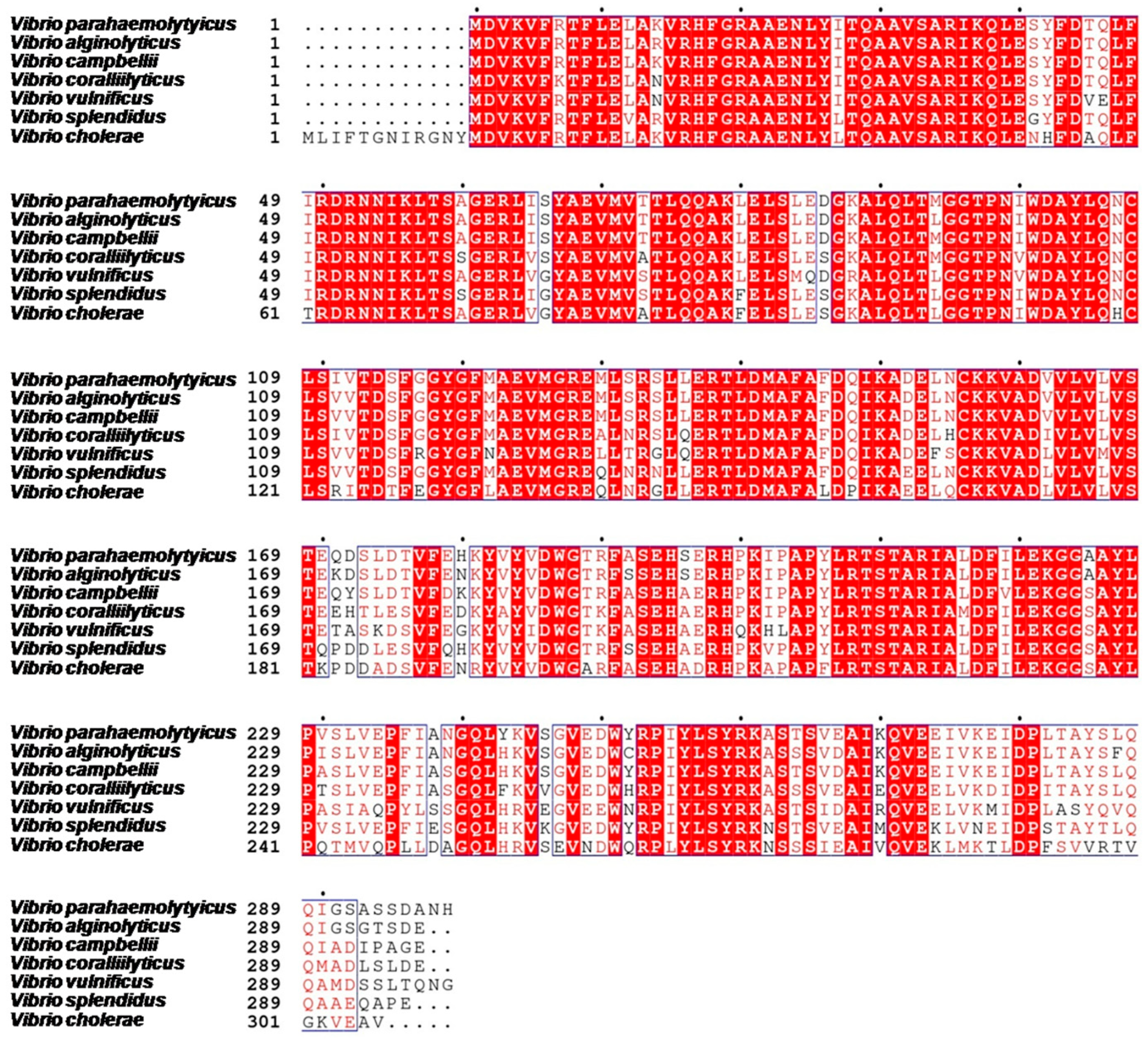
Figure 3.
RNA-seq reveals the genes affected by vpa0041. (A) The pie chart shows the number of genes affected by vpa0041 according to ∆vpa0041 compared to WT cultured in BHI agar plates. (B) KEGG analysis reveals the different pathways associated with vpa0041 in ∆vpa0041 compared to WT. The orange shows the up-regulated genes; the green shows the down-regulated genes.
Figure 3.
RNA-seq reveals the genes affected by vpa0041. (A) The pie chart shows the number of genes affected by vpa0041 according to ∆vpa0041 compared to WT cultured in BHI agar plates. (B) KEGG analysis reveals the different pathways associated with vpa0041 in ∆vpa0041 compared to WT. The orange shows the up-regulated genes; the green shows the down-regulated genes.

Figure 4.
VPA0041 regulates the expression of lateral flagellar genes. (A) RNA-seq performed the expression pattern of all lateral flagellar genes. The green columns indicate the significant down-regulated genes in Δvpa0041 compared to WT strain (fold change (Δvpa0041/WT) ≤ 0.5 and p < 0.05), the blue columns indicate the gene not influenced by VPA0041. The gray arrows indicate the configuration of the lateral flagellar genes on the chromosome, and the red arrows indicate the predicted promoters. (B) The expression pattern of several lateral flagellar genes in WT, ∆vpa0041, and vpa0041+ was examined by qRT-PCR. Data showed the mean ± SE from three independent experiments. ** p < 0.01; * p < 0.05, calculated by Student’s t-test.
Figure 4.
VPA0041 regulates the expression of lateral flagellar genes. (A) RNA-seq performed the expression pattern of all lateral flagellar genes. The green columns indicate the significant down-regulated genes in Δvpa0041 compared to WT strain (fold change (Δvpa0041/WT) ≤ 0.5 and p < 0.05), the blue columns indicate the gene not influenced by VPA0041. The gray arrows indicate the configuration of the lateral flagellar genes on the chromosome, and the red arrows indicate the predicted promoters. (B) The expression pattern of several lateral flagellar genes in WT, ∆vpa0041, and vpa0041+ was examined by qRT-PCR. Data showed the mean ± SE from three independent experiments. ** p < 0.01; * p < 0.05, calculated by Student’s t-test.
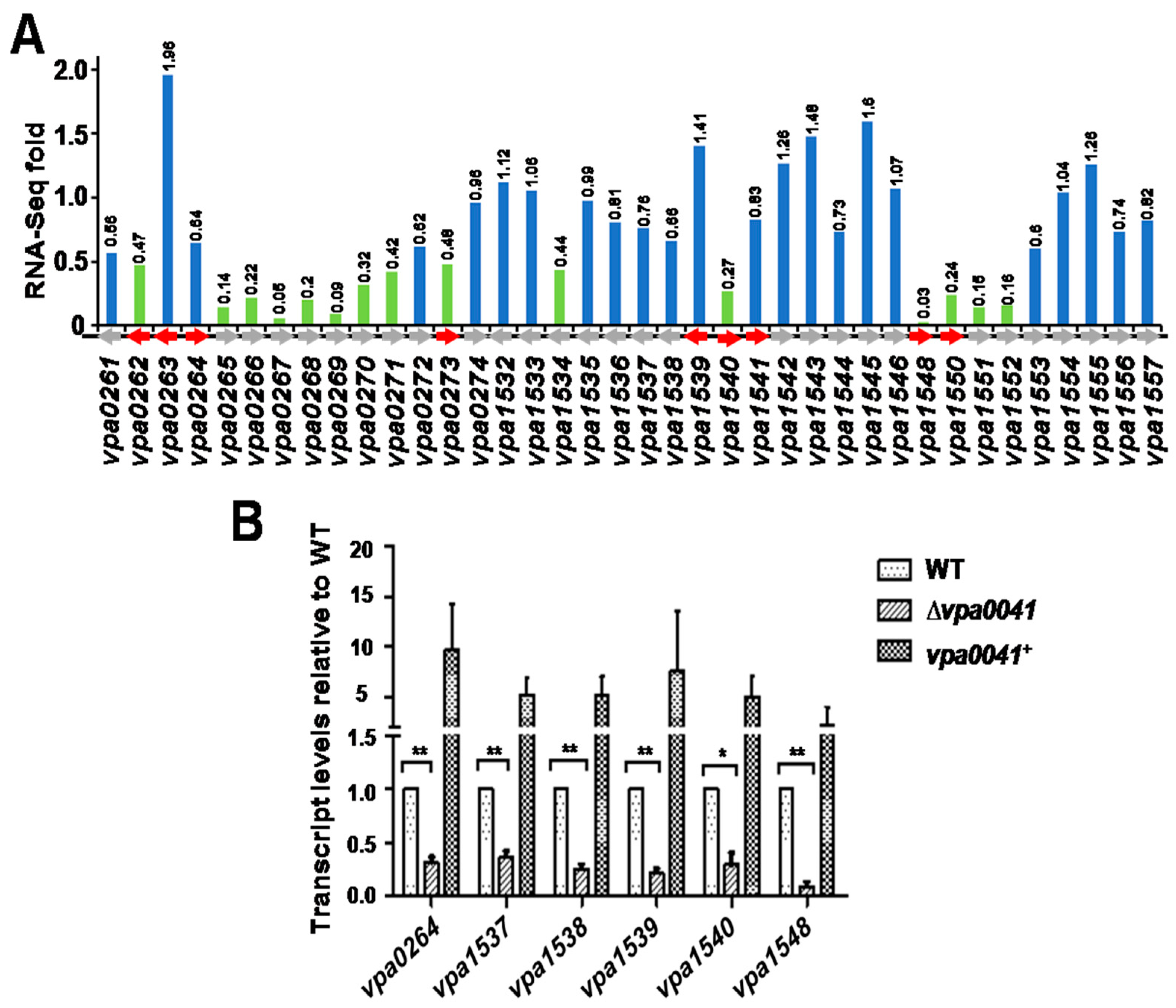
Figure 5.
VPA0041 bound widely to the lateral flagellar promoters. (A–D) EMSA was performed with the DNA fragment from the three lateral flagellar promoters: vpa0264, vpa1548, and vpa1550. The increasing amounts of VPA0041 protein were used as indicated, and 15 ng of each probe with 1 μg non-specific competitor DNA poly(dI:dC). The gyrB gene was used as a negative control.
Figure 5.
VPA0041 bound widely to the lateral flagellar promoters. (A–D) EMSA was performed with the DNA fragment from the three lateral flagellar promoters: vpa0264, vpa1548, and vpa1550. The increasing amounts of VPA0041 protein were used as indicated, and 15 ng of each probe with 1 μg non-specific competitor DNA poly(dI:dC). The gyrB gene was used as a negative control.

Figure 6.
Identification of VPA0041-binding sites in the promoter of vpa0264. (A) The promoter region of vpa0264. Four DNA probes of the vpa0264 promoters were designed to analysis the binding sited of VPA0041. (B) EMSA analysis of VPA0041 binding sites with the various truncations. (C) The binding motif of VPA0041 protein generated by the MEME Suit tool.
Figure 6.
Identification of VPA0041-binding sites in the promoter of vpa0264. (A) The promoter region of vpa0264. Four DNA probes of the vpa0264 promoters were designed to analysis the binding sited of VPA0041. (B) EMSA analysis of VPA0041 binding sites with the various truncations. (C) The binding motif of VPA0041 protein generated by the MEME Suit tool.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial strains and plasmids used in this study.
| Strain or Plasmid | Relevant Characteristics | Reference |
|---|---|---|
| E. coli | ||
| DH5α λpir | Host for π requiring plasmids | [53] |
| SM10 λpir | Host for π requiring plasmids, conjugal donor | [54] |
| BL21(DE3) | Host strain for protein expression | Novagen |
| V. parahaemolyticus | ||
| RIMD 2210633 | Clinical isolate. Carbr | [23] |
| ∆vpa0041 | RIMD 2210633, in-frame deletion in vpa0041, Carbr | This study |
| vpa0041+ | ∆vpa0041, pMMB207 expressing the vpa0041-his gene, Carbr, Cmr | This study |
| Plasmids | ||
| pDM4 | Suicide vector, pir dependent, R6K, SacBR, Cmr | [55] |
| pMMB207 | IncQ lacIq Δbla Ptac-lac lacZa, Cmr | [56] |
| pET30a | Expressing vector, Kmr | Novagen |
| vpa0041::pDM4 | vpa0041 up and down sequences clones into pDM4, Cmr | This study |
| vpa0041::pMMB207 | RBS and vpa0041-his sequences clones into pMMB207, Cmr | This study |
| vpa0041::pET30a | vpa0041 ORF clones into pET30a, Kmr | This study |
Table 2.
Primers used in this study.
| Primer Name | Primer Sequence (5′ to 3′) | Target |
|---|---|---|
| vpa0041-up-F | GATAACAATTTGTGGAATCCCGGGAAGAAGAAATGGGTCAGAAGCGTT | vpa0041 deletion mutant |
| vpa0041-up-R | CGTCCGATGACTTAACATCCATAAACCAACTCCTG | vpa0041 deletion mutant |
| vpa0041-down-F | GGATGTTAAGTCATCGGACGCAAACCATTAAC | vpa0041 deletion mutant |
| vpa0041-down-R | AGTGTATATCAAGCTTATCGATACCCGCACAAAGACAGTGAAGGCAAT | vpa0041 deletion mutant |
| vpa0041-out-F | TTGTTGCGACCGTTTAGCGTATG | vpa0041 deletion mutant |
| vpa0041-out-R | CGCACAAAGACAGTGAAGGCAAT | vpa0041 deletion mutant |
| vpa0041-in-F | TACGCAAGCAGCGGTCAGTG | vpa0041 deletion mutant |
| vpa0041-in-R | GCTCAGAAGCGAAGCGTGTT | vpa0041 deletion mutant |
| pDM4-F | GGTGCTCCAGTGGCTTCTGTTTCTA | vpa0041 deletion mutant |
| pDM4-R | CAGCAACTTAAATAGCCTCTAAT | vpa0041 deletion mutant |
| vpa0041-F | CCGCGAGCTCTAAGGAGGTAGGATAATAATGGATGTTAAGGTCTTTAGAACAT | vpa0041+ |
| vpa0041-R | GCGTCGACTTAGTGATGATGATGATGATGATGGTTTGCGTCCGATGATGC | vpa0041+ |
| pMMB207-F | CTCCCGTTCTGGATAATGTTTTTTG | vpa0041+ |
| pMMB207-R | TCTTCTCTCATCCGCCAAAACAGCC | vpa0041+ |
| vpa0041-his-F | GGAATTCCATATGATGGATGTTAAGGTCTTTAGAAC | VPA0041 expression |
| vpa0041-his-R | GGAATTCCATATGATGGATGTTAAGGTCTTTAGAAC | VPA0041 expression |
| pET30a-F | TAGTTATTGCTCAGCGGTGGC | VPA0041 expression |
| pET30a-R | ACGATGCGTCCGGCGTAGAG | VPA0041 expression |
| gyrb-RT-F | TTACCGTCATGGTGAGCCTG | qRT-PCR |
| gyrb-RT-R | CACGCAGACGTTTTGCTAGG | qRT-PCR |
| vpa0264-RT-F | GCAGGTTCAGGCCCAGTATT | qRT-PCR |
| vpa0264-RT-R | TCATGTTGAGAAACGTCAGGCT | qRT-PCR |
| vpa1537-RT-F | CGCTTGAGAAAACGACAGTGG | qRT-PCR |
| vpa1537-RT-R | CCTACTAATGCGGTCTCGGC | qRT-PCR |
| vpa1538-RT-F | CACGTACGCACATATCCGGT | qRT-PCR |
| vpa1538-RT-R | ACGAACACCTTGCTCAACCT | qRT-PCR |
| vpa1539-RT-F | ATTAGTGAGGGTGCGCCTTT | qRT-PCR |
| vpa1539-RT-R | GGTGAAGGGAAGGAATGGCA | qRT-PCR |
| vpa1540-RT-F | CAACGCCAGTTCGTCTTAACG | qRT-PCR |
| vpa1540-RT-R | ACGGCCAGTAAAGAGAGGTTG | qRT-PCR |
| vpa1548-RT-F | GCTGGTGGCCTTATCGAAGA | qRT-PCR |
| vpa1548-RT-R | TACTGCGAAGTCTGCATCCAT | qRT-PCR |
| FAM-F | FAM-TGCCTGCAGGTCGACGAT | EMSA |
| gyrB-EMSA-F | TGCCTGCAGGTCGACGATTGCCTGCAGGTCGACGATGCGCGCG | EMSA |
| gyrB-EMSA-R | TGCCAGCGCACCGCTGACCGCAG | EMSA |
| vpa0264-EMSA-1F | TGCCTGCAGGTCGACGATGTTTGTCCTGTCGAAAGAATTC | EMSA |
| vpa0264-EMSA-1R | CATGCATCTTTCCTTACAGTCGGCT | EMSA |
| vpa0264-EMSA-2F | TGCCTGCAGGTCGACGATTAGAGTTTTCCCCCTAATTTT | EMSA |
| vpa0264-EMSA-3F | TGCCTGCAGGTCGACGATTCCACTCTTGTTTGTAAGTCAT | EMSA |
| vpa0264-EMSA-4F | TGCCTGCAGGTCGACGATGTATCTTGTTTGTATCTTGGCG | EMSA |
| vpa1548-EMSA-1F | TGCCTGCAGGTCGACGATGCCGATCAAAGCACATCGGAA | EMSA |
| vpa1548-EMSA-1R | CATCTTAGTCTCCTTAGTTTATCAC | EMSA |
| vpa1548-EMSA-2F | TGCCTGCAGGTCGACGATCTTAGTGGAATGCAAGTCACT | EMSA |
| vpa1548-EMSA-3F | TGCCTGCAGGTCGACGATAAATTTTTAATTTTCAAATTA | EMSA |
| vpa1548-EMSA-4F | TGCCTGCAGGTCGACGATAGTGACTAGGGAATATCCCAAG | EMSA |
| vpa1550-EMSA-1F | TGCCTGCAGGTCGACGATTTAGTCGCCTTGTCTTATAGGGA | EMSA |
| vpa1550-EMSA-1R | CACGAGCTTACTCCCTCTCTCATTG | EMSA |
| vpa1550-EMSA-2F | TGCCTGCAGGTCGACGATGGAACACATAAGGTGGAAAATAC | EMSA |
| vpa1550-EMSA-3F | TGCCTGCAGGTCGACGATCAAGTCAATGTTTTAAAAGAAT | EMSA |
| vpa1550-EMSA-4F | TGCCTGCAGGTCGACGATAGACATACTTTCAAGGCATAGAG | EMSA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, M.; Meng, H.; Li, Y.; Gu, D. A Novel Transcription Factor VPA0041 Was Identified to Regulate the Swarming Motility in Vibrio parahaemolyticus. Pathogens 2022, 11, 453. https://doi.org/10.3390/pathogens11040453
AMA Style
Li M, Meng H, Li Y, Gu D. A Novel Transcription Factor VPA0041 Was Identified to Regulate the Swarming Motility in Vibrio parahaemolyticus. Pathogens. 2022; 11(4):453. https://doi.org/10.3390/pathogens11040453
Chicago/Turabian StyleLi, Mingzhu, Hongmei Meng, Yang Li, and Dan Gu. 2022. "A Novel Transcription Factor VPA0041 Was Identified to Regulate the Swarming Motility in Vibrio parahaemolyticus" Pathogens 11, no. 4: 453. https://doi.org/10.3390/pathogens11040453
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.