
Pathogen Spillover to an Invasive Tick Species: First Detection of Bourbon Virus in Haemaphysalis longicornis in the United States



Abstract
:1. Introduction
2. Results
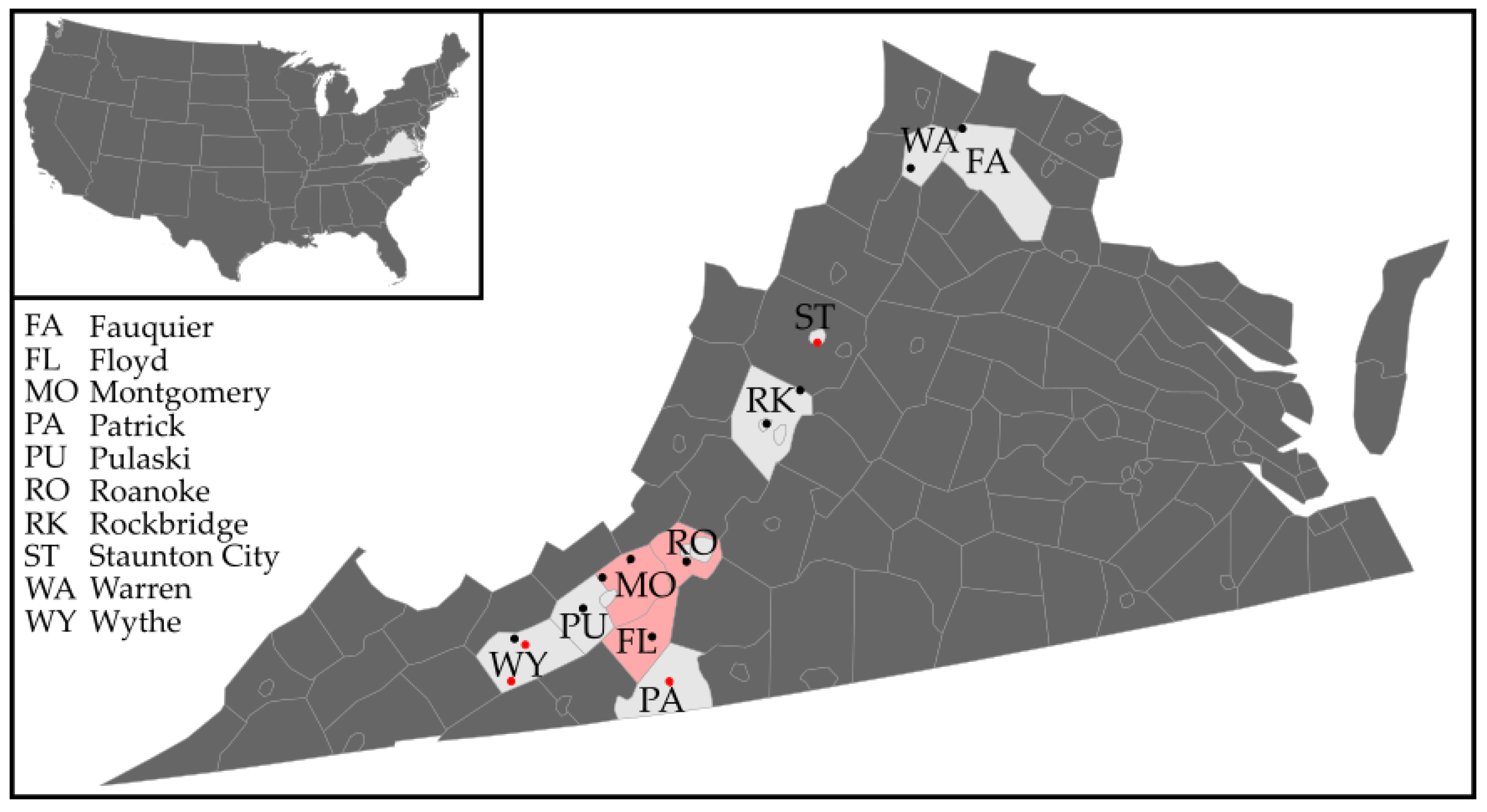
2.1. Haemaphysalis longicornis Collections in Western Virginia
2.2. Arbovirus Detection in Ticks Utilizing Real-Time RT-PCR
2.3. Virus Confirmation and Sequence Analysis
2.4. Tick Homogenate Inoculation on Vero-76 Cells for Attempted BRBV Culturing
2.5. Wildlife Serology
3. Discussion
4. Materials and Methods
4.1. Tick Vollection and Identification
4.2. Whole Nucleic Acid Extraction and Real-Time RT-PCR Detection of BRBV and HRTV
4.3. Sequence Analysis
4.4. Bourbon Virus Isolation Attempt on Cell Culture
4.5. Wildlife Serology
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kosoy, O.I.; Lambert, A.J.; Hawkinson, D.J.; Pastula, D.M.; Goldsmith, C.S.; Hunt, D.C.; Staples, J.E. Novel thogotovirus associated with febrile illness and death, United States, 2014. Emerg. Infect. Dis. 2015, 21, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Savage, H.M.; Burkhalter, K.L.; Godsey, M.S., Jr.; Panella, N.A.; Ashley, D.C.; Nicholson, W.L.; Lambert, A.J. Bourbon virus in field-collected ticks, Missouri, USA. Emerg. Infect. Dis. 2017, 23, 2017–2022. [Google Scholar] [CrossRef]
- Bricker, T.L.; Shafiuddin, M.; Gounder, A.P.; Janowski, A.B.; Zhao, G.; Williams, G.D.; Jagger, B.W.; Diamond, M.S.; Bailey, T.; Kwon, J.H.; et al. Therapeutic efficacy of favipiravir against Bourbon virus in mice. PLoS Pathog. 2019, 15, e1007790. [Google Scholar] [CrossRef]
- Temmam, S.; Monteil-Bouchard, S.; Robert, C.; Baudoin, J.P.; Sambou, M.; Aubadie-Ladrix, M.; Labas, N.; Raoult, D.; Mediannikov, O.; Desnues, C. Characterization of viral communities of biting midges and identification of novel Thogotovirus species and Rhabdovirus genus. Viruses 2016, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Gutiérrez, M.A.; Nunes, M.R.; Guzman, H.; Uribe, S.; Gómez, J.C.G.; Vasco, J.D.S.; Cardoso, J.F.; Popov, V.L.; Widen, S.G.; Wood, T.G.; et al. Sinu virus, a novel and divergent orthomyxovirus related to members of the genus Thogotovirus isolated from mosquitoes in Colombia. Virology 2017, 501, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.R.; Casals, J. Dhori virus, a new agent isolated from Hyalomma dromedarii in India. Indian J. Med. Res. 1973, 61, 1416–1420. [Google Scholar] [PubMed]
- Butenko, A.M.; Leshchinskaia, E.V.; Semashko, I.V.; Donets, M.A.; Martianova, L.I. Dhori virus—A causative agent of human disease. 5 cases of laboratory infection. Vopr. Virusol. 1987, 32, 724–729. [Google Scholar]
- Hubálek, Z.; Rudolf, I. Tick-borne viruses in Europe. Parasitol. Res. 2012, 111, 9–36. [Google Scholar] [CrossRef]
- Moore, D.Á.; Causey, O.R.; Carey, D.E.; Reddy, S.; Cooke, A.R.; Akinkugbe, F.M.; David-West, T.S.; Kemp, G.E. Arthropod-borne viral infections of man in Nigeria, 1964–1970. Ann. Trop. Med. Parasitol. 1975, 69, 49–64. [Google Scholar] [CrossRef]
- Lambert, A.J.; Velez, J.O.; Brault, A.C.; Calvert, A.E.; Bell-Sakyi, L.; Bosco-Lauth, A.M.; Staples, J.E.; Kosoy, O.I. Molecular, serological and in vitro culture-based characterization of Bourbon virus, a newly described human pathogen of the genus Thogotovirus. J. Clin. Virol. 2015, 73, 127–132. [Google Scholar] [CrossRef] [Green Version]
- McMullan, L.K.; Folk, S.M.; Kelly, A.J.; MacNeil, A.; Goldsmith, C.S.; Metcalfe, M.G.; Batten, B.C.; Albarino, C.G.; Zaki, S.R.; Rollin, P.E.; et al. A new phlebovirus associated with severe febrile illness in Missouri. N. Engl. J. Med. 2012, 367, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, H.; Mu, D.; Yin, W.; Yu, H. Epidemiological analysis on severe fever with thrombocytopenia syndrome under the national surveillance data from 2011 to 2014, China. Zhonghua Liu Xing Bing Xue Za Zhi. 2015, 36, 598–602. [Google Scholar] [PubMed]
- Kato, H.; Yamagishi, T.; Shimada, T.; Matsui, T.; Shimojima, M.; Saijo, M.; Oishi, K.; SFTS Epidemiological Research Group-Japan. Epidemiological and clinical features of severe fever with thrombocytopenia syndrome in Japan, 2013–2014. PLoS ONE 2016, 11, e0165207. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Park, S.W.; Bae, I.G.; Kim, S.H.; Ryu, S.Y.; Kim, H.A.; Jang, H.C.; Hur, J.; Jun, J.B.; Jung, Y.; et al. Severe fever with thrombocytopenia syndrome in South Korea, 2013–2015. PLoS Negl. Trop. Dis. 2016, 10, e0005264. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Heartland Virus: Statistics & Maps. Available online: https://www.cdc.gov/heartland-virus/statistics/index.html (accessed on 16 January 2022).
- Muehlenbachs, A.; Fata, C.R.; Lambert, A.J.; Paddock, C.D.; Velez, J.O.; Blau, D.M.; Staples, J.E.; Karlekar, M.B.; Bhatnagar, J.; Nasci, R.S.; et al. Heartland virus–associated death in Tennessee. Clin. Infect. Dis. 2014, 59, 845–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fill, M.M.A.; Compton, M.L.; McDonald, E.C.; Moncayo, A.C.; Dunn, J.R.; Schaffner, W.; Bhatnagar, J.; Zaki, S.R.; Jones, T.F.; Shieh, W.J. Novel clinical and pathologic findings in a Heartland virus–associated death. Clin. Infect. Dis. 2017, 64, 510–512. [Google Scholar] [CrossRef]
- Carlson, A.L.; Pastula, D.M.; Lambert, A.J.; Staples, J.E.; Muehlenbachs, A.; Turabelidze, G.; Eby, C.S.; Keller, J.; Hess, B.; Buller, R.S.; et al. Heartland virus and hemophagocytic lymphohistiocytosis in immunocompromised patient, Missouri, USA. Emerg. Infect. Dis. 2018, 24, 893. [Google Scholar] [CrossRef]
- Savage, H.M.; Godsey, M.S., Jr.; Panella, N.A.; Burkhalter, K.L.; Manford, J.; Trevino-Garrison, I.C.; Straily, A.; Wilson, S.; Bowen, J.; Raghavan, R.K. Surveillance for tick-borne viruses near the location of a fatal human case of bourbon virus (Family Orthomyxoviridae: Genus Thogotovirus) in Eastern Kansas, 2015. J. Med. Entomol. 2018, 55, 701–705. [Google Scholar] [CrossRef]
- Godsey, M.S., Jr.; Rose, D.; Burkhalter, K.L.; Breuner, N.; Bosco-Lauth, A.M.; Kosoy, O.I.; Savage, H.M. Experimental Infection of Amblyomma americanum (Acari: Ixodidae) With Bourbon Virus (Orthomyxoviridae: Thogotovirus). J. Med. Entomol. 2021, 58, 873–879. [Google Scholar] [CrossRef]
- Savage, H.M.; Godsey, M.S., Jr.; Panella, N.A.; Burkhalter, K.L.; Ashley, D.C.; Lash, R.R.; Ramsay, B.; Patterson, T.; Nicholson, W.L. Surveillance for Heartland virus (Bunyaviridae: Phlebovirus) in Missouri during 2013: First detection of virus in adults of Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2016, 53, 607–612. [Google Scholar] [CrossRef]
- Jackson, K.C.; Gidlewski, T.; Root, J.J.; Bosco-Lauth, A.M.; Lash, R.R.; Harmon, J.R.; Brault, A.C.; Panella, N.A.; Nicholson, W.L.; Komar, N. Bourbon virus in wild and domestic animals, Missouri, USA, 2012–2013. Emerg. Infect. Dis. 2019, 25, 1752. [Google Scholar] [CrossRef]
- Komar, N.; Hamby, N.; Palamar, M.B.; Staples, J.E.; Williams, C. Indirect Evidence of Bourbon Virus (Thogotovirus, Orthomyxoviridae) Infection in North Carolina. N. C. Med. J. 2020, 81, 214–215. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Lauth, A.M.; Panella, N.A.; Root, J.J.; Gidlewski, T.; Lash, R.R.; Harmon, J.R.; Burkhalter, K.L.; Godsey, M.S.; Savage, H.M.; Nicholson, W.L.; et al. Serological investigation of heartland virus (Bunyaviridae: Phlebovirus) exposure in wild and domestic animals adjacent to human case sites in Missouri 2012–2013. Am. J. Trop. Med. Hyg. 2015, 92, 1163. [Google Scholar] [CrossRef] [PubMed]
- Riemersma, K.K.; Komar, N. Heartland virus neutralizing antibodies in vertebrate wildlife, United States, 2009–2014. Emerg. Infect. Dis. 2015, 21, 1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, H.M.; Godsey, M.S., Jr.; Lambert, A.; Panella, N.A.; Burkhalter, K.L.; Harmon, J.R.; Lash, R.R.; Ashley, D.C.; Nicholson, W.L. First detection of heartland virus (Bunyaviridae: Phlebovirus) from field collected arthropods. Am. J. Trop. Med. Hyg. 2013, 89, 445. [Google Scholar] [CrossRef]
- Rainey, T.; Occi, J.L.; Robbins, R.G.; Egizi, A. Discovery of Haemaphysalis longicornis (Ixodida: Ixodidae) parasitizing a sheep in New Jersey, United States. J. Med. Entomol. 2018, 55, 757–759. [Google Scholar] [CrossRef]
- USDA-APHIS. Available online: https://www.aphis.usda.gov/animal_health/animal_diseases/tick/downloads/longhorned-tick-sitrep.pdf. (accessed on 9 February 2022).
- Herrin, B.H.; Zajac, A.M.; Little, S.E. Confirmation of Borrelia burgdorferi sensu stricto and Anaplasma phagocytophilum in Ixodes scapularis, Southwestern Virginia. Vector Borne Zoonotic Dis. 2014, 14, 821–823. [Google Scholar] [CrossRef]
- Oakes, V.J.; Todd, S.M.; Carbonello, A.A.; Michalak, P.; Lahmers, K.K. Coinfection of cattle in Virginia with Theileria orientalis Ikeda genotype and Anaplasma marginale. J. Vet. Diagn. Investig. 2022, 34, 36–41. [Google Scholar] [CrossRef]
- Cumbie, A.N.; Whitlow, A.M.; Eastwood, G. First Evidence of Powassan Virus (Flaviviridae) in Ixodes scapularis in Appalachian Virginia, USA. Am. J. Trop. Med. Hyg. 2021. [CrossRef]
- Oakes, V.J.; Yabsley, M.J.; Schwartz, D.; LeRoith, T.; Bissett, C.; Broaddus, C.; Schlater, J.L.; Todd, S.M.; Boes, K.M.; Brookhart, M.; et al. Theileria orientalis Ikeda genotype in cattle, Virginia, USA. Emerg. Infect. Dis. 2019, 25, 1653. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.T.; White, S.; Shaw, D.; Egizi, A.; Lahmers, K.; Ruder, M.G.; Yabsley, M.J. Theileria orientalis Ikeda in host-seeking Haemaphysalis longicornis in Virginia, USA. Ticks Tick Borne Dis. 2020, 11, 101450. [Google Scholar] [CrossRef]
- Stanley, H.M.; Ford, S.L.; Snellgrove, A.N.; Hartzer, K.; Smith, E.B.; Krapiunaya, I.; Levin, M.L. The ability of the invasive Asian longhorned tick Haemaphysalis longicornis (Acari: Ixodidae) to acquire and transmit Rickettsia rickettsii (Rickettsiales: Rickettsiaceae), the agent of Rocky Mountain spotted fever, under laboratory conditions. J. Med. Entomol. 2020, 57, 1635–1639. [Google Scholar] [CrossRef]
- Price, K.J.; Ayres, B.N.; Maes, S.E.; Witmier, B.J.; Chapman, H.A.; Coder, B.L.; Boyer, C.N.; Eisen, R.J.; Nicholson, W.L. First detection of human pathogenic variant of Anaplasma phagocytophilum in field-collected Haemaphysalis longicornis, Pennsylvania, USA. Zoonoses Public Health 2021, 69, 143–148. [Google Scholar] [CrossRef]
- Eastwood, G.; (Virginia Polytechnic Institute and State University, Blacksburg, VA, USA). Unpublished work. 2022.
- Tufts, D.M.; VanAcker, M.C.; Fernandez, M.P.; DeNicola, A.; Egizi, A.; Diuk-Wasser, M.A. Distribution, Host-Seeking Phenology, and Host and Habitat Associations of Haemaphysalis longicornis Ticks, Staten Island, New York, USA. Emerg. Infect. Dis. 2019, 25, 792–796. [Google Scholar] [CrossRef] [Green Version]
- Tufts, D.M.; Goodman, L.B.; Benedict, M.C.; Davis, A.D.; VanAcker, M.C.; Diuk-Wasser, M. Association of the invasive Haemaphysalis longicornis tick with vertebrate hosts, other native tick vectors, and tick-borne pathogens in New York City, USA. Int. J. Parasitol. 2021, 51, 149–157. [Google Scholar] [CrossRef]
- White, S.A.; Bevins, S.N.; Ruder, M.G.; Shaw, D.; Vigil, S.L.; Randall, A.; Deliberto, T.J.; Dominguez, K.; Thompson, A.T.; Mertins, J.W.; et al. Surveys for ticks on wildlife hosts and in the environment at Asian longhorned tick (Haemaphysalis longicornis)-positive sites in Virginia and New Jersey, 2018. Transbound. Emerg. Dis. 2021, 68, 605–614. [Google Scholar] [CrossRef]
- Paddock, C.D.; Yabsley, M.J. Ecological havoc, the rise of white-tailed deer, and the emergence of Amblyomma americanum-associated zoonoses in the United States. Curr. Top. Microbiol. Immunol. 2007, 315, 289–324. [Google Scholar] [CrossRef]
- Eastwood, G.; Shepard, J.J.; Misencik, M.J.; Andreadis, T.G.; Armstrong, P.M. Local persistence of novel regional variants of La Crosse virus in the Northeast USA. Parasit. Vectors. 2020, 13, 1–8. [Google Scholar] [CrossRef]
- Godsey, M.S., Jr.; Savage, H.M.; Burkhalter, K.L.; Bosco-Lauth, A.M.; Delorey, M.J. Transmission of Heartland virus (Bunyaviridae: Phlebovirus) by experimentally infected Ambylomma americanum (Acari: Ixodidae). J. Med. Entomol. 2016, 53, 1226–1233. [Google Scholar] [CrossRef] [Green Version]
- Davies, C.R.; Jones, L.D.; Nuttall, P.A. Experimental studies on the transmission cycle of Thogoto virus, a candidate orthomyxovirus, in Rhipicephalus appendiculatus. Am. J. Trop. Med. Hyg. 1986, 35, 1256–1262. [Google Scholar] [CrossRef]
- Dupuis, A.P.; Prusinski, M.A.; O’Connor, C.; Maffei, J.G.; Ngo, K.A.; Koetzner, C.A.; Santoriello, M.P.; Romano, C.L.; Xu, G.; Ribbe, F.; et al. Heartland Virus Transmission, Suffolk County, New York, USA. Emerg. Infect. Dis. 2021, 27, 3128–3132. [Google Scholar] [CrossRef]
- Pritt, B.S. Haemaphysalis longicornis is in the United States and biting humans: Where do we go from here? Clin. Infect. Dis. 2020, 70, 317–318. [Google Scholar] [CrossRef]
- Cumbie, A.N.; Whitlow, A.M.; Arneson, A.; Du, Z.; Eastwood, G. The distribution, seasonal abundance and environmental factors contributing to the presence of the Asian longhorned tick (Haemaphysalis longicornis, Acari: Ixodidae) in central Appalachian Virginia. J. Med. Entomol. 2022. submitted for publication. [Google Scholar]
- Egizi, A.M.; Robbins, R.G.; Beati, L.; Nava, S. A pictorial key to differentiate the recently detected exotic Haemaphysalis longicornis Neumann, 1901 (Acari, Ixodidae) from native congeners in North America. ZooKeys. 2019, 818, 117. [Google Scholar] [CrossRef] [Green Version]
- Black, W.C.; Piesman, J. Phylogeny of hard-and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [Green Version]
- de la Fuente, J.; Van Den Bussche, R.A.; Kocan, K.M. Molecular phylogeny and biogeography of North American isolates of Anaplasma marginale (Rickettsiaceae: Ehrlichieae). Vet. Parasitol. 2001, 97, 65–76. [Google Scholar] [CrossRef]
- Crowder, C.D.; Rounds, M.A.; Phillipson, C.A.; Picuri, J.M.; Matthews, H.E.; Halverson, J.; Schutzer, S.E.; Ecker, D.J.; Eshoo, M.W. Extraction of total nucleic acids from ticks for the detection of bacterial and viral pathogens. J. Med. Entomol. 2010, 47, 89–94. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Molec. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Eastwood, G.; Sang, R.C.; Guerbois, M.; Taracha, E.L.N.; Weaver, S.C. Enzootic circulation of chikungunya virus in East Africa: Serological evidence in non-human Kenyan primates. Am. J. Trop. Med. Hyg. 2017, 97, 1399–1404. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Number of Collected H. longicornis Ticks | |||||||
|---|---|---|---|---|---|---|---|
| County | Larvae | Nymphs | Females | Total Ticks | Total Pools | No. Pools Tested in This Study | No. Ticks Tested in This Study |
| Fauquier | 7 | 64 | 1 | 72 | 7 | 7 | 72 |
| Floyd | 0 | 1 | 0 | 1 | 1 | 1 | 1 |
| Montgomery | 0 | 2 | 2 | 4 | 3 | 3 | 4 |
| Patrick | 0 | 1 | 0 | 1 | 1 | 1 | 1 |
| Pulaski | 157 1 | 8 | 15 | 180 | 5 | 3 | 18 |
| Rockbridge | 18 | 15 | 0 | 33 | 5 | 5 | 33 |
| Roanoke | 282 | 0 | 0 | 282 | 2 | 2 | 282 |
| Staunton City | 5 | 10 | 0 | 15 | 2 | 2 | 15 |
| Warren | 3 | 15 | 0 | 18 | 2 | 2 | 18 |
| Wythe | 126 | 740 | 297 | 1163 | 80 | 8 | 192 |
| Total | 598 | 856 | 315 | 1769 | 108 | 34 | 636 |
| Sample Description | RT-PCR Screening a | ||||
|---|---|---|---|---|---|
| PB1 Gene (841 bp) | PB2 Gene (357 bp) | PB2 Gene (152 bp) | |||
| Sample ID | VA County | Pool Size and Life Stage (N = Nymph, L = Larvae, F = Adult Female) | C(q) b | C(q) | C(q) |
| RH2-01 | Patrick | 1 N | 38.2, 35.2 | 38.8 | 33.5 |
| ST1-01 | Staunton | 10 N | 38.6, 37.3 | 37.8 | 38.3 |
| WY2-34 | Wythe | 125 L | 39.4, 37.0 | 36.1 | 32.9 |
| WY3-08 | Wythe | 10 F | 38.2, 38.8 | 38.3 | 34.5 |
| Sample ID | County | Tick Sample | Fragment Size | Genbank Accession No. | Comparative Genbank Accession No. (Query Coverage, Sequence Identity) |
|---|---|---|---|---|---|
| RH2-01 | Patrick | Nymph | 418 bp | ON153184 | MT628410 1 (100%, 99.9%) |
| KU708253 2 (100%, 99.9%) | |||||
| ST1-01 | Staunton | Nymph | 345 bp | ON153185 | MK453529 3 (100%, 99.4%) |
| MT628410 1 (100%, 99.4%) | |||||
| WY2-34 | Wythe | Pool of 125 larvae | 418 bp | ON153186 | MT628410 1 (100%, 100%) |
| KU708253 2 (100%, 100%) | |||||
| WY3-08 | Wythe | Female | 405 bp | ON153187 | MT628410 1 (100%, 99.8%) |
| KU708253 2 (100%, 99.8%) |
| County | Species | No. Individuals Sampled | No. Seropositive Samples | Sera Titer Range |
|---|---|---|---|---|
| Floyd | White-tailed deer | 33 | 5 | 1:20–1:80 |
| Northern raccoon | 1 | 0 | - | |
| Striped skunk | 1 | 0 | - | |
| Giles | White-tailed deer | 1 | 0 | - |
| Montgomery | White-tailed deer | 30 | 4 | 1:20–1:160 |
| Striped skunk | 1 | 0 | - | |
| Eastern cottontail | 1 | 0 | - | |
| Pulaski | White-tailed deer | 17 | 0 | - |
| Roanoke | Groundhog | 5 | 3 | 1:40–1:80 |
| Northern raccoon | 3 | 1 | 1:20 |
| Gene Target | Amplicon Size | Primer Name | Primer Sequences (5′–3′) | Reference |
|---|---|---|---|---|
| PB1 (segment 2) | 841 bp | PB1_F | CACCAAGAACATGTCTGAGCC | This study |
| PB1_R | CTCAGTTCACCTGTAACCTCTGCC | |||
| PB2 (segment 1) | 357 bp | PB2_F | GTGCAARAGGGAGGTAGATATTGG | This study |
| PB2_R | CTTTGAGTGATRAGYCCTCGGG | |||
| PB2 (segment 1) | 152 bp | PB2_Inner_F | CAGAATCCTTGATCGGGCCAG | This study |
| PB2_Inner_R | GCATCCTATGGTGCTGAACTGTGG | |||
| HRTV small (S) segment | 86 bp | HRTV1-FOR HRTV1-REV HRTV1-Probe | TGCAGGCTGCTCATTTATTC CCTGTGGAAGAAACCTCTCC 56-FAM/CCTGACCTGTCTCGACTGCCCA /ZEN/-IBFQ | Savage et al., 2013 |
| 16S rRNA | 454 bp | 16S+1 | CTGCTCAATGATTTTTTAAATTGCTGTGG | Black and Piesman, 1994 |
| 16S−1 | CCGGTCTGAACTCAGATCAAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cumbie, A.N.; Trimble, R.N.; Eastwood, G. Pathogen Spillover to an Invasive Tick Species: First Detection of Bourbon Virus in Haemaphysalis longicornis in the United States. Pathogens 2022, 11, 454. https://doi.org/10.3390/pathogens11040454
Cumbie AN, Trimble RN, Eastwood G. Pathogen Spillover to an Invasive Tick Species: First Detection of Bourbon Virus in Haemaphysalis longicornis in the United States. Pathogens. 2022; 11(4):454. https://doi.org/10.3390/pathogens11040454
Chicago/Turabian StyleCumbie, Alexandra N., Rebecca N. Trimble, and Gillian Eastwood. 2022. "Pathogen Spillover to an Invasive Tick Species: First Detection of Bourbon Virus in Haemaphysalis longicornis in the United States" Pathogens 11, no. 4: 454. https://doi.org/10.3390/pathogens11040454