
Waterborne Pathogens: Detection Methods and Challenges


Abstract
:1. Introduction
2. Emerging Waterborne Pathogens and Related Diseases
3. Detection Methods for Waterborne Pathogens

{kind=link}
{kind=link}
{kind=link}
| Pathogen | Associated disease b | Health significance c | Persistence in drinking water supplies d |
|---|---|---|---|
| Bacteria | |||
| Campylobacter spp., C. jejuni | Diarrhea, gastroenteritis | High | Moderate |
| Yersinia enterocolitica | Diarrhea, reactive arthritis | High | Long |
| Escherichia coli, particularly enterohemorragic E. coli (EHEC), and other such as enteropathogenic (EPEC), enterotoxigenic (ETEC), and enteroinvasive (EIEC) | Acute diarrhea, bloody diarrhea and gastroenteritis | High | Moderate |
| Burkholderia pseudomallei | Meliodosis | High | May multiply |
| Legionella pneumophila and related bacteria | Acute respiratory illness, pneumonia (legionellosis) | High | May multiply |
| Non-tuberculous mycobacteria | Pulmonary disease, skin infection | Low | May multiply |
| Pseudomonas aeruginosa | Infections on lungs, urinary tract, and kidney Can cause inflammation and sepsis | Moderate | May multiply |
| Salmonella enterica serotype Typhi | Typhoid fever, paratyphoid fever and other serious salmonellosis | High | Moderate |
| Other salmonellae | Gastroenteritis, reactive arthritis | High | May multiply |
| Shigella spp. | Bacillary dysentery or shigellosis | High | Short |
| Vibrio cholerae | Gastroenteritis, cholera | High | Short to long |
| Helicobacter pylori | Chronic gastritis, ulcer disease and gastric cancer | Low | Moderate |
| Virus | |||
| Adenovirus | Gastroenteritis | High | Long |
| Enteroviruses | Gastroenteritis | High | Long |
| Hepatitis A virus | Hepatitis | High | Long |
| Hepatitis E virus | Infectious hepatitis; miscarriage and death | High | Long |
| Rotavirus | Gastroenteritis | High | Long |
| Sapoviruses | Acute viral gastroenteritis | High | Long |
| Astroviruses | Diarrhea | High | Long |
| Norovirus | Gastroenteritis | High | Long |
| Protozoa | |||
| Acanthamoeba spp. | Amoebic meningoencephalitis, keratitis, encephalitis | High | May multiply |
| Cryptosporidium parvum | Cryptosporidiosis, diarrhea | High | Long |
| Cryptosporidium cayetanensis | Diarrhea | High | Long |
| Entamoeba histolytica | Amoeba dysentery | High | Moderate |
| Giardia intestinalis | Diarrhea | High | Moderate |
| Naegleria fowleri | Infection of the brain called primary amebic meningoencephalitis (PAM) | High | May multiply in warm water |
| Toxoplasma gondii | Toxoplasmosis, miscarriage, birth defects | High | Long |
| Helminths | |||
| Dracunculus medinensis | Dracunculiasis (Guinea worm disease, ulcerating skin infection). | High | Moderate |
| Schistosoma spp. | Schistosomiasis, , liver and kidney damage, itchy skin, fever, chills, cough and muscle aches. | High | Short |

| Detection Method | Water Pathogens | Detection Limit (LOD) | Matrix Sample | References |
|---|---|---|---|---|
| PCR | E. coli | 1 cfu /100 mL | Contaminated tap water. | [34] |
| Enterotoxigenic E. coli (ETEC) | 4 cfu/mL | Water samples spiked by ETEC and nonpathogenic E. coli. | [35] | |
| Cryptosporidium and Giardia | 1 to 10 oocyst and 5–50 cysts | Environmental water samples. | [36] | |
| Multiplex PCR | EHEC, Shigella sp., Vibrio parahaemolyticus, P. aeruginosa and Salmonella sp. | 101 cfu, 102 cfu, 102 cfu, 102 cfu and 101 cfu | Polluted water and natural water. | [37] |
| Quantitative PCR (qPCR) | Adenovirus (adenovirus fiber gene in AdV40 and AdV41) | 5–8 copies of AdV40/41 | Wastewater, drinking water, recreational waters, and rivers. | [38] |
| L. monocytogenes, V. cholerae, V. parahaemolyticus, Pseudogulbenkiana sp., S. typhimurium, S. flexneri, C. perfringens and pathogenic E. coli (Microfluidic qPCR) | From 102 to 104 cells per ca. 200 mg fecal samples of pathogens 100 cells/L | Spiked environmental water samples (pond) and natural fresh water lake. | [39] | |
| Adenovirus, Aichi virus, astrovirus, enterovirus, human norovirus, rotavirus, sapovirus, and hepatitis A and E viruses (Microfluidic qPCR) | 2 copies/μL of cDNA/DNA | River water contaminated with effluents from a wastewater treatment plant. | [40] | |
| Real-time PCR | V. cholerae | 1 cfu/100 mL | Ballast water. | [41] |
| C. hominis, C. parvum, C. meleagridis, C. wrairi | 1 oocyst | Sewage and river water. | [42,43] | |
| Astrovirus | 5–7 GC logs /100 mL | Sewage. | [44] | |
| L. monocytogenes on biofilms | 6 × 102 cfu/cm2 | Artificial biofilms. | [45] | |
| Microarrays | Bacillus anthracis, Brucella abortus, C. botulinum, Coxiella burnetii, Francisella tularensi, Rickettsia prowazekii, C. perfringens, S. aureus, V. cholerae, V. alginolyticus, Yersinia pestis Western equine encephalitis, Eastern equine encephalitis, Ebola, Venezuelan equine encephalitis virus, Alexandrium cantenella, Fusarium sporotrichioides | 10 fg of B. anthracis DNA, 500 fg of F. tularensis DNA and Y. pestis DNA | Environmental water and ocean water spiked with pathogen. | [46] |
| Y. enterocolitica, E. coli, S.enterica Typhimurium, | 1 × 107 S. typhimurium cells (104 S. enterica genomes) | Wastewater. | [47] | |
| Cryptosporidium, Acanthamoeba spp., Blastocystis hominis, Entamoeba spp., Giardia intestinalis, Naegleria spp. | 1 × 103 target genes, or 50 Cryptosporidium parvum oocysts, per assay | Municipal wastewater treatment plants. | [48] | |
| Microarray | E. coli, A. hydrophila (DNA microarray and real-time qPCR techniques) | 103 A. hydrophila cells per sample, 5 ng of E. coli and 10 ng of A. hydrophila | Municipal wastewater treatment plant in each stage of the disinfection process. | [49] |
| C. parvum, C. hominis, E. faecium, B. anthracis, F. tularensis | 100 ng of DNA, Microarray assay: 20 genomic copies without a PCR pre-amplification step | Tap water spiked with multiple organisms. | [50] | |
| Pyrosequencing | Y. pestis | 0.9 cfu/mL | Milk. | [51] |
| B. anthracis (Immunomagnetic separation and pyrosequencing) | 6 cfu/mL | Bottled water, milk and juice. | [52] | |
| Comamonadaceae, Proteobacteria, Bacteroidetes, Planctomycetes, and Elusimicrobia | 1.3 ×105 cells/mL | Drinking water of the non-chlorinated distribution system. | [53] | |
| Biosensors | C. parvum | 105 oocysts/mL | Oocysts diluted in PBS. | [54] |
| E. coli 0157:H7 (NASBA +LFA biosensor) | 40 cells/mL | Drinking water. | [55] | |
| V. cholerae (Amperometric) | 8 cfu/mL | Ground and sea water. | [56] | |
| Microcystis spp. (Optical fibre) | 30 ng/L | Lake water. | [57,58] | |
| Fluorescence in situ hybridization (FISH) | F. psychrophilum | 7.3 × 105 cells/mL | Culture suspension. | [59] |
| E. coli (catalyzed reporter deposition (CARD)-FISH) | 8.9 ±1.5 16S rRNA molecules per cell | Mixed pure cultures and sludge. | [60] | |
| E. coli | 1,400 ± 170 16S rRNA copies per E. coli | Activated sludge. | [60] | |
| Immunology-based methods | E. coli O157:H7 and S. enterica typhimurium. | 1.8 × 103 cfu/mL of E. coli and 9.2 × 103 cfu/mL of Salmonella | Contaminated food. | [61,62] |
3.1. Polymerase Chain Reaction (PCR)
3.2. Oligonucleotide DNA Microarrays
3.3. Pyrosequencing
3.4. Fluorescence in Situ Hybridization (FISH)

3.5. Immunology-Based Methods
3.6. Biosensor Based Methods
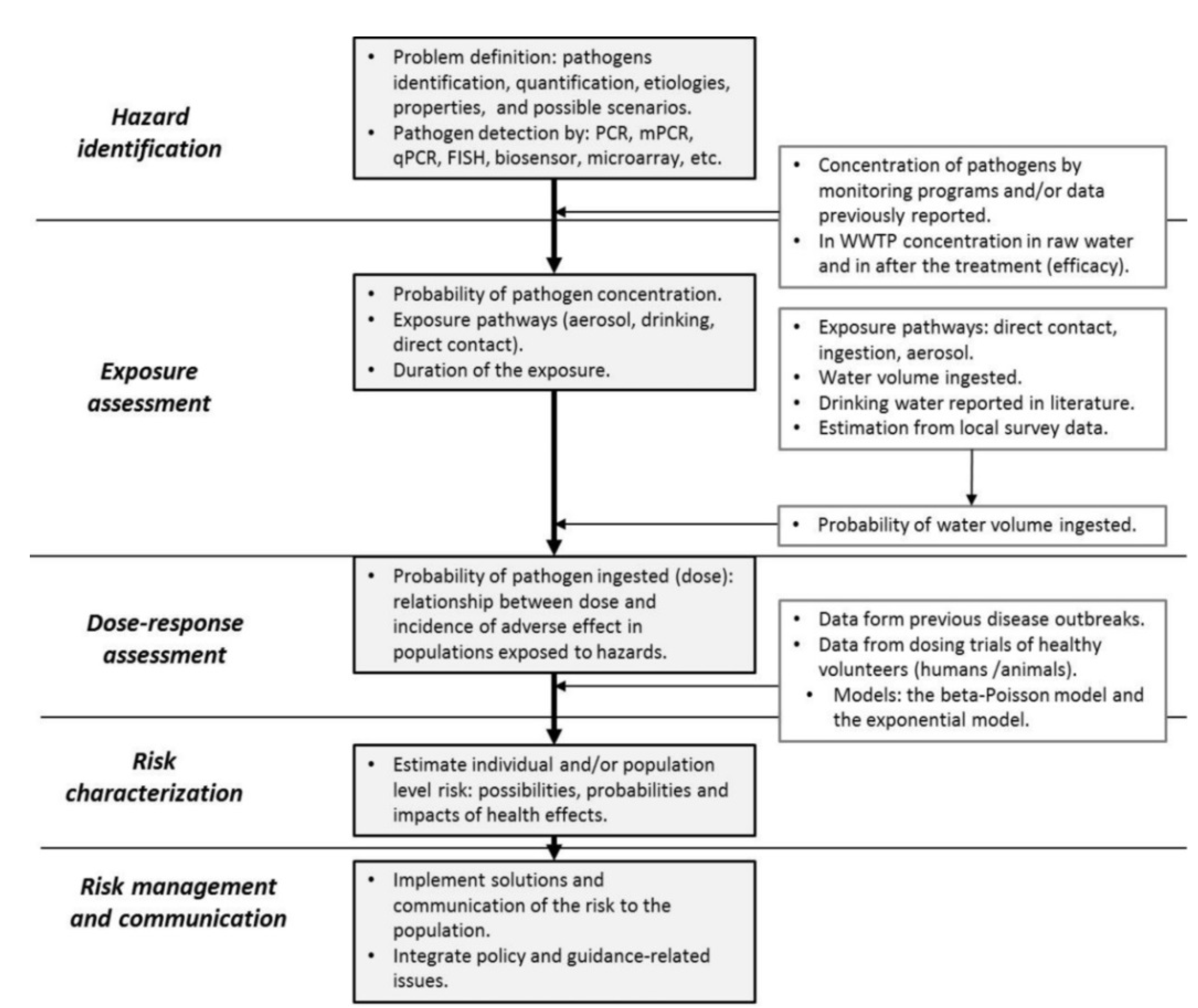
4. Quantitative Microbial Risk Assessment of Waterborne Disease

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization (WHO). Water Sanitation and Health. 2015. Available online: http://www.who.int/water_sanitation_health/diseases (assessed on 17 February 2015).
- Bitton, G. Microbiology of Drinking Water Production and Distribution, 1st ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; p. 312. [Google Scholar]
- Ingerson-Mahar, M.; Reid, A. Microbes in Pipes: The Microbiology of the Water Distribution System A Report on an American Academy of Microbiology Colloquium; ASM Academy: Boulder, CO, USA, 2012; p. 26. [Google Scholar]
- Alhamlan, F.S.; Al-Qahtani, A.A.; Al-Ahdal, M.N. Recommended advanced techniques for waterborne pathogen detection in developing countries. J. Infect. Dev. Ctries 2015, 9, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, H.; Schwartzbrod, L.; Dei-Cas, E. Microbial agents associated with waterborne diseases. Crit. Rev. Microbiol. 2002, 28, 371–409. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Global Water, Sanitation, and Hygiene (WASH). Available online: http://www.cdc.gov/healthywater/global/ (assesed on 17 February 2015).
- World Health Organization (WHO). Progress on Sanitation and Drinking-Water: 2010 Update Report; World Health Organization (WHO): Geneva, Switzerland, 2010. [Google Scholar]
- Pandey, P.K.; Kass, P.H.; Soupir, M.L.; Biswas, S.; Singh, V.P. Contamination of water resources by pathogenic bacteria. AMB Express 2014, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Millennium Development Goals: Progress towards the Health-Related Millennium Development Goals; World Health Organization (WHO): Geneva, Switzerland, 2010. Available online: http://www.who.int/topics/millennium_development_goals/en/ (assessed on 17 February 2015).
- Craun, M.F.; Craun, G.F.; Calderon, R.L.; Beach, M.J. Waterborne outbreaks reported in the United States. J. Water Health 2006, 4, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Craun, G.F.; Brunkard, J.M.; Yoder, J.S.; Roberts, V.A.; Carpenter, J.; Wade, T.; Calderon, R.L.; Roberts, J.M.; Beach, M.J.; Roy, S.L. Causes of outbreaks associated with drinking water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010, 23, 507–528. [Google Scholar] [CrossRef] [PubMed]
- Hrudey, S.E.; Payment, P.; Huck, P.M.; Gillham, R.W.; Hrudey, E.J. A fatal waterborne disease epidemic in Walkerton, Ontario: Comparison with other waterborne outbreaks in the developed world. Water Sci. Technol. 2003, 47, 7–14. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention (CDC). Surveillance for waterborne disease outbreaks associated with drinking water and other nonrecreational water—United States, 2009–2010; Centers for Disease Control and Prevention (CDC): Atlanta, GA, 2013; Volume 62, pp. 714–720. [Google Scholar]
- Lopez, A.L.; Macasaet, L.Y.; Ylade, M.; Tayag, E.A.; Ali, M. Epidemiology of cholera in the Philippines. PLoS Negl. Trop. Dis. 2015, 9, e3440. [Google Scholar] [CrossRef] [PubMed]
- Orata, F.D.; Keim, P.S.; Boucher, Y. The 2010 cholera outbreak in Haiti: How science solved a controversy. PLoS Pathog. 2014, 10, e1003967. [Google Scholar] [CrossRef] [PubMed]
- Tappero, J.W.; Tauxe, R.V. Lessons learned during public health response to cholera epidemic in Haiti and the Dominican Republic. Emerg. Infect. Dis. 2011, 17, 2087–2093. [Google Scholar] [CrossRef] [PubMed]
- Muniesa, M.; Hammerl, J.A.; Hertwig, S.; Appel, B.; Brussow, H. Shiga toxin-producing Escherichia coli O104:H4: A new challenge for microbiology. Appl. Environ. Microbiol. 2012, 78, 4065–4073. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disaese Provention and Control (ECDPC). Annual Epidemiological Report Food-and Waterborne Diseases and Zoonoses; European Centre for Disaese Provention and Control (ECDPC): Stockholm, Sweden, 2014. [Google Scholar]
- Hunter, P.R.; Chalmers, R.M.; Hughes, S.; Syed, Q. Self-reported diarrhea in a control group: A strong association with reporting of low-pressure events in tap water. Clin. Infect. Dis. 2005, 40, e32–e34. [Google Scholar] [CrossRef] [PubMed]
- Dore, M.H. Global Drinking Water Management and Conservation Optimal Decision-Making, 1st ed.; Springer International Publishing: Switzerland, 2015; p. 303. [Google Scholar]
- Blackburn, B.G.; Craun, G.F.; Yoder, J.S.; Hill, V.; Calderon, R.L.; Chen, N.; Lee, S.H.; Levy, D.A.; Beach, M.J. Surveillance for waterborne-disease outbreaks associated with drinking water--United States, 2001–2002. MMWR Surveill. Summ. 2004, 53, 23–45. [Google Scholar] [PubMed]
- Nwachcuku, N.; Gerba, C.P. Emerging waterborne pathogens: Can we kill them all? Curr. Opin. Biotechnol. 2004, 15, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J. Where do emerging pathogens come from? Microbe 2006, 1, 511–515. [Google Scholar]
- World Health Organization (WHO). Guidelines fror Drinking Water Quality–Volume 1: Recommendations; World Health Organization (WHO): Geneva, Switzwerland, 2008. Available online: http://www.who.int/water_sanitation_health/dwq/gdwq3rev/en/ (assessed on 27 April 2015).
- Straub, T.M.; Chandler, D.P. Towards a unified system for detecting waterborne pathogens. J. Microbiol. Methods 2003, 53, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Kostic, T.; Stessl, B.; Wagner, M.; Sessitsch, A. Microarray analysis reveals the actual specificity of enrichment media used for food safety assessment. J. Food Prot. 2011, 74, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lin, C.W.; Wang, J.; Oh, D.H. Advances in rapid detection methods for foodborne pathogens. J. Microbiol. Biotechnol. 2014, 24, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Law, J.W.; Ab Mutalib, N.S.; Chan, K.G.; Lee, L.H. Rapid methods for the detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2014, 5, 770. [Google Scholar] [PubMed]
- Cabral, J.P. Water microbiology. Bacterial pathogens and water. Int. J. Environ. Res. Public Health 2010, 7, 3657–3703. [Google Scholar] [CrossRef] [PubMed]
- Nygård, K. Doctoral. Water and infection: Epidemiological studies of epidemic and endemic Waterborne Disease. PhD thesis, University of Oslo, Oslo, Norway, 2008. [Google Scholar]
- McLellan, S.L.; Eren, A.M. Discovering new indicators of fecal pollution. Trends Microbiol. 2014, 22, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Gilbride, K. Molecular Methods for the Detection of Waterborne Pathogens. In Waterborne Pathogens, Detection Methods and Applications; Bridle, H., Ed.; Elsevier B.V.: London, UK, 2014; p. 387. [Google Scholar]
- Girones, R.; Ferrus, M.A.; Alonso, J.L.; Rodriguez-Manzano, J.; Calgua, B.; Correa Ade, A.; Hundesa, A.; Carratala, A.; Bofill-Mas, S. Molecular detection of pathogens in water--the pros and cons of molecular techniques. Water Res. 2010, 44, 4325–4339. [Google Scholar] [CrossRef] [PubMed]
- Tsen, H.Y.; Lin, C.K.; Chi, W.R. Development and use of 16s rRNA gene targeted PCR primers for the identification of Escherichia coli cells in waters. J. Appl. Microbiol. 1998, 85, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Vajpayee, P.; Shanker, R. Rapid culture-independent quantitative detection of enterotoxigenic Escherichia coli in surface waters by real-time PCR with molecular beacon. Environ. Sci. Technol. 2008, 42, 4577–4582. [Google Scholar] [CrossRef] [PubMed]
- Rochelle, P.A.; Ferguson, D.M.; Handojo, T.J.; de Leon, R.; Stewart, M.H.; Wolfe, R.L. An assay combining cell culture with reverse transcriptase PCR to detect and determine the infectivity of waterborne Cryptosporidium parvum. Appl. Environ. Microbiol. 1997, 63, 2029–2037. [Google Scholar] [PubMed]
- Fan, H.; Wu, Q.; Kou, X. Co-detection of five species of water-borne bacteria by multiplex PCR. Life Sci. J. 2008, 5, 47–54. [Google Scholar]
- Jothikumar, N.; Cromeans, T.L.; Hill, V.R.; Lu, X.; Sobsey, M.D.; Erdman, D.D. Quantitative real-time PCR assays for detection of human adenoviruses and identification of serotypes 40 and 41. Appl. Environ. Microbiol. 2005, 71, 3131–3136. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Nakamura, T.; Ozawa, S.; Kobayashi, A.; Sano, D.; Okabe, S. Water quality monitoring and risk assessment by simultaneous multipathogen quantification. Environ. Sci. Technol. 2014, 48, 4744–4749. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Kitamura, G.; Segawa, T.; Kobayashi, A.; Miura, T.; Sano, D.; Okabe, S. Microfluidic quantitative PCR for simultaneous quantification of multiple viruses in environmental water samples. Appl. Environ. Microbiol. 2014, 80, 7505–7511. [Google Scholar] [CrossRef] [PubMed]
- Fykse, E.M.; Nilsen, T.; Nielsen, A.D.; Tryland, I.; Delacroix, S.; Blatny, J.M. Real-time PCR and NASBA for rapid and sensitive detection of Vibrio cholerae in ballast water. Mar. Pollut. Bull. 2012, 64, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Masago, Y.; Oguma, K.; Katayama, H.; Ohgaki, S. Quantification and genotyping of Cryptosporidium spp. in river water by quenching probe PCR and denaturing gradient gel electrophoresis. Water Sci. Technol. 2006, 54, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Guy, R.A.; Payment, P.; Krull, U.J.; Horgen, P.A. Real-time PCR for quantification of Giardia and Cryptosporidium in environmental water samples and sewage. Appl. Environ. Microbiol. 2003, 69, 5178–5185. [Google Scholar] [CrossRef] [PubMed]
- Le Cann, P.; Ranarijaona, S.; Monpoeho, S.; Le Guyader, F.; Ferre, V. Quantification of human astroviruses in sewage using real-time RT-PCR. Res. Microbiol. 2004, 155, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Guilbaud, M.; de Coppet, P.; Bourion, F.; Rachman, C.; Prevost, H.; Dousset, X. Quantitative detection of Listeria monocytogenes in biofilms by real-time PCR. Appl. Environ. Microbiol. 2005, 71, 2190–2194. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.J.; Strout, C.L.; DeSantis, T.Z.; Stilwell, J.L.; Carrano, A.V.; Andersen, G.L. Sequence-specific identification of 18 pathogenic microorganisms using microarray technology. Mol. Cell Probes 2002, 16, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.; Berthiaume, F.; Lemarchand, K.; Harel, J.; Payment, P.; Bayardelle, P.; Masson, L.; Brousseau, R. Waterborne pathogen detection by use of oligonucleotide-based microarrays. Appl. Environ. Microbiol. 2005, 71, 8548–8557. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Seto, P.; Korczak, R. DNA microarray-based detection and identification of waterborne protozoan pathogens. J. Microbiol. Methods 2010, 80, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Lauder, H.; Cruwys, H.; Falletta, P.; Beaudette, L.A. Development and application of an oligonucleotide microarray and real-time quantitative PCR for detection of wastewater bacterial pathogens. Sci. Total Environ. 2008, 398, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, N.E.; Francisco, R.; Nichols, T.L.; Robinson, D.; Schaefer, F.W., 3rd; Schaudies, R.P.; Villegas, E.N. Detection of multiple waterborne pathogens using microsequencing arrays. J. Appl. Microbiol. 2013, 114, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Amoako, K.K.; Thomas, M.C.; Kong, F.; Janzen, T.W.; Hahn, K.R.; Shields, M.J.; Goji, N. Rapid detection and antimicrobial resistance gene profiling of Yersinia pestis using pyrosequencing technology. J. Microbiol. Methods 2012, 90, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Amoako, K.K.; Janzen, T.W.; Shields, M.J.; Hahn, K.R.; Thomas, M.C.; Goji, N. Rapid detection and identification of Bacillus anthracis in food using pyrosequencing technology. Int. J. Food Microbiol. 2013, 165, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Lautenschlager, K.; Hwang, C.; Liu, W.T.; Boon, N.; Koster, O.; Vrouwenvelder, H.; Egli, T.; Hammes, F. A microbiology-based multi-parametric approach towards assessing biological stability in drinking water distribution networks. Water Res. 2013, 47, 3015–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, M.F.; Vesey, G.; Look, N.L.; Herbert, B.R.; Simpson-Stroot, J.M.; Lim, D.V. Development of a Cryptosporidium oocyst assay using an automated fiber optic-based biosensor. J. Biol. Eng. 2007, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Baeumner, A.J. Highly sensitive and specific detection of viable Escherichia coli in drinking water. Anal. Biochem. 2002, 303, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.K.; Sharma, M.K.; Goel, A.K.; Singh, L.; Sekhar, K. Amperometric immunosensor for the detection of Vibrio cholerae O1 using disposable screen-printed electrodes. Anal. Sci. 2006, 22, 1207–1211. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; He, M.; Shi, H.C.; Zhu, A.N. Development of evanescent wave all-fiber immunosensor for environmental water analysis. Biosens. Bioelectron. 2008, 23, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Long, F.; He, M.; Zhu, A.N.; Shi, H.C. Portable optical immunosensor for highly sensitive detection of microcystin-LR in water samples. Biosens. Bioelectron. 2009, 24, 2346–2351. [Google Scholar] [CrossRef] [PubMed]
- Strepparava, N.; Wahli, T.; Segner, H.; Polli, B.; Petrini, O. Fluorescent in situ hybridization: A new tool for the direct identification and detection of F. psychrophilum. PLoS ONE 2012, 7, e49280. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Yilmaz, L.S.; Noguera, D.R.; Daims, H.; Wagner, M. Quantification of target molecules needed to detect microorganisms by fluorescence in situ hybridization (FISH) and catalyzed reporter deposition-FISH. Appl. Environ. Microbiol. 2008, 74, 5068–5077. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Jee, Y.-H.; Ko, G. Immunomagnetic separation combined with real-time reverse transcriptase PCR assays for detection of norovirus in contaminated food. Appl. Environ. Microbiol. 2008, 74, 4226–4230. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, Y.T.; Kim, Y.K. Optical enzyme-linked immunosorbant assay on a strip for detection of Salmonella typhimurium. Biochip J. 2010, 4, 110–116. [Google Scholar] [CrossRef]
- Mandal, P.K.; Biswas, A.K.; Choi, K.; Pal, U.K. Methods for rapid detection of foodborne pathogens: An overview. Am. J. Food. Technol. 2011, 6, 8–102. [Google Scholar]
- Hartman, L.J.; Coyne, S.R.; Norwood, D.A. Development of a novel internal positive control for Taqman based assays. Mol. Cell Probes 2005, 19, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Omar, K.B.; Barnard, T.G. Detection of diarrhoeagenic Escherichia coli in clinical and environmental water sources in South Africa using single-step 11-gene m-PCR. World J. Microbiol. Biotechnol. 2014, 30, 2663–2671. [Google Scholar] [CrossRef] [PubMed]
- Valasek, M.A.; Repa, J.J. The power of real-time PCR. Adv. Physiol. Educ. 2005, 29, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Omiccioli, E.; Amagliani, G.; Brandi, G.; Magnani, M. A new platform for real-time PCR detection of Salmonella spp., Listeria monocytogenes and Escherichia coli O157 in milk. Food Microbiol. 2009, 26, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Hein, I.; Lehner, A.; Rieck, P.; Klein, K.; Brandl, E.; Wagner, M. Comparison of different approaches to quantify Staphylococcus aureus cells by real-time quantitative PCR and application of this technique for examination of cheese. Appl. Environ. Microbiol. 2001, 67, 3122–3126. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Watt, P.M.; Grieve, C.M.; Sharma, V.K.; Lyons, S.R. Multiplex fluorogenic real-time PCR for detection and quantification of Escherichia coli O157:H7 in dairy wastewater wetlands. Appl. Environ. Microbiol. 2002, 68, 4853–4862. [Google Scholar] [CrossRef] [PubMed]
- LaGier, M.J.; Joseph, L.A.; Passaretti, T.V.; Musser, K.A.; Cirino, N.M. A real-time multiplexed PCR assay for rapid detection and differentiation of Campylobacter jejuni and Campylobacter coli. Mol. Cell Probes 2004, 18, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Segawa, T.; Okabe, S. Simultaneous quantification of multiple food- and waterborne pathogens by use of microfluidic quantitative PCR. Appl. Environ. Microbiol. 2013, 79, 2891–2898. [Google Scholar] [CrossRef] [PubMed]
- Life Technologies, Biotrove Open Array System. Available on: http://www.lifetechnologies.com/ (assessed on 17 February 2015).
- Bertrand, I.; Gantzer, C.; Chesnot, T.; Schwartzbrod, J. Improved specificity for Giardia lamblia cyst quantification in wastewater by development of a real-time PCR method. J. Microbiol. Methods 2004, 57, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, K.A.; Griffin, D.W.; Paul, J.H. Detection, quantitation and identification of enteroviruses from surface waters and sponge tissue from the florida keys using real-time RT-PCR. Water Res. 2002, 36, 2505–2514. [Google Scholar] [CrossRef] [PubMed]
- Baeumner, A.J.; Humiston, M.C.; Montagna, R.A.; Durst, R.A. Detection of viable oocysts of Cryptosporidium parvum following nucleic acid sequence based amplification. Anal. Chem. 2001, 73, 1176–1180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J. Microarrays for bacterial detection and microbial community analysis. Curr. Opin. Microbiol. 2003, 6, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Trevino, V.; Falciani, F.; Barrera-Saldana, H.A. DNA microarrays: A powerful genomic tool for biomedical and clinical research. Mol. Med. 2007, 13, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Severgnini, M.; Cremonesi, P.; Consolandi, C.; De Bellis, G.; Castiglioni, B. Advances in DNA microarray technology for the detection of foodborne pathogens. Food Bioprocess. Tech. 2011, 4, 936–953. [Google Scholar] [CrossRef]
- Gilbride, K.A.; Lee, D.Y.; Beaudette, L.A. Molecular techniques in wastewater: Understanding microbial communities, detecting pathogens, and real-time process control. J. Microbiol. Methods 2006, 66, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Inoue, D.; Hinoura, T.; Suzuki, N.; Pang, J.; Malla, R.; Shrestha, S.; Chapagain, S.K.; Matsuzawa, H.; Nakamura, T.; Tanaka, Y.; et al. High-throughput DNA microarray detection of pathogenic bacteria in shallow well groundwater in the Kathmandu Valley, Nepal. Curr. Microbiol. 2015, 70, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.V.; Hashsham, S.A.; Gulari, E.; Rouillard, J.M.; Aw, T.G.; Rose, J.B. Detection and characterization of human pathogenic viruses circulating in community wastewater using multi target microarrays and polymerase chain reaction. J. Water Health 2013, 11, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Urisman, A.; Liu, Y.T.; Springer, M.; Ksiazek, T.G.; Erdman, D.D.; Mardis, E.R.; Hickenbotham, M.; Magrini, V.; Eldred, J.; et al. Viral discovery and sequence recovery using DNA microarrays. PLoS Biol. 2003, 1, E2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leski, T.A.; Lin, B.; Malanoski, A.P.; Wang, Z.; Long, N.C.; Meador, C.E.; Barrows, B.; Ibrahim, S.; Hardick, J.P.; Aitichou, M.; et al. Testing and validation of high density resequencing microarray for broad range biothreat agents detection. PLoS ONE 2009, 4, e6569. [Google Scholar] [CrossRef] [PubMed]
- Vahey, M.; Nau, M.E.; Barrick, S.; Cooley, J.D.; Sawyer, R.; Sleeker, A.A.; Vickerman, P.; Bloor, S.; Larder, B.; Michael, N.L.; et al. Performance of the Affymetrix GeneChip HIV PRT 440 platform for antiretroviral drug resistance genotyping of human immunodeficiency virus type 1 clades and viral isolates with length polymorphisms. J. Clin. Microbiol. 1999, 37, 2533–2537. [Google Scholar] [PubMed]
- Nau, M.E.; Emerson, L.R.; Martin, R.K.; Kyle, D.E.; Wirth, D.F.; Vahey, M. Technical assessment of the Affymetrix yeast expression GeneChip ye6100 platform in a heterologous model of genes that confer resistance to antimalarial drugs in yeast. J. Clin. Microbiol. 2000, 38, 1901–1908. [Google Scholar] [PubMed]
- Nelson, T.A.; Holmes, S.; Alekseyenko, A.V.; Shenoy, M.; Desantis, T.; Wu, C.H.; Andersen, G.L.; Winston, J.; Sonnenburg, J.; Pasricha, P.J.; et al. Phylochip microarray analysis reveals altered gastrointestinal microbial communities in a rat model of colonic hypersensitivity. Neurogastroenterol. Motil. 2011, 23, 169–177, e141–e162. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.G.; Rose, J.B. Detection of pathogens in water: From phylochips to qPCR to pyrosequencing. Curr. Opin. Biotechnol. 2012, 23, 422–430. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Brodie, E.L.; Moberg, J.P.; Zubieta, I.X.; Piceno, Y.M.; Andersen, G.L. High-density universal 16S rRNA microarray analysis reveals broader diversity than typical clone library when sampling the environment. Microb Ecol. 2007, 53, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, A.; Ehn, M.; Hober, S. Pyrosequencing: History, biochemistry and future. Clin. Chim. Acta 2006, 363, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.F., Jr.; Fouad, A.F.; Rocas, I.N. Pyrosequencing as a tool for better understanding of human microbiomes. J. Oral Microbiol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.Z.; Ishoey, T.; Novotny, M.A.; McLean, J.S.; Lasken, R.S.; Williamson, S.J. Single virus genomics: A new tool for virus discovery. PLoS ONE 2011, 6, e17722. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Foo, S.H.; Lin, L.; Liu, E.T.; Gharizadeh, B.; Goel, S. Pyrosequencing enhancement for better detection limit and sequencing homopolymers. Biochem. Biophys. Res. Commun. 2010, 401, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.Y.; Hwang, C.; Ling, F.; Andersen, G.L.; LeChevallier, M.W.; Liu, W.T. Pyrosequencing analysis of bacterial biofilm communities in water meters of a drinking water distribution system. Appl. Environ. Microbiol. 2010, 76, 5631–5635. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Leddy, M.; Murinda, S.E. Potential human pathogenic bacteria in a mixed urban watershed as revealed by pyrosequencing. PLoS ONE 2013, 8, e79490. [Google Scholar] [CrossRef] [PubMed]
- Van der Wielen, P.W.; Heijnen, L.; van der Kooij, D. Pyrosequence analysis of the hsp65 genes of nontuberculous Mycobacterium communities in unchlorinated drinking water in the Netherlands. Appl. Environ. Microbiol. 2013, 79, 6160–6166. [Google Scholar] [CrossRef] [PubMed]
- Djikeng, A.; Kuzmickas, R.; Anderson, N.G.; Spiro, D.J. Metagenomic analysis of RNA viruses in a fresh water lake. PLoS ONE 2009, 4, e7264. [Google Scholar] [CrossRef] [PubMed]
- Allegra, S.; Berger, F.; Berthelot, P.; Grattard, F.; Pozzetto, B.; Riffard, S. Use of flow cytometry to monitor Legionella viability. Appl. Environ. Microbiol. 2008, 74, 7813–7816. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Horn, M.; Daims, H. Fluorescence in situ hybridisation for the identification and characterisation of prokaryotes. Curr. Opin. Microbiol. 2003, 6, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Alagappan, A.; Bergquist, P.L.; Ferrari, B.C. Development of a two-color fluorescence in situ hybridization technique for species-level identification of human-infectious Cryptosporidium spp. Appl. Environ. Microbiol. 2009, 75, 5996–5998. [Google Scholar] [CrossRef] [PubMed]
- Mirus. Label. IT Fluorescence In Situ Hybridization Kits. Available online: https://www.mirusbio.com/products/labeling/label-it-fluorescence-in-situ-hybridization-kits (assessed on 17 February 2015).
- Malic, S.; Hill, K.E.; Hayes, A.; Percival, S.L.; Thomas, D.W.; Williams, D.W. Detection and identification of specific bacteria in wound biofilms using peptide nucleic acid fluorescent in situ hybridization (PNA FISH). Microbiology 2009, 155, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Wingender, J.; Flemming, H.C. Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health 2011, 214, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Armisen, T.; Servais, P. Enumeration of viable E. coli in rivers and wastewaters by fluorescent in situ hybridization. J. Microbiol. Methods 2004, 58, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D. The Public Health Significance of Viable but Nonculturable Bacteria. In Nonculturable Microorganisms in the Environment; Colwell Grimes, D.J., Ed.; ASM Press: Washington, DC, USA, 2000; pp. 277–299. [Google Scholar]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [PubMed]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [PubMed]
- Rowan, N.J. Viable but non-culturable forms of food and waterborne bacteria: Quo vadis? Trends Food Sci. Technol. 2004, 15, 462–467. [Google Scholar] [CrossRef]
- Dwidjosiswojo, Z.; Richard, J.; Moritz, M.M.; Dopp, E.; Flemming, H.C.; Wingender, J. Influence of copper ions on the viability and cytotoxicity of Pseudomonas aeruginosa under conditions relevant to drinking water environments. Int. J. Hyg. Environ. Health 2011, 214, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Almeida, V.; Almeida Santos, M.; Chambel, L. Resuscitation of Escherichia coli VBNC cells depends on a variety of environmental or chemical stimuli. J. Appl. Microbiol. 2011, 110, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.S.; Mayo, M.W.; Bruno, J.G.; Bronk, B.V.; Batt, C.A.; Chambers, J.P. A review of molecular recognition technologies for detection of biological threat agents. Biosens. Bioelectron. 2000, 15, 549–578. [Google Scholar] [CrossRef] [PubMed]
- Velusamy, V.; Arshak, K.; Korostynska, O.; Oliwa, K.; Adley, C. An overview of foodborne pathogen detection: In the perspective of biosensors. Biotechnol Adv. 2010, 28, 232–254. [Google Scholar] [CrossRef] [PubMed]
- Connelly, J.T.; Baeumner, A.J. Biosensors for the detection of waterborne pathogens. Anal. Bioanal. Chem. 2012, 402, 117–127. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Method 1623.1: Cryptosporidium and Giardia in Water by Filtration/IMS/FA; Water, E.P.A., Ed.; U.S. Environmental Protection Agency: Washington, DC, USA, 2012.
- U.S. Environmental Protection Agency. Method 1622: Cryptosporidium in Water by filtration/ims/fa; Water, E.P.A., Ed.; U.S. Environmental Protection Agency: Washington, DC, USA, 1999.
- Bridle, H.; Desmulliez, M. Biosensors for the Detection of Waterborne Pathogens. In Waterborne Pathogens. Detection Methods and Applications; Bridle, H., Ed.; Elsevier B.V.: London, UK, 2014. [Google Scholar]
- Xu, S.; Mutharasan, R. The Coming Together of the Sciences: Biosensors for the Detection of Waterborne Pathogens Using antibodies and Gene-Based Recognition Chemistries. In Environmental Microbiology: Current Technology and Water Applications; Sen, K., Ashbolt, N.J., Eds.; Caister Academic Press: Norfolk, VA, USA, 2011. [Google Scholar]
- Singh, A.; Poshtiban, S.; Evoy, S. Recent advances in bacteriophage based biosensors for food-borne pathogen detection. Sensors 2013, 13, 1763–1786. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.D.; Yu, Q.; Chen, S.; Homola, J.; Jiang, S. Comparison of E. coli O157:H7 preparations methods used for detection with surface plasmon resonance sensor. Sensors and Actuat B: Cheml 2005, 107, 202–208. [Google Scholar] [CrossRef]
- Ho, J.A.; Hsu, H.W.; Huang, M.R. Liposome-based microcapillary immunosensor for detection of Escherichia coli O157:H7. Anal. Biochem. 2004, 330, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Pires, N.M.; Dong, T.; Hanke, U.; Hoivik, N. Recent developments in optical detection technologies in lab-on-a chip devices for biosensing applications. Sensors (Basel). 2014, 15458–15479. [Google Scholar]
- Yoon, J.Y.; Kim, B. Lab-on-a-chip pathogen sensors for food safety. Sensors 2012, 12, 10713–10741. [Google Scholar] [CrossRef] [PubMed]
- Ashbolt, N.J.; Schoen, M.E.; Soller, J.A.; Roser, D.J. Predicting pathogen risks to aid beach management: The real value of quantitative microbial risk assessment (QMRA). Water Res. 2010, 44, 4692–4703. [Google Scholar] [CrossRef] [PubMed]
- Dunn, G.; Harris, L.; Cook, C.; Prystajecky, N. A comparative analysis of current microbial water quality risk assessment and management practices in British Columbia and Ontario, Canada. Sci. Total Environ. 2014, 468–469, 544–552. [Google Scholar]
- Haas, C.N.; Rose, J.B.; Gerba, C.P. Quantitative Microbial Risk Assessment, 1st ed.; John Wiley & Sons: New York, NY, USA, 1999; p. 464. [Google Scholar]
- Haas, C.N.; Eisenberg, J.N.S. Risk Assessmen. In World Health Organization (WHO). Water Quality: Guidelines, Standards and Health; Fewtrell, L., Bartram, J., Eds.; IWA Publishing: London, UK, 2001. [Google Scholar]
- Hunter, P.R.; Payment, P.; Ashbolt, N.; Bartram, J. Assessment of risk. In Assessing Microbial Safety of Drinking Water: Improving Approaches and Methods; Dufour, A., Snozzi, M., Koster, W., Bartram, J., Ronchi, E., Fewtrell, L., Eds.; World Health Organization by IWA Publishing, 2003; pp. 79–103. [Google Scholar]
- U.S. Department of Agriculture/Food Safety and Inspection Service (USDA/FSIS) and U.S. Environmental Protection Agency (EPA). Microbial Risk Assessment Guideline: Pathogenic Microorganisms with Focus on Food and Water, USDA/FSIS and EPA Ed. ed; FSIS Publication; EPA Publication: USA, 2012. [Google Scholar]
- Teunis, P.F.; Ogden, I.D.; Strachan, N.J. Hierarchical dose response of E. coli O157:H7 from human outbreaks incorporating heterogeneity in exposure. Epidemiol. Infect. 2008, 136, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Schoen, M.E.; Ashbolt, N.J. Assessing pathogen risk to swimmers at non-sewage impacted recreational beaches. Environ. Sci. Technol. 2010, 44, 2286–2291. [Google Scholar] [CrossRef] [PubMed]
- Messner, M.J.; Chappell, C.L.; Okhuysen, P.C. Risk assessment for Cryptosporidium: A hierarchical bayesian analysis of human dose response data. Water Res. 2001, 35, 3934–3940. [Google Scholar] [CrossRef] [PubMed]
- Whelan, G.; Kim, K.; Pelton, M.A.; Soller, J.A.; Castleton, K.J.; Molina, M.; Pachepsky, Y.; Zepp, R. Environ. Modell. Softw. Integrated Environ. Model. Framew. Perform. Quant. Microb. Risk Assess. 2014, 55, 77–91. [Google Scholar]
- Nauta, M.J. Modelling bacterial growth in quantitative microbiological risk assessment: Is it possible? Int. J. Food Microbiol. 2002, 73, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Reynols, K.A.; Mena, K.D. Quantitative Microbial Risk Assessment of Waterborne Disease. In Global Issues in Water, Sanitation, and Health: Workshop Summary, Institute of Medicine (US) Forum on Microbial Threats Ed.; National Academies Press (US): Washington, DC, USA, 2009; pp. 178–199. [Google Scholar]
- World Health Organization (WHO). Guidelines for Safe Recreational Water Environments Volume 1 Coastal and Fresh Waters; World Health Organization (WHO): Geneva, Switzerland, 2003. [Google Scholar]
- Blackmer, F.; Reynolds, K.A.; Gerba, C.P.; Pepper, I.L. Use of integrated cell culture-PCR to evaluate the effectiveness of poliovirus inactivation by chlorine. Appl. Environ. Microbiol. 2000, 66, 2267–2268. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, K.A.; Gerba, C.P.; Abbaszadegan, M.; Pepper, L.L. ICC/PCR detection of enteroviruses and hepatitis a virus in environmental samples. Can. J. Microbiol. 2001, 47, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, S.; Olaniran, A.O. Treated wastewater effluent as a source of microbial pollution of surface water resources. Int. J. Environ. Res. Public Health 2014, 11, 249–270. [Google Scholar] [CrossRef]
- Machdar, E.; van der Steen, N.P.; Raschid-Sally, L.; Lens, P.N. Application of quantitative microbial risk assessment to analyze the public health risk from poor drinking water quality in a low income area in Accra, Ghana. Sci. Total Environ. 2013, 449, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Balleste, E.; Blanch, A.R. Bifidobacterial diversity and the development of new microbial source tracking indicators. Appl. Environ. Microbiol. 2011, 77, 3518–3525. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.; Ufnar, J.A.; Griffith, J.F.; Gooch, J.A.; Stewart, J.R. A real-time qPCR assay for the detection of the nifH gene of Methanobrevibacter smithii, a potential indicator of sewage pollution. J. Appl. Microbiol. 2010, 109, 1946–1956. [Google Scholar] [CrossRef] [PubMed]
- Villemur, R.; Imbeau, M.; Vuong, M.N.; Masson, L.; Payment, P. An environmental survey of surface waters using mitochondrial DNA from human, bovine and porcine origin as fecal source tracking markers. Water Res. 2015, 69, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, J.M.; Raley, M.E.; Levine, J.F. Mitochondrial multiplex real-time PCR as a source tracking method in fecal-contaminated effluents. Environ. Sci. Technol. 2007, 41, 3277–3283. [Google Scholar] [CrossRef] [PubMed]
- Schijven, J.F.; Teunis, P.F.; Rutjes, S.A.; Bouwknegt, M.; de Roda Husman, A.M. QMRAspot: A tool for quantitative microbial risk assessment from surface water to potable water. Water Res. 2011, 45, 5564–5576. [Google Scholar] [CrossRef] [PubMed]
- Tseng, L.Y.; Jiang, S.C. Comparison of recreational health risks associated with surfing and swimming in dry weather and post-storm conditions at Southern California beaches using quantitative microbial risk assessment (QMRA). Mar. Pollut. Bull. 2012, 64, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Ten Veldhuis, J.A.; Clemens, F.H.; Sterk, G.; Berends, B.R. Microbial risks associated with exposure to pathogens in contaminated urban flood water. Water Res. 2010, 44, 2910–2918. [Google Scholar] [CrossRef] [PubMed]
- Blokker, M.; Smeets, P.; Medema, G. QMRA in drinking water distribution system. Proc. Eng. 2014, 89, 151–159. [Google Scholar] [CrossRef]
- Soller, J.A.; Schoen, M.E.; Bartrand, T.; Ravenscroft, J.E.; Ashbolt, N.J. Estimated human health risks from exposure to recreational waters impacted by human and non-human sources of faecal contamination. Water Res. 2010, 44, 4674–4691. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Castillo, F.Y.; Loera-Muro, A.; Jacques, M.; Garneau, P.; Avelar-González, F.J.; Harel, J.; Guerrero-Barrera, A.L. Waterborne Pathogens: Detection Methods and Challenges. Pathogens 2015, 4, 307-334. https://doi.org/10.3390/pathogens4020307
Ramírez-Castillo FY, Loera-Muro A, Jacques M, Garneau P, Avelar-González FJ, Harel J, Guerrero-Barrera AL. Waterborne Pathogens: Detection Methods and Challenges. Pathogens. 2015; 4(2):307-334. https://doi.org/10.3390/pathogens4020307
Chicago/Turabian StyleRamírez-Castillo, Flor Yazmín, Abraham Loera-Muro, Mario Jacques, Philippe Garneau, Francisco Javier Avelar-González, Josée Harel, and Alma Lilián Guerrero-Barrera. 2015. "Waterborne Pathogens: Detection Methods and Challenges" Pathogens 4, no. 2: 307-334. https://doi.org/10.3390/pathogens4020307