
Metallic Nanoparticles and Core-Shell Nanosystems in the Treatment, Diagnosis, and Prevention of Parasitic Diseases
 , ,
, ,
Abstract
:1. Introduction
2. Different Strategies for Using MNPs and MNPs-Containing Nanosystems in Diagnostics, Prevention, and Therapy of Selected Parasitic Diseases
2.1. Leishmaniasis
2.2. African Trypanosomiasis (African Coma)
2.3. Chagas’ Disease, American Trypanosomiasis
2.4. Toxoplasmosis
2.5. Malaria
2.6. Schistosomiasis
3. Immune Response during Host—Parasite Interaction. Can MNPs and MNPs-Containing Nanosystems Be Used as Immunomodulators during the Course of Parasitic Diseases?
4. Parasitic Invasion of Host Cells as Potential Targets for Nanosystem Action
5. Challenges and Prospects for Future Use of Nanomaterials against Human Parasites
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sitotaw, B.; Mekuriaw, H.; Damtie, D. Prevalence of intestinal parasitic infections and associated risk factors among Jawi primary school children, Jawi town, north-west Ethiopia. BMC Infect. Dis. 2019, 19, 341. [Google Scholar] [CrossRef] [Green Version]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weedall, G.D.; Hall, N. Sexual reproduction and genetic exchange in parasitic protists. Parasitology 2015, 142 (Suppl. S1), S120–S127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, A.S.; Vaughan, A.M.; Kappe, S.H. Malaria parasite development in the mosquito and infection of the mammalian host. Annu. Rev. Microbiol. 2009, 63, 195–221. [Google Scholar] [CrossRef] [Green Version]
- Tyungu, D.L.; McCormick, D.; Lau, C.L.; Chang, M.; Murphy, J.R.; Hotez, P.J.; Mejia, R.; Pollack, H. Toxocara species environmental contamination of public spaces in New York City. PLoS Negl. Trop. Dis. 2020, 14, e0008249. [Google Scholar] [CrossRef] [PubMed]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Public health risks associated with food-borne parasites. EFSA J. 2018, 16, e05495. [Google Scholar] [CrossRef]
- Pereira, A.; Martins, Â.; Brancal, H.; Vilhena, H.; Silva, P.; Pimenta, P.; Diz-Lopes, D.; Neves, N.; Coimbra, M.; Alves, A.C.; et al. Parasitic zoonoses associated with dogs and cats: A survey of Portuguese pet owners’ awareness and deworming practices. Parasit. Vectors 2016, 9, 245. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Tian, L.G.; Liu, Q.; Qian, M.B.; Fu, Q.; Steinmann, P.; Chen, J.X.; Yang, G.J.; Yang, K.; Zhou, X.N. Water-related parasitic diseases in China. Int. J. Environ. Res. Public Health 2013, 10, 1977–2016. [Google Scholar] [CrossRef] [Green Version]
- De Niz, M.; Meehan, G.R.; Brancucci, N.M.B.; Marti, M.; Rotureau, B.; Figueiredo, L.M.; Frischknecht, F. Intravital imaging of host-parasite interactions in skin and adipose tissues. Cell Microbiol. 2019, 21, e13023. [Google Scholar] [CrossRef]
- Richard, R.L.; Ithoi, I.; Abd Majid, M.A.; Wan Sulaiman, W.Y.; Tan, T.C.; Nissapatorn, V.; Lim, Y.A. Monitoring of Waterborne Parasites in Two Drinking Water Treatment Plants: A Study in Sarawak, Malaysia. Int. J. Environ. Res. Public Health 2016, 13, 641. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.J.; Jore, S.; Lund, V.; Grahek-Ogden, D. Risk assessment of parasites in Norwegian drinking water: Opportunities and challenges. Food Waterborne Parasitol. 2021, 22, e00112. [Google Scholar] [CrossRef]
- Tarekegn, Z.S.; Dejene, H.; Addisu, A.; Dagnachew, S. Potential risk factors associated with seropositivity for Toxoplasma gondii among pregnant women and HIV infected individuals in Ethiopia: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2020, 14, e0008944. [Google Scholar] [CrossRef] [PubMed]
- Virus, M.A.; Ehrhorn, E.G.; Lui, L.M.; Davis, P.H. Neurological and neurobehavioral disorders associated with Toxoplasma gondii infection in humans. J. Parasitol. Res. 2021, 2021, 6634807. [Google Scholar] [CrossRef] [PubMed]
- Macchiaroli, N.; Preza, M.; Pérez, M.G.; Kamenetzky, L.; Cucher, M.; Koziol, U.; Castillo, E.; Berriman, M.; Brehm, K.; Rosenzvit, M.C. Expression profiling of Echinococcus multilocularis miRNAs throughout metacestode development in vitro. PLoS Negl. Trop. Dis. 2021, 15, e0009297. [Google Scholar] [CrossRef] [PubMed]
- Utzinger, J.; Raso, G.; Brooker, S.; De Savigny, D.; Tanner, M.; Ornbjerg, N.; Singer, B.H.; N’goran, E.K. Schistosomiasis and neglected tropical diseases: Towards integrated and sustainable control and a word of caution. Parasitology 2009, 136, 1859–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qokoyi, N.K.; Masamba, P.; Kappo, A.P. Proteins as targets in anti-schistosomal drug discovery and vaccine development. Vaccines 2021, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Sato, S. Plasmodium-a brief introduction to the parasites causing human malaria and their basic biology. J. Physiol. Anthropol. 2021, 40, 1. [Google Scholar] [CrossRef]
- Magri, A.; Galuppi, R.; Fioravanti, M. Autochthonous. Pathogens 2021, 10, 334. [Google Scholar] [CrossRef]
- Bouchemal, K.; Bories, C.; Loiseau, P.M. Strategies for prevention and treatment of Trichomonas vaginalis infections. Clin. Microbiol. Rev. 2017, 30, 811–825. [Google Scholar] [CrossRef] [Green Version]
- Leitsch, D. Recent advances in the molecular biology of the protist parasite. Fac. Rev. 2021, 10, 26. [Google Scholar] [CrossRef]
- Amanzougaghene, N.; Mediannikov, O.; Ly, T.D.A.; Gautret, P.; Davoust, B.; Fenollar, F.; Izri, A. Molecular investigation and genetic diversity of Pediculus and Pthirus lice in France. Parasit. Vectors 2020, 13, 177. [Google Scholar] [CrossRef] [Green Version]
- Arrúa, E.C.; Seremeta, K.P.; Bedogni, G.R.; Okulik, N.B.; Salomon, C.J. Nanocarriers for effective delivery of benznidazole and nifurtimox in the treatment of chagas disease: A review. Acta Trop. 2019, 198, 105080. [Google Scholar] [CrossRef]
- Carrion, C.; Robles, N.; Sola-Morales, O.; Aymerich, M.; Ruiz Postigo, J.A. Mobile health strategies to tackle skin neglected tropical diseases with recommendations from innovative experiences: Systematic review. JMIR Mhealth Uhealth 2020, 8, e22478. [Google Scholar] [CrossRef]
- Volpedo, G.; Costa, L.; Ryan, N.; Halsey, G.; Satoskar, A.; Oghumu, S. Nanoparticulate drug delivery systems for the treatment of neglected tropical protozoan diseases. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e144118. [Google Scholar] [CrossRef] [Green Version]
- Al Jalali, V.; Zeitlinger, M. Systemic and target-site pharmacokinetics of antiparasitic agents. Clin. Pharmacokinet. 2020, 59, 827–847. [Google Scholar] [CrossRef] [Green Version]
- Sarangi, B.; Jana, U.; Sahoo, J.; Mohanta, G.P.; Manna, P.K. Systematic approach for the formulation and optimization of atorvastatin loaded solid lipid NANOAPARTICLES using response surface methodology. Biomed. Microdevices 2018, 20, 53. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chen, D.; Pan, Y.; Qu, W.; Hao, H.; Wang, X.; Liu, Z.; Xie, S. Nanoparticles for antiparasitic drug delivery. Drug Deliv. 2019, 26, 1206–1221. [Google Scholar] [CrossRef] [PubMed]
- Deda, D.K.; Iglesias, B.A.; Alves, E.; Araki, K.; Garcia, C.R.S. Porphyrin derivative nanoformulations for therapy and antiparasitic agents. Molecules 2020, 25, 2080. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.X.; Weller, P.F. Antiparasitic drugs. N. Engl. J. Med. 1996, 334, 1178–1184. [Google Scholar] [CrossRef]
- Franco, J.R.; Simarro, P.P.; Diarra, A.; Jannin, J.G. Epidemiology of human African trypanosomiasis. Clin. Epidemiol. 2014, 6, 257–275. [Google Scholar] [CrossRef]
- Altcheh, J.; Castro, L.; Dib, J.C.; Grossmann, U.; Huang, E.; Moscatelli, G.; Pinto Rocha, J.J.; Ramírez, T.E.; Group, C.S. Prospective, historically controlled study to evaluate the efficacy and safety of a new paediatric formulation of nifurtimox in children aged 0 to 17 years with Chagas disease one year after treatment (CHICO). PLoS Negl. Trop. Dis. 2021, 15, e0008912. [Google Scholar] [CrossRef] [PubMed]
- Skłodowski, K.; Chmielewska-Deptuła, S.J.; Piktel, E.; Wolak, P.; Wollny, T.; Bucki, R. Metallic nanosystems in the development of antimicrobial strategies with high antimicrobial activity and high biocompatibility. Int. J. Mol. Sci. 2023, 24, 2104. [Google Scholar] [CrossRef] [PubMed]
- Marques Neto, L.M.; Kipnis, A.; Junqueira-Kipnis, A.P. Role of metallic nanoparticles in vaccinology: Implications for infectious disease vaccine development. Front. Immunol. 2017, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Niemirowicz, K.; Piktel, E.; Wilczewska, A.Z.; Markiewicz, K.H.; Durnaś, B.; Wątek, M.; Puszkarz, I.; Wróblewska, M.; Niklińska, W.; Savage, P.B.; et al. Core-shell magnetic nanoparticles display synergistic antibacterial effects against. Int. J. Nanomed. 2016, 11, 5443–5455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piktel, E.; Niemirowicz, K.; Wątek, M.; Wollny, T.; Deptuła, P.; Bucki, R. Recent insights in nanotechnology-based drugs and formulations designed for effective anti-cancer therapy. J. Nanobiotechnol. 2016, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Khezerlou, A.; Alizadeh-Sani, M.; Azizi-Lalabadi, M.; Ehsani, A. Nanoparticles and their antimicrobial properties against pathogens including bacteria, fungi, parasites and viruses. Microb. Pathog. 2018, 123, 505–526. [Google Scholar] [CrossRef] [PubMed]
- Durnaś, B.; Wnorowska, U.; Pogoda, K.; Deptuła, P.; Wątek, M.; Piktel, E.; Głuszek, S.; Gu, X.; Savage, P.B.; Niemirowicz, K.; et al. Candidacidal activity of selected ceragenins and human cathelicidin LL-37 in experimental settings mimicking infection sites. PLoS ONE 2016, 11, e0157242. [Google Scholar] [CrossRef]
- Durnaś, B.; Piktel, E.; Wątek, M.; Wollny, T.; Góźdź, S.; Smok-Kalwat, J.; Niemirowicz, K.; Savage, P.B.; Bucki, R. Anaerobic bacteria growth in the presence of cathelicidin LL-37 and selected ceragenins delivered as magnetic nanoparticles cargo. BMC Microbiol. 2017, 17, 167. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.S.; Othman, R.Y. Recent advances in Toxoplasma gondii immunotherapeutics. Korean J. Parasitol. 2014, 52, 581–593. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Han, Y.; Kato, K. Exploring amino acid-capped nanoparticles for selective anti-parasitic action and improved host biocompatibility. J. Biomed. Nanotechnol. 2018, 14, 847–867. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Han, Y.; Kato, K. Nanoparticles show potential to retard bradyzoites in vitro formation of Toxoplasma gondii. Folia Parasitol. 2019, 66, 1–6. [Google Scholar] [CrossRef]
- MubarakAli, D.; Thajuddin, N.; Jeganathan, K.; Gunasekaran, M. Plant extract mediated synthesis of silver and gold nanoparticles and its antibacterial activity against clinically isolated pathogens. Colloids Surf. B Biointerfaces 2011, 85, 360–365. [Google Scholar] [CrossRef]
- Huynh, K.H.; Pham, X.H.; Kim, J.; Lee, S.H.; Chang, H.; Rho, W.Y.; Jun, B.H. Synthesis, properties, and biological applications of metallic alloy nanoparticles. Int. J. Mol. Sci. 2020, 21, 5174. [Google Scholar] [CrossRef]
- Yetisgin, A.A.; Cetinel, S.; Zuvin, M.; Kosar, A.; Kutlu, O. Therapeutic nanoparticles and their targeted delivery applications. Molecules 2020, 25, 2193. [Google Scholar] [CrossRef]
- Torres-Sangiao, E.; Holban, A.M.; Gestal, M.C. Advanced nanobiomaterials: Vaccines, diagnosis and treatment of infectious diseases. Molecules 2016, 21, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinsiku, A.A.; Dare, E.O.; Ajani, O.O.; Adekoya, J.A.; Adeyemi, A.O.; Ejilude, O.; Oyeyemi, K.D. Dataset on the evaluation of antimicrobial activity and optical properties of green synthesized silver and its allied bimetallic nanoparticles. Data Brief 2018, 21, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Riezk, A.; Van Bocxlaer, K.; Yardley, V.; Murdan, S.; Croft, S.L. Activity of amphotericin B-loaded chitosan nanoparticles against experimental cutaneous leishmaniasis. Molecules 2020, 25, 4002. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Acosta, C.A.; Ramírez-Salcedo, J.; Morales-Serna, F.N.; Fajer-Ávila, E.J.; Chávez-Sánchez, C.; Lara, H.H.; García-Gasca, A. Molecular effects of silver nanoparticles on monogenean parasites: Lessons from Caenorhabditis elegans. Int. J. Mol. Sci. 2020, 21, 5889. [Google Scholar] [CrossRef] [PubMed]
- Petros, R.A.; DeSimone, J.M. Strategies in the design of nanoparticles for therapeutic applications. Nat. Rev. Drug Discov. 2010, 9, 615–627. [Google Scholar] [CrossRef]
- Kong, F.Y.; Zhang, J.W.; Li, R.F.; Wang, Z.X.; Wang, W.J.; Wang, W. Unique roles of gold nanoparticles in drug delivery, targeting and imaging applications. Molecules 2017, 22, 1445. [Google Scholar] [CrossRef] [Green Version]
- Dadfar, S.M.; Roemhild, K.; Drude, N.I.; von Stillfried, S.; Knüchel, R.; Kiessling, F.; Lammers, T. Iron oxide nanoparticles: Diagnostic, therapeutic and theranostic applications. Adv. Drug Deliv. Rev. 2019, 138, 302–325. [Google Scholar] [CrossRef] [PubMed]
- Benamrouz, S.; Conseil, V.; Creusy, C.; Calderon, E.; Dei-Cas, E.; Certad, G. Parasites and malignancies, a review, with emphasis on digestive cancer induced by Cryptosporidium parvum (Alveolata: Apicomplexa). Parasite 2012, 19, 101–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, J.E. HIV-related cerebral toxoplasmosis revisited: Current concepts and controversies of an old disease. J. Int. Assoc. Provid. AIDS Care 2019, 18, 2325958219867315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, H.H.; Nash, T.E.; Del Brutto, O.H. Clinical symptoms, diagnosis, and treatment of neurocysticercosis. Lancet Neurol. 2014, 13, 1202–1215. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, M.; Hunter, P.R.; Chalmers, R.M.; Tyler, K.M. Cryptosporidium pathogenicity and virulence. Clin. Microbiol. Rev. 2013, 26, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Adamu, H.; Petros, B.; Zhang, G.; Kassa, H.; Amer, S.; Ye, J.; Feng, Y.; Xiao, L. Distribution and clinical manifestations of Cryptosporidium species and subtypes in HIV/AIDS patients in Ethiopia. PLoS Negl. Trop. Dis. 2014, 8, e2831. [Google Scholar] [CrossRef] [Green Version]
- Moyano, A.; Serrano-Pertierra, E.; Salvador, M.; Martínez-García, J.C.; Rivas, M.; Blanco-López, M.C. Magnetic lateral flow immunoassays. Diagnostics 2020, 10, 288. [Google Scholar] [CrossRef]
- Thakur, S.; Joshi, J.; Kaur, S. Leishmaniasis diagnosis: An update on the use of parasitological, immunological and molecular methods. J. Parasit. Dis. 2020, 44, 253–272. [Google Scholar] [CrossRef]
- Tangpukdee, N.; Duangdee, C.; Wilairatana, P.; Krudsood, S. Malaria diagnosis: A brief review. Korean J. Parasitol. 2009, 47, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Tallury, P.; Malhotra, A.; Byrne, L.M.; Santra, S. Nanobioimaging and sensing of infectious diseases. Adv. Drug Deliv. Rev. 2010, 62, 424–437. [Google Scholar] [CrossRef]
- Baptista, V.; Peng, W.K.; Minas, G.; Veiga, M.I.; Catarino, S.O. Review of microdevices for hemozoin-based malaria detection. Biosensors 2022, 12, 110. [Google Scholar] [CrossRef]
- Nash, M.A.; Waitumbi, J.N.; Hoffman, A.S.; Yager, P.; Stayton, P.S. Multiplexed enrichment and detection of malarial biomarkers using a stimuli-responsive iron oxide and gold nanoparticle reagent system. ACS Nano 2012, 6, 6776–6785. [Google Scholar] [CrossRef] [Green Version]
- Adekiya, T.A.; Kondiah, P.P.D.; Choonara, Y.E.; Kumar, P.; Pillay, V. A review of nanotechnology for targeted anti-schistosomal therapy. Front. Bioeng. Biotechnol. 2020, 8, 32. [Google Scholar] [CrossRef]
- Bruni, N.; Stella, B.; Giraudo, L.; Della Pepa, C.; Gastaldi, D.; Dosio, F. Nanostructured delivery systems with improved leishmanicidal activity: A critical review. Int. J. Nanomed. 2017, 12, 5289–5311. [Google Scholar] [CrossRef] [Green Version]
- Varela-Aramburu, S.; Ghosh, C.; Goerdeler, F.; Priegue, P.; Moscovitz, O.; Seeberger, P.H. Targeting and Inhibiting. ACS Appl. Mater. Interfaces 2020, 12, 43380–43387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Zhang, Y.L.; Zhou, J.P.; Lv, H.X. Solid lipid nanoparticles modified with stearic acid-octaarginine for oral administration of insulin. Int. J. Nanomed. 2012, 7, 3333–3339. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhong, X.; Zhang, X.; Lv, W.; Yu, J.; Xiao, Y.; Liu, S.; Huang, J. Preparation of intravenous injection nanoformulation via co-assemble between cholesterylated gemcitabine and cholesterylated mPEG: Enhanced cellular uptake and intracellular drug controlled release. J. Microencapsul. 2017, 34, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Mikušová, V.; Mikuš, P. Advances in chitosan-based nanoparticles for drug delivery. Int. J. Mol. Sci. 2021, 22, 9652. [Google Scholar] [CrossRef] [PubMed]
- Ghasemiyeh, P.; Mohammadi-Samani, S. Potential of Nanoparticles as permeation enhancers and targeted delivery options for skin: Advantages and disadvantages. Drug Des. Dev. Ther. 2020, 14, 3271–3289. [Google Scholar] [CrossRef]
- Luo, Y.H.; Chang, L.W.; Lin, P. Metal-based nanoparticles and the immune system: Activation, inflammation, and potential applications. Biomed. Res. Int. 2015, 2015, 143720. [Google Scholar] [CrossRef] [Green Version]
- Reddy, N.; Rapisarda, M. Properties and applications of nanoparticles from plant proteins. Materials 2021, 14, 3607. [Google Scholar] [CrossRef]
- Elgqvist, J. Nanoparticles as theranostic vehicles in experimental and clinical applications-focus on prostate and breast cancer. Int. J. Mol. Sci. 2017, 18, 1102. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, F.M.S.; Cruz, R.E.; Pinheiro, G.R.G.; Caliari, M.V. Comorbidities involving parasitic diseases: A look at the benefits and complications. Exp. Biol. Med. 2022, 247, 1819–1826. [Google Scholar] [CrossRef]
- Amina, S.J.; Guo, B. A review on the synthesis and functionalization of gold nanoparticles as a drug delivery vehicle. Int. J. Nanomed. 2020, 15, 9823–9857. [Google Scholar] [CrossRef]
- Genetu Bayih, A.; Debnath, A.; Mitre, E.; Huston, C.D.; Laleu, B.; Leroy, D.; Blasco, B.; Campo, B.; Wells, T.N.C.; Willis, P.A.; et al. Susceptibility testing of medically important parasites. Clin. Microbiol. Rev. 2017, 30, 647–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aderibigbe, B.A. Metal-based nanoparticles for the treatment of infectious diseases. Molecules 2017, 22, 1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyemi, O.S.; Molefe, N.I.; Awakan, O.J.; Nwonuma, C.O.; Alejolowo, O.O.; Olaolu, T.; Maimako, R.F.; Suganuma, K.; Han, Y.; Kato, K. Metal nanoparticles restrict the growth of protozoan parasites. Artif. Cells Nanomed. Biotechnol. 2018, 46, S86–S94. [Google Scholar] [CrossRef] [Green Version]
- Nadhman, A.; Khan, M.I.; Nazir, S.; Khan, M.; Shahnaz, G.; Raza, A.; Shams, D.F.; Yasinzai, M. Annihilation of Leishmania by daylight responsive ZnO nanoparticles: A temporal relationship of reactive oxygen species-induced lipid and protein oxidation. Int. J. Nanomed. 2016, 11, 2451–2461. [Google Scholar] [CrossRef] [Green Version]
- Delavari, M.; Dalimi, A.; Ghaffarifar, F.; Sadraei, J. In vitro study on cytotoxic effects of ZnO nanoparticles on promastigote and amastigote forms of Leishmania major (MRHO/IR/75/ER). Iran. J. Parasitol. 2014, 9, 6–13. [Google Scholar]
- Jebali, A.; Kazemi, B. Nano-based antileishmanial agents: A toxicological study on nanoparticles for future treatment of cutaneous leishmaniasis. Toxicol. Vitr. 2013, 27, 1896–1904. [Google Scholar] [CrossRef] [PubMed]
- Fanti, J.R.; Tomiotto-Pellissier, F.; Miranda-Sapla, M.M.; Cataneo, A.H.D.; Andrade, C.G.T.J.; Panis, C.; Rodrigues, J.H.D.S.; Wowk, P.F.; Kuczera, D.; Costa, I.N.; et al. Biogenic silver nanoparticles inducing Leishmania amazonensis promastigote and amastigote death in vitro. Acta Trop. 2018, 178, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Rahul, S.; Chandrashekhar, P.; Hemant, B.; Bipinchandra, S.; Mouray, E.; Grellier, P.; Satish, P. In vitro antiparasitic activity of microbial pigments and their combination with phytosynthesized metal nanoparticles. Parasitol. Int. 2015, 64, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Perinoto, Â.; Maki, R.M.; Colhone, M.C.; Santos, F.R.; Migliaccio, V.; Daghastanli, K.R.; Stabeli, R.G.; Ciancaglini, P.; Paulovich, F.V.; de Oliveira, M.C.F.; et al. Biosensors for efficient diagnosis of leishmaniasis: Innovations in bioanalytics for a neglected disease. Anal. Chem. 2010, 82, 9763–9768. [Google Scholar] [CrossRef] [PubMed]
- Alti, D.; Veeramohan Rao, M.; Rao, D.N.; Maurya, R.; Kalangi, S.K. Gold-Silver bimetallic nanoparticles reduced with herbal leaf extracts induce ROS-mediated death in both promastigote and amastigote stages of Leishmania donovani. ACS Omega 2020, 5, 16238–16245. [Google Scholar] [CrossRef]
- Sumaira; Salman Hashmi, S.; Ali, G.S.; Zia, M.; Haider Abbasi, B. Comparative antileishmanial efficacy of the biosynthesised ZnO NPs from genus Verbena. IET Nanobiotechnol. 2018, 12, 1067–1073. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Khalil, M.F.; Bauomy, A.A.; Diab, M.S.; Al-Quraishy, S. Efficacy of gold nanoparticles against nephrotoxicity induced by Schistosoma mansoni infection in mice. Biomed. Environ. Sci. 2016, 29, 773–781. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Khalil, M.F.; Diab, M.S.M.; Bauomy, A.A.; Al-Quraishy, S. Effect of gold nanoparticles on mice splenomegaly induced by schistosomiasis mansoni. Saudi J. Biol. Sci. 2017, 24, 1418–1423. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Lin, Y.; Weng, R.; Chen, R.; Li, J.; Lv, Z. Diagnosis, monitoring, and control of schistosomiasis-an update. J. Biomed. Nanotechnol. 2018, 14, 430–455. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Bauomy, A.A.; Diab, M.S.; Wahab, R.; Delic, D.; Al-Quraishy, S. Impact of gold nanoparticles on brain of mice infected with Schistosoma mansoni. Parasitol. Res. 2015, 114, 3711–3719. [Google Scholar] [CrossRef]
- Hendiger, E.B.; Padzik, M.; Sifaoui, I.; Reyes-Batlle, M.; López-Arencibia, A.; Rizo-Liendo, A.; Bethencourt-Estrella, C.J.; Nicolás-Hernández, D.S.; Chiboub, O.; Rodríguez-Expósito, R.L.; et al. Silver Nanoparticles as a novel potential preventive agent against Acanthamoeba keratitis. Pathogens 2020, 9, 350. [Google Scholar] [CrossRef]
- Barabadi, H.; Alizadeh, Z.; Rahimi, M.T.; Barac, A.; Maraolo, A.E.; Robertson, L.J.; Masjedi, A.; Shahrivar, F.; Ahmadpour, E. Nanobiotechnology as an emerging approach to combat malaria: A systematic review. Nanomedicine 2019, 18, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Murugan, K.; Panneerselvam, C.; Subramaniam, J.; Madhiyazhagan, P.; Hwang, J.S.; Wang, L.; Dinesh, D.; Suresh, U.; Roni, M.; Higuchi, A.; et al. Eco-friendly drugs from the marine environment: Spongeweed-synthesized silver nanoparticles are highly effective on Plasmodium falciparum and its vector Anopheles stephensi, with little non-target effects on predatory copepods. Environ. Sci. Pollut. Res. Int. 2016, 23, 16671–16685. [Google Scholar] [CrossRef] [PubMed]
- Murugan, K.; Panneerselvam, C.; Samidoss, C.M.; Madhiyazhagan, P.; Suresh, U.; Roni, M.; Chandramohan, B.; Subramaniam, J.; Dinesh, D.; Rajaganesh, R.; et al. In vivo and in vitro effectiveness of Azadirachta indica-synthesized silver nanocrystals against Plasmodium berghei and Plasmodium falciparum, and their potential against malaria mosquitoes. Res. Vet. Sci. 2016, 106, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, A.; Murugan, K.; Panneerselvam, C.; Madhiyazhagan, P.; Dinesh, D.; Vadivalagan, C.; Aziz, A.T.; Chandramohan, B.; Suresh, U.; Rajaganesh, R.; et al. Earthworm-mediated synthesis of silver nanoparticles: A potent tool against hepatocellular carcinoma, Plasmodium falciparum parasites and malaria mosquitoes. Parasitol. Int. 2016, 65, 276–284. [Google Scholar] [CrossRef]
- Mishra, A.; Kaushik, N.K.; Sardar, M.; Sahal, D. Evaluation of antiplasmodial activity of green synthesized silver nanoparticles. Colloids Surf. B Biointerfaces 2013, 111, 713–718. [Google Scholar] [CrossRef]
- Antczak, M.; Dzitko, K.; Długońska, H. Human toxoplasmosis—Searching for novel chemotherapeutics. Biomed. Pharmacother. 2016, 82, 677–684. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, Q.; Shen, H.; Zhang, Y.; Zhang, S.; Li, X.; Acharya, G. Seroepidemiological map of Toxoplasma gondii infection and associated risk factors in preconception period in China: A nationwide cross-sectional study. J. Obstet. Gynaecol. Res. 2018, 44, 1134–1139. [Google Scholar] [CrossRef]
- Abdellahi, L.; Iraji, F.; Mahmoudabadi, A.; Hejazi, S.H. Vaccination in Leishmaniasis: A review article. Iran. Biomed. J. 2022, 26, 1–35. [Google Scholar] [CrossRef]
- Pace, D.; Williams, T.N.; Grochowska, A.; Betts, A.; Attard-Montalto, S.; Boffa, M.J.; Vella, C. Manifestations of paediatric Leishmania infantum infections in Malta. Travel. Med. Infect. Dis. 2011, 9, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Scorza, B.M.; Carvalho, E.M.; Wilson, M.E. Cutaneous manifestations of human and murine Leishmaniasis. Int. J. Mol. Sci. 2017, 18, 1296. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, M.G.; Pagliari, C.; Queiroz, A.; Machado, P.R.L.; Carvalho, L.; Scott, P.; Carvalho, E.M.; Arruda, S. Tissue damage in human cutaneous Leishmaniasis: Correlations between inflammatory cells and molecule expression. Front. Cell. Infect. Microbiol. 2020, 10, 355. [Google Scholar] [CrossRef]
- Saleem, K.; Khursheed, Z.; Hano, C.; Anjum, I.; Anjum, S. Applications of nanomaterials in Leishmaniasis: A focus on recent advances and challenges. Nanomaterials 2019, 9, 1749. [Google Scholar] [CrossRef] [Green Version]
- Nafari, A.; Cheraghipour, K.; Sepahvand, M.; Shahrokhi, G.; Gabal, E.; Mahmoudvand, H. Nanoparticles: New agents toward treatment of leishmaniasis. Parasite Epidemiol. Control 2020, 10, e00156. [Google Scholar] [CrossRef]
- Ali, A.; Ambreen, S.; Javed, R.; Tabassum, S.; Ul Haq, I.; Zia, M. ZnO nanostructure fabrication in different solvents transforms physio-chemical, biological and photodegradable properties. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 74, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Allahverdiyev, A.M.; Abamor, E.S.; Bagirova, M.; Ustundag, C.B.; Kaya, C.; Kaya, F.; Rafailovich, M. Antileishmanial effect of silver nanoparticles and their enhanced antiparasitic activity under ultraviolet light. Int. J. Nanomed. 2011, 6, 2705–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadpour, R.; Dobrovolskaia, M.A.; Cheney, D.L.; Greish, K.F.; Ghandehari, H. Subchronic and chronic toxicity evaluation of inorganic nanoparticles for delivery applications. Adv. Drug Deliv. Rev. 2019, 144, 112–132. [Google Scholar] [CrossRef]
- El-Khadragy, M.; Alolayan, E.M.; Metwally, D.M.; El-Din, M.F.S.; Alobud, S.S.; Alsultan, N.I.; Alsaif, S.S.; Awad, M.A.; Abdel Moneim, A.E. Clinical efficacy associated with enhanced antioxidant enzyme activities of silver nanoparticles biosynthesized using Moringa oleifera leaf extract, against cutaneous Leishmaniasis in a murine model of Leishmania major. Int. J. Environ. Res. Public Health 2018, 15, 1037. [Google Scholar] [CrossRef] [Green Version]
- Malapermal, V.; Botha, I.; Krishna, S.B.N.; Mbatha, J.N. Enhancing antidiabetic and antimicrobial performance of. Saudi J. Biol. Sci. 2017, 24, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Swami, R.K.; Narula, A. Harnessing the potential of roots of traditional power plant: Ocimum. Front. Plant Sci. 2021, 12, 765024. [Google Scholar] [CrossRef]
- Salassa, B.N.; Romano, P.S. Autophagy: A necessary process during the. Virulence 2019, 10, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, O.S.; Arowolo, A.T.; Hetta, H.F.; Al-Rejaie, S.; Rotimi, D.; Batiha, G.E. Apoferritin and apoferritin-capped metal nanoparticles inhibit Arginine kinase of of Trypanosoma brucei. Molecules 2020, 25, 3432. [Google Scholar] [CrossRef]
- Mulenga, P.; Chenge, F.; Boelaert, M.; Mukalay, A.; Lutumba, P.; Lumbala, C.; Luboya, O.; Coppieters, Y. Integration of human african trypanosomiasis control activities into primary healthcare services: A scoping review. Am. J. Trop. Med. Hyg. 2019, 101, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Onyilagha, C.; Uzonna, J.E. Host immune responses and immune evasion strategies in African trypanosomiasis. Front. Immunol. 2019, 10, 2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimani, N.M.; Backhaus, S.; Matasyoh, J.C.; Kaiser, M.; Herrmann, F.C.; Schmidt, T.J.; Langer, K. Preparation of sesquiterpene lactone-loaded PLA nanoparticles and evaluation of their Antitrypanosomal activity. Molecules 2019, 24, 2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, J.; Steiner, I.; Kennedy, P.G.E. Generation of neuroinflammation in human African trypanosomiasis. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baptista, P.V.; McCusker, M.P.; Carvalho, A.; Ferreira, D.A.; Mohan, N.M.; Martins, M.; Fernandes, A.R. Nano-strategies to fight multidrug resistant bacteria—“A Battle of the Titans”. Front. Microbiol. 2018, 9, 1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albalawi, A.E.; Alanazi, A.D.; Baharvand, P.; Sepahvand, M.; Mahmoudvand, H. High Potency of organic and inorganic nanoparticles to treat cystic echinococcosis: An evidence-based review. Nanomaterials 2020, 10, 2538. [Google Scholar] [CrossRef]
- Echeverría, L.E.; Marcus, R.; Novick, G.; Sosa-Estani, S.; Ralston, K.; Zaidel, E.J.; Forsyth, C.; RIbeiro, A.L.P.; Mendoza, I.; Falconi, M.L.; et al. WHF IASC roadmap on chagas disease. Glob. Heart 2020, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, V.; Dias, N.; Paiva, T.; Hagström-Bex, L.; Nitz, N.; Pratesi, R.; Hecht, M. Current trends in the pharmacological management of Chagas disease. Int. J. Parasitol. Drugs Drug Resist. 2020, 12, 7–17. [Google Scholar] [CrossRef]
- Dumonteil, E.; Herrera, C.; Buekens, P. A therapeutic preconceptional vaccine against Chagas disease: A novel indication that could reduce congenital transmission and accelerate vaccine development. PLoS Negl. Trop. Dis. 2019, 13, e0006985. [Google Scholar] [CrossRef] [Green Version]
- Costales, J.A.; Sánchez-Gómez, A.; Silva-Aycaguer, L.C.; Cevallos, W.; Tamayo, S.; Yumiseva, C.A.; Jacobson, J.O.; Martini, L.; Carrera, C.A.; Grijalva, M.J. A national survey to determine prevalence of Trypanosoma cruzi infection among pregnant women in Ecuador. Am. J. Trop. Med. Hyg. 2015, 92, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Quijia Quezada, C.; Azevedo, C.S.; Charneau, S.; Santana, J.M.; Chorilli, M.; Carneiro, M.B.; Bastos, I.M.D. Advances in nanocarriers as drug delivery systems in Chagas disease. Int. J. Nanomed. 2019, 14, 6407–6424. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.D. Nano-medicines a hope for Chagas disease! Front. Mol. Biosci. 2021, 8, 655435. [Google Scholar] [CrossRef] [PubMed]
- Mazzeti, A.L.; Capelari-Oliveira, P.; Bahia, M.T.; Mosqueira, V.C.F. Review on experimental treatment strategies against Trypanosoma cruzi. J. Exp. Pharmacol. 2021, 13, 409–432. [Google Scholar] [CrossRef] [PubMed]
- Olivera, M.J.; Cucunubá, Z.M.; Valencia-Hernández, C.A.; Herazo, R.; Agreda-Rudenko, D.; Flórez, C.; Duque, S.; Nicholls, R.S. Risk factors for treatment interruption and severe adverse effects to benznidazole in adult patients with Chagas disease. PLoS ONE 2017, 12, e0185033. [Google Scholar] [CrossRef] [PubMed]
- Tanowitz, H.B.; Machado, F.S.; Spray, D.C.; Friedman, J.M.; Weiss, O.S.; Lora, J.N.; Nagajyothi, J.; Moraes, D.N.; Garg, N.J.; Nunes, M.C.; et al. Developments in the management of Chagas cardiomyopathy. Expert. Rev. Cardiovasc. Ther. 2015, 13, 1393–1409. [Google Scholar] [CrossRef] [Green Version]
- Bern, C. Antitrypanosomal therapy for chronic Chagas’ disease. N. Engl. J. Med. 2011, 364, 2527–2534. [Google Scholar] [CrossRef]
- Jackson, Y.; Alirol, E.; Getaz, L.; Wolff, H.; Combescure, C.; Chappuis, F. Tolerance and safety of nifurtimox in patients with chronic chagas disease. Clin. Infect. Dis. 2010, 51, e69–e75. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, E.; Rojas, D.A.; Urbina, F.; Solari, A. The use of antioxidants as potential co-adjuvants to treat chronic Chagas disease. Antioxidants 2021, 10, 1022. [Google Scholar] [CrossRef]
- Cevey, Á.; Penas, F.N.; Alba Soto, C.D.; Mirkin, G.A.; Goren, N.B. IL-10/STAT3/SOCS3 axis is involved in the anti-inflammatory effect of benznidazole. Front. Immunol. 2019, 10, 1267. [Google Scholar] [CrossRef]
- Al-Kassas, R.; Bansal, M.; Shaw, J. Nanosizing techniques for improving bioavailability of drugs. J. Control Release 2017, 260, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Hasslocher-Moreno, A.M.; Saraiva, R.M.; Sangenis, L.H.C.; Xavier, S.S.; de Sousa, A.S.; Costa, A.R.; de Holanda, M.T.; Veloso, H.H.; Mendes, F.S.N.S.; Costa, F.A.C.; et al. Benznidazole decreases the risk of chronic Chagas disease progression and cardiovascular events: A long-term follow up study. EClinicalMedicine 2021, 31, 100694. [Google Scholar] [CrossRef]
- Brito, T.K.; Viana, R.L.S.; Moreno, C.J.G.; Barbosa, J.D.S.; Júnior, F.L.D.S.; de Medeiros, M.J.C.; Melo-Silveira, R.F.; Almeida-Lima, J.; Pontes, D.D.L.; Silva, M.S.; et al. Synthesis of silver nanoparticle employing corn cob xylan as a reducing agent with anti-Trypanosoma cruzi activity. Int. J. Nanomed. 2020, 15, 965–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasit. Vectors 2020, 13, 588. [Google Scholar] [CrossRef] [PubMed]
- Konstantinovic, N.; Guegan, H.; Stäjner, T.; Belaz, S.; Robert-Gangneux, F. Treatment of toxoplasmosis: Current options and future perspectives. Food Waterborne Parasitol. 2019, 15, e00036. [Google Scholar] [CrossRef]
- Machala, L.; Kodym, P.; Malý, M.; Geleneky, M.; Beran, O.; Jilich, D. Toxoplasmosis in immunocompromised patients. Epidemiol. Mikrobiol. Imunol. 2015, 64, 59–65. [Google Scholar]
- Abou-El-Naga, I.F.; El Kerdany, E.D.; Mady, R.F.; Shalaby, T.I.; Zaytoun, E.M. The effect of lopinavir/ritonavir and lopinavir/ritonavir loaded PLGA nanoparticles on experimental toxoplasmosis. Parasitol. Int. 2017, 66, 735–747. [Google Scholar] [CrossRef]
- Miller, C.M.; Boulter, N.R.; Ikin, R.J.; Smith, N.C. The immunobiology of the innate response to Toxoplasma gondii. Int. J. Parasitol. 2009, 39, 23–39. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, Y.M.; Liu, M.; Lu, Y.Q.; Liu, X.Y.; Zhang, Y.L.; Jiang, Z.S.; Yang, T.T.; Sun, Y.; Lan, K.; et al. Development of a risk scoring system for prognostication in HIV-related toxoplasma encephalitis. BMC Infect. Dis. 2020, 20, 923. [Google Scholar] [CrossRef]
- da Cruz, A.B.; Maia, M.M.; Pereira, I.S.; Taniwaki, N.N.; Namiyama, G.M.; Telles, J.P.M.; Vidal, J.E.; Spegiorin, L.C.J.F.; Brandão de Mattos, C.C.; Mattos, L.C.; et al. Human extracellular vesicles and correlation with two clinical forms of toxoplasmosis. PLoS ONE 2020, 15, e0229602. [Google Scholar] [CrossRef] [Green Version]
- Daher, D.; Shaghlil, A.; Sobh, E.; Hamie, M.; Hassan, M.E.; Moumneh, M.B.; Itani, S.; El Hajj, R.; Tawk, L.; El Sabban, M.; et al. Comprehensive overview of Toxoplasma gondii—Induced and associated diseases. Pathogens 2021, 10, 1351. [Google Scholar] [CrossRef]
- Zhang, K.; Lin, G.; Han, Y.; Li, J. Serological diagnosis of toxoplasmosis and standardization. Clin. Chim. Acta 2016, 461, 83–89. [Google Scholar] [CrossRef]
- Ybañez, R.H.D.; Ybañez, A.P.; Nishikawa, Y. Review on the current trends of toxoplasmosis serodiagnosis in humans. Front. Cell. Infect. Microbiol. 2020, 10, 204. [Google Scholar] [CrossRef]
- Smit, G.S.A.; Vu, B.T.L.; Do, D.T.; Do, Q.H.; Pham, H.Q.; Speybroeck, N.; Devleesschauwer, B.; Padalko, E.; Roets, E.; Dorny, P. Sero-epidemiological status and risk factors of toxoplasmosis in pregnant women in Northern Vietnam. BMC Infect. Dis. 2019, 19, 329. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, S.; Farid, A.; Rabae, I.; El-Amir, A. Novel IMB-ELISA assay for rapid diagnosis of human toxoplasmosis using SAG1 antigen. Jpn. J. Infect. Dis. 2015, 68, 474–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, Q.; Hou, P.; Chen, M.; Hui, W.; Vermorken, A.; Luo, Z.; Li, H.; Li, Q.; Cui, Y. Gold magnetic nanoparticle conjugate-based lateral flow assay for the detection of IgM class antibodies related to TORCH infections. Int. J. Mol. Med. 2015, 36, 1319–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assolini, J.P.; Concato, V.M.; Goncalves, M.D.; Carloto, A.C.M.; Conchon-Costa, I.; Pavanelli, W.R.; Melanda, F.N.; Costa, I.N. Nanomedicine advances in toxoplasmosis: Diagnostic, treatment, and vaccine applications. Parasitol. Res. 2017, 116, 1603–1615. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Liu, Y.; Chen, Y.; Yang, Q.; Chun, P.; Yao, K.; Han, X.; Wang, S.; Yu, S.; Wang, Q. A novel dynamic flow immunochromatographic test (DFICT) using gold nanoparticles for the serological detection of Toxoplasma gondii infection in dogs and cats. Biosens. Bioelectron. 2015, 72, 133–139. [Google Scholar] [CrossRef]
- Medawar-Aguilar, V.; Jofre, C.F.; Fernández-Baldo, M.A.; Alonso, A.; Angel, S.; Raba, J.; Pereira, S.V.; Messina, G.A. Serological diagnosis of Toxoplasmosis disease using a fluorescent immunosensor with chitosan-ZnO-nanoparticles. Anal. Biochem. 2019, 564–565, 116–122. [Google Scholar] [CrossRef]
- Aly, I.; Taher, E.E.; El Nain, G.; El Sayed, H.; Mohammed, F.A.; Hamad, R.S.; Bayoumy, E.M. Advantages of bioconjugated silica-coated nanoparticles as an innovative diagnosis for human toxoplasmosis. Acta Trop. 2018, 177, 19–24. [Google Scholar] [CrossRef]
- Sousa, S.; Castro, A.; Correia da Costa, J.M.; Pereira, E. Biosensor based immunoassay: A new approach for serotyping of Toxoplasma gondii. Nanomaterials 2021, 11, 2065. [Google Scholar] [CrossRef]
- Mammari, N.; Halabi, M.A.; Yaacoub, S.; Chlala, H.; Dardé, M.L.; Courtioux, B. Modulates the host cell responses: An overview of apoptosis pathways. Biomed. Res. Int. 2019, 2019, 6152489. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Singla, L.D.; Zhou, H. Vaccines against Toxoplasma gondii: Status, challenges and future directions. Hum. Vaccin. Immunother. 2012, 8, 1305–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongert, E.; Roberts, C.W.; Gargano, N.; Förster-Waldl, E.; Förster-Wald, E.; Petersen, E. Vaccines against Toxoplasma gondii: Challenges and opportunities. Mem. Inst. Oswaldo Cruz 2009, 104, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Chu, K.B.; Quan, F.S. Advances in Toxoplasma gondii vaccines: Current strategies and challenges for vaccine development. Vaccines 2021, 9, 413. [Google Scholar] [CrossRef]
- Yu, Z.; He, K.; Cao, W.; Aleem, M.T.; Yan, R.; Xu, L.; Song, X.; Li, X. Nano vaccines for T. gondii ribosomal P2 protein with nanomaterials as a promising DNA vaccine against toxoplasmosis. Front. Immunol. 2022, 13, 839489. [Google Scholar] [CrossRef]
- Chahal, J.S.; Khan, O.F.; Cooper, C.L.; McPartlan, J.S.; Tsosie, J.K.; Tilley, L.D.; Sidik, S.M.; Lourido, S.; Langer, R.; Bavari, S.; et al. Dendrimer-RNA nanoparticles generate protective immunity against lethal Ebola, H1N1 influenza, and Toxoplasma gondii challenges with a single dose. Proc. Natl. Acad. Sci. USA 2016, 113, E4133–E4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Kato, K. Inorganic nanoparticles kill Toxoplasma gondii via changes in redox status and mitochondrial membrane potential. Int. J. Nanomed. 2017, 12, 1647–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teimouri, A.; Azami, S.J.; Keshavarz, H.; Esmaeili, F.; Alimi, R.; Mavi, S.A.; Shojaee, S. Anti-Toxoplasma activity of various molecular weights and concentrations of chitosan nanoparticles on tachyzoites of RH strain. Int. J. Nanomed. 2018, 13, 1341–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etewa, S.E.; El-Maaty, D.A.A.; Hamza, R.S.; Metwaly, A.S.; Sarhan, M.H.; Abdel-Rahman, S.A.; Fathy, G.M.; El-Shafey, M.A. Assessment of spiramycin-loaded chitosan nanoparticles treatment on acute and chronic toxoplasmosis in mice. J. Parasit. Dis. 2018, 42, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Tessarolo, L.D.; de Menezes, R.R.P.P.; Mello, C.P.; Lima, D.B.; Magalhães, E.P.; Bezerra, E.M.; Sales, F.A.M.; Barroso Neto, I.L.; Oliveira, M.F.; Dos Santos, R.P.; et al. Nanoencapsulation of benznidazole in calcium carbonate increases its selectivity to Trypanosoma cruzi. Parasitology 2018, 145, 1191–1198. [Google Scholar] [CrossRef]
- Rahimi, M.T.; Ahmadpour, E.; Rahimi Esboei, B.; Spotin, A.; Kohansal Koshki, M.H.; Alizadeh, A.; Honary, S.; Barabadi, H.; Ali Mohammadi, M. Scolicidal activity of biosynthesized silver nanoparticles against Echinococcus granulosus protoscolices. Int. J. Surg. 2015, 19, 128–133. [Google Scholar] [CrossRef]
- Benelli, G. Gold nanoparticles—Against parasites and insect vectors. Acta Trop. 2018, 178, 73–80. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Whiteley, C.G. Interaction of metal nanoparticles with recombinant arginine kinase from Trypanosoma brucei: Thermodynamic and spectrofluorimetric evaluation. Biochim. Biophys. Acta 2014, 1840, 701–706. [Google Scholar] [CrossRef] [Green Version]
- Tomiotto-Pellissier, F.; Miranda-Sapla, M.M.; Machado, L.F.; Bortoleti, B.T.D.S.; Sahd, C.S.; Chagas, A.F.; Assolini, J.P.; Oliveira, F.J.A.; Pavanelli, W.R.; Conchon-Costa, I.; et al. Nanotechnology as a potential therapeutic alternative for schistosomiasis. Acta Trop. 2017, 174, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Gupta, S.S. Anti-leishmanial nanotherapeutics: A current perspective. Curr. Drug Metab. 2019, 20, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Diouani, M.F.; Ouerghi, O.; Belgacem, K.; Sayhi, M.; Ionescu, R.; Laouini, D. Casein-conjugated gold nanoparticles for amperometric detection of Leishmania infantum. Biosensors 2019, 9, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dykman, L.A. Gold nanoparticles for preparation of antibodies and vaccines against infectious diseases. Expert Rev. Vaccines 2020, 19, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Sutthanont, N.; Attrapadung, S.; Nuchprayoon, S. Larvicidal activity of synthesized silver nanoparticles from Curcuma zedoaria essential oil against Culex quinquefasciatus. Insects 2019, 10, 27. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Han, Y.; Kato, K. Modulation of host HIF-1α activity and the tryptophan pathway contributes to the anti-Toxoplasma gondii potential of nanoparticles. Biochem. Biophys. Rep. 2017, 11, 84–92. [Google Scholar] [CrossRef]
- Gaafar, M.R.; Mady, R.F.; Diab, R.G.; Shalaby, T.I. Chitosan and silver nanoparticles: Promising anti-toxoplasma agents. Exp. Parasitol. 2014, 143, 30–38. [Google Scholar] [CrossRef]
- Dunay, I.R.; Heimesaat, M.M.; Bushrab, F.N.; Müller, R.H.; Stocker, H.; Arasteh, K.; Kurowski, M.; Fitzner, R.; Borner, K.; Liesenfeld, O. Atovaquone maintenance therapy prevents reactivation of toxoplasmic encephalitis in a murine model of reactivated toxoplasmosis. Antimicrob. Agents Chemother. 2004, 48, 4848–4854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shubar, H.M.; Lachenmaier, S.; Heimesaat, M.M.; Lohman, U.; Mauludin, R.; Mueller, R.H.; Fitzner, R.; Borner, K.; Liesenfeld, O. SDS-coated atovaquone nanosuspensions show improved therapeutic efficacy against experimental acquired and reactivated toxoplasmosis by improving passage of gastrointestinal and blood-brain barriers. J. Drug Target. 2011, 19, 114–124. [Google Scholar] [CrossRef]
- Sordet, F.; Aumjaud, Y.; Fessi, H.; Derouin, F. Assessment of the activity of atovaquone-loaded nanocapsules in the treatment of acute and chronic murine toxoplasmosis. Parasite 1998, 5, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaguarnera, L. Influence of resveratrol on the immune response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [Green Version]
- Polat, Z.A.; Cetin, A.; Savage, P.B. Evaluation of the in vitro activity of ceragenins against Trichomonas vaginalis. Acta Parasitol. 2016, 61, 376–381. [Google Scholar] [CrossRef]
- Azami, S.J.; Teimouri, A.; Keshavarz, H.; Amani, A.; Esmaeili, F.; Hasanpour, H.; Elikaee, S.; Salehiniya, H.; Shojaee, S. Curcumin nanoemulsion as a novel chemical for the treatment of acute and chronic toxoplasmosis in mice. Int. J. Nanomed. 2018, 13, 7363–7374. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Lei, C.; Li, J.; Wu, Z.; Shen, G.; Yu, R. A piezoelectric immunoagglutination assay for Toxoplasma gondii antibodies using gold nanoparticles. Biosens. Bioelectron. 2004, 19, 701–709. [Google Scholar] [CrossRef]
- Wang, H.; Li, Q.; Reyes, S.; Zhang, J.; Zeng, Q.; Zhang, P.; Xie, L.; Lee, P.J.; Roncal, N.; Melendez, V.; et al. Nanoparticle formulations of decoquinate increase antimalarial efficacy against liver stage Plasmodium infections in mice. Nanomedicine 2014, 10, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Pissinate, K.; dos Santos Martins-Duarte, É.; Schaffazick, S.R.; de Oliveira, C.P.; Vommaro, R.C.; Guterres, S.S.; Pohlmann, A.R.; de Souza, W. Pyrimethamine-loaded lipid-core nanocapsules to improve drug efficacy for the treatment of toxoplasmosis. Parasitol. Res. 2014, 113, 555–564. [Google Scholar] [CrossRef]
- Tran, T.M.; Crompton, P.D. Decoding the complexities of human malaria through systems immunology. Immunol. Rev. 2020, 293, 144–162. [Google Scholar] [CrossRef] [PubMed]
- Castro-Sesquen, Y.E.; Kim, C.; Gilman, R.H.; Sullivan, D.J.; Searson, P.C. Nanoparticle-based histidine-rich protein-2 assay for the detection of the malaria parasite Plasmodium falciparum. Am. J. Trop. Med. Hyg. 2016, 95, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A. Role of the European pharmacist in the implementation of the latest WHO guidelines for malaria. Pathogens 2023, 12, 729. [Google Scholar] [CrossRef]
- Rahman, K.; Khan, S.U.; Fahad, S.; Chang, M.X.; Abbas, A.; Khan, W.U.; Rahman, L.; Haq, Z.U.; Nabi, G.; Khan, D. Nano-biotechnology: A new approach to treat and prevent malaria. Int. J. Nanomed. 2019, 14, 1401–1410. [Google Scholar] [CrossRef] [Green Version]
- Ponarulselvam, S.; Panneerselvam, C.; Murugan, K.; Aarthi, N.; Kalimuthu, K.; Thangamani, S. Synthesis of silver nanoparticles using leaves of Catharanthus roseus Linn. G. Don and their antiplasmodial activities. Asian Pac. J. Trop. Biomed. 2012, 2, 574–580. [Google Scholar] [CrossRef] [Green Version]
- Kojom Foko, L.P.; Eya’ane Meva, F.; Eboumbou Moukoko, C.E.; Ntoumba, A.A.; Ngaha Njila, M.I.; Belle Ebanda Kedi, P.; Ayong, L.; Lehman, L.G. A systematic review on anti-malarial drug discovery and antiplasmodial potential of green synthesis mediated metal nanoparticles: Overview, challenges and future perspectives. Malar. J. 2019, 18, 337. [Google Scholar] [CrossRef] [Green Version]
- Maizels, R.M.; McSorley, H.J. Regulation of the host immune system by helminth parasites. J. Allergy Clin. Immunol. 2016, 138, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Mukai, K.; Tsai, M.; Starkl, P.; Marichal, T.; Galli, S.J. IgE and mast cells in host defense against parasites and venoms. Semin. Immunopathol. 2016, 38, 581–603. [Google Scholar] [CrossRef] [Green Version]
- Tedla, M.G.; Every, A.L.; Scheerlinck, J.Y. Investigating immune responses to parasites using transgenesis. Parasit. Vectors 2019, 12, 303. [Google Scholar] [CrossRef] [Green Version]
- Obata-Ninomiya, K.; Domeier, P.P.; Ziegler, S.F. Basophils and eosinophils in nematode infections. Front. Immunol. 2020, 11, 583824. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.E.; Sutherland, T.E. Host protective roles of type 2 immunity: Parasite killing and tissue repair, flip sides of the same coin. Semin. Immunol. 2014, 26, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.M.; Smith, N.C.; Ikin, R.J.; Boulter, N.R.; Dalton, J.P.; Donnelly, S. Immunological interactions between 2 common pathogens, Th1-inducing protozoan Toxoplasma gondii and the Th2-inducing helminth Fasciola hepatica. PLoS ONE 2009, 4, e5692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coban, C.; Lee, M.S.J.; Ishii, K.J. Tissue-specific immunopathology during malaria infection. Nat. Rev. Immunol. 2018, 18, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Haldar, K.; Mohandas, N. Malaria, erythrocytic infection, and anemia. Hematol. Am. Soc. Hematol. Educ. Program 2009, 2009, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Najer, A.; Wu, D.; Bieri, A.; Brand, F.; Palivan, C.G.; Beck, H.P.; Meier, W. Nanomimics of host cell membranes block invasion and expose invasive malaria parasites. ACS Nano 2014, 8, 12560–12571. [Google Scholar] [CrossRef]
- Jewett, T.J.; Sibley, L.D. The toxoplasma proteins MIC2 and M2AP form a hexameric complex necessary for intracellular survival. J. Biol. Chem. 2004, 279, 9362–9369. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.M.; Huynh, M.H.; Coppens, I.; Parussini, F.; Moreno, S.; Carruthers, V.B. A cleavable propeptide influences Toxoplasma infection by facilitating the trafficking and secretion of the TgMIC2-M2AP invasion complex. Mol. Biol. Cell 2006, 17, 4551–4563. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
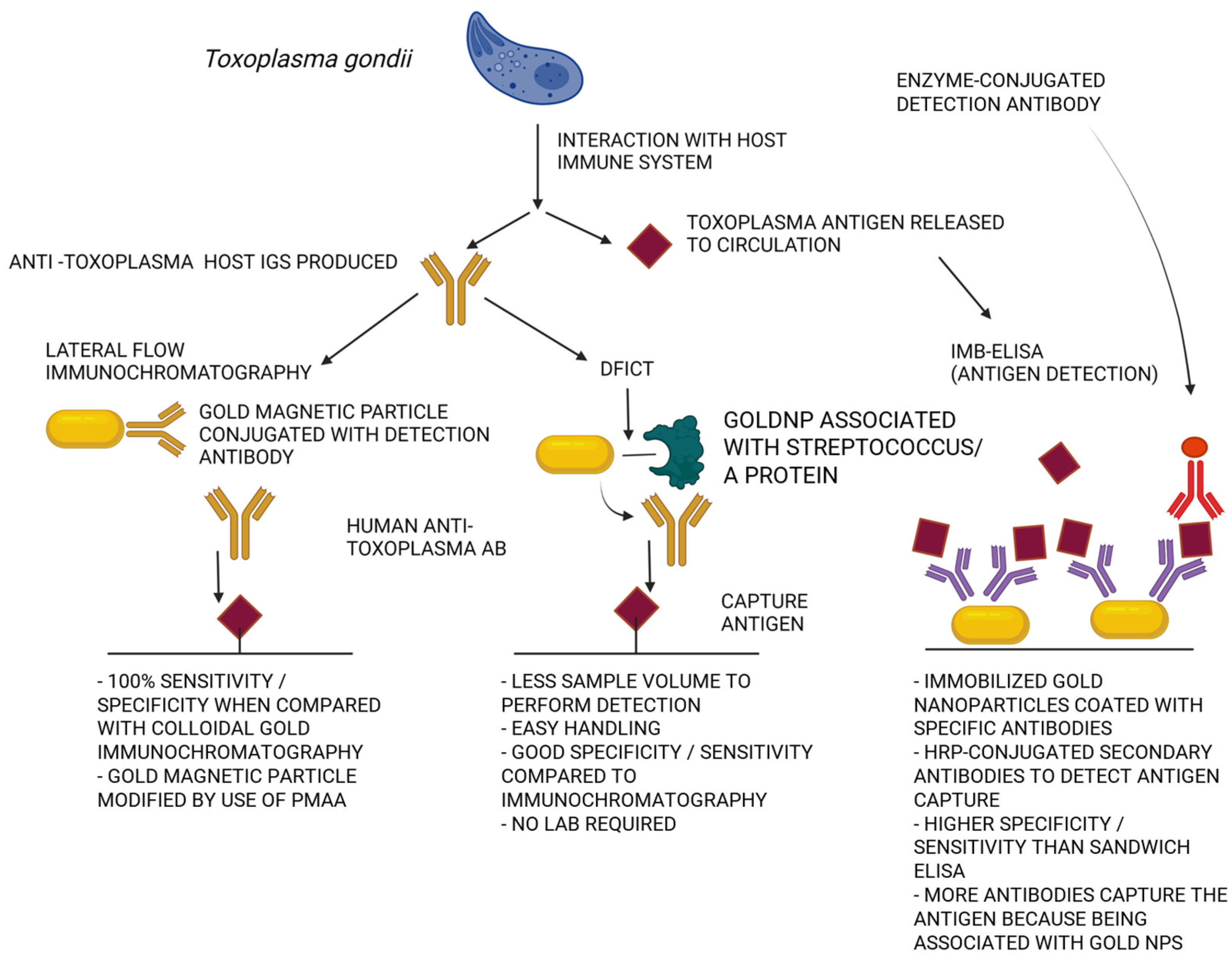{kind=link}
| Group of Antiparasitic Drugs | Antiparasitic Drugs |
|---|---|
| Anti-malarials | artemisinin chloroquine, amodiaquine, atovaquone, primaquine, mefloquine, atovaquone-proguanil, doxycycline |
| Anti-babesial agents | atovaquone, azithromycin, clindamycin, quinine |
| Anti-amoebic agents | iodoquinol, paromomycin sulfate, diloxanide furoate, metronidazole, tinidazole, emetine, chloroquine |
| Anti-giardial agents | metronidazol, tinidazole, furazolidone, albendazole |
| Trypanocidal agents | nifurtimox, benznidazole, pentamidine, eflornithine, suramin, melarsoprol |
| Anti-leishmanial agents | meglumine antimoniate, pentamidine, amphotericin B |
| Anti-toxoplasma agents | sulfadiazine, pyrimethamine, sulfamethazine, sulfamerazine |
| Anti-cestodal drugs | praziquantel, niclosamide, albendazole |
| Anti-nematodal drugs | praziquantel, metrifonate, oxamniquine Albendazole, diethylcarbamazine, ivermectin |
| Species of the Parasite | Nanoparticles | Therapeutic Result | Bibliography |
|---|---|---|---|
| Trypanosoma brucei | AgNP, AuNP, PtNP | strong anti-parasitic effect | [77] |
| Leishmania tropica | ZnONP | effect on the cell membrane of the parasite | [78] |
| Leishmania major | ZnONP | induction of apoptosis, toxic effect on the promastigote of L. major | [79] |
| AgNP, AuNP, TiO2NP, ZnONPiMgONP | high antiparasitic activity | [80,81,82,83,84] | |
| Leishmania donovanii | ZnONP | cytotoxic effect on the amastigote | [85] |
| Schistosma mansoni | AuNP | antihelmintic action | [63,86,87] |
| carbon nanoparticles with gold and silver, magnetic nanoparticles, carbon nanotubes, colloidal gold | high anti-parasitic activity | [86,88,89] | |
| Acanthamoeba keratitis | AgNP | anti-amoebic effect on Acanthamoeba trophozoites | [90,91,92,93,94,95] |
| Plasmodium falciparum | metal oxide nanoparticles Fe3O4, MgO, ZrO2, Al2O3, CeO2, AuNP | demonstrated anti-plasmoid activity | [65,91] |
| Toxoplasma gondii | AuNP AgNP | high anti-parasitic activity | [96,97] |
| Nanoparticles and Nanosystems Used in Management of Toxoplasmosis | |
|---|---|
| in Diagnostics: | in Therapy: |
| Nanoparticles of zinc oxide (ZnO-NP) covered with chitosan [149] | Chitosan and silver nanoparticles [171] |
| Silicon coated nanoparticles (SiO2NP) [150] | Curcumin nanoemulsion [177] |
| Gold nanoparticles [178,179] | Gold and iron nanoparticles [33] Lipid-core nanocapsules with pyrimethamine [180] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Król, G.; Fortunka, K.; Majchrzak, M.; Piktel, E.; Paprocka, P.; Mańkowska, A.; Lesiak, A.; Karasiński, M.; Strzelecka, A.; Durnaś, B.; et al. Metallic Nanoparticles and Core-Shell Nanosystems in the Treatment, Diagnosis, and Prevention of Parasitic Diseases. Pathogens 2023, 12, 838. https://doi.org/10.3390/pathogens12060838
Król G, Fortunka K, Majchrzak M, Piktel E, Paprocka P, Mańkowska A, Lesiak A, Karasiński M, Strzelecka A, Durnaś B, et al. Metallic Nanoparticles and Core-Shell Nanosystems in the Treatment, Diagnosis, and Prevention of Parasitic Diseases. Pathogens. 2023; 12(6):838. https://doi.org/10.3390/pathogens12060838
Chicago/Turabian StyleKról, Grzegorz, Kamila Fortunka, Michał Majchrzak, Ewelina Piktel, Paulina Paprocka, Angelika Mańkowska, Agata Lesiak, Maciej Karasiński, Agnieszka Strzelecka, Bonita Durnaś, and et al. 2023. "Metallic Nanoparticles and Core-Shell Nanosystems in the Treatment, Diagnosis, and Prevention of Parasitic Diseases" Pathogens 12, no. 6: 838. https://doi.org/10.3390/pathogens12060838