
Deliberate Establishment of Asymptomatic Bacteriuria—A Novel Strategy to Prevent Recurrent UTI

{kind=link}
{kind=link}
Abstract
:1. Background
2. Properties of ABU Strains
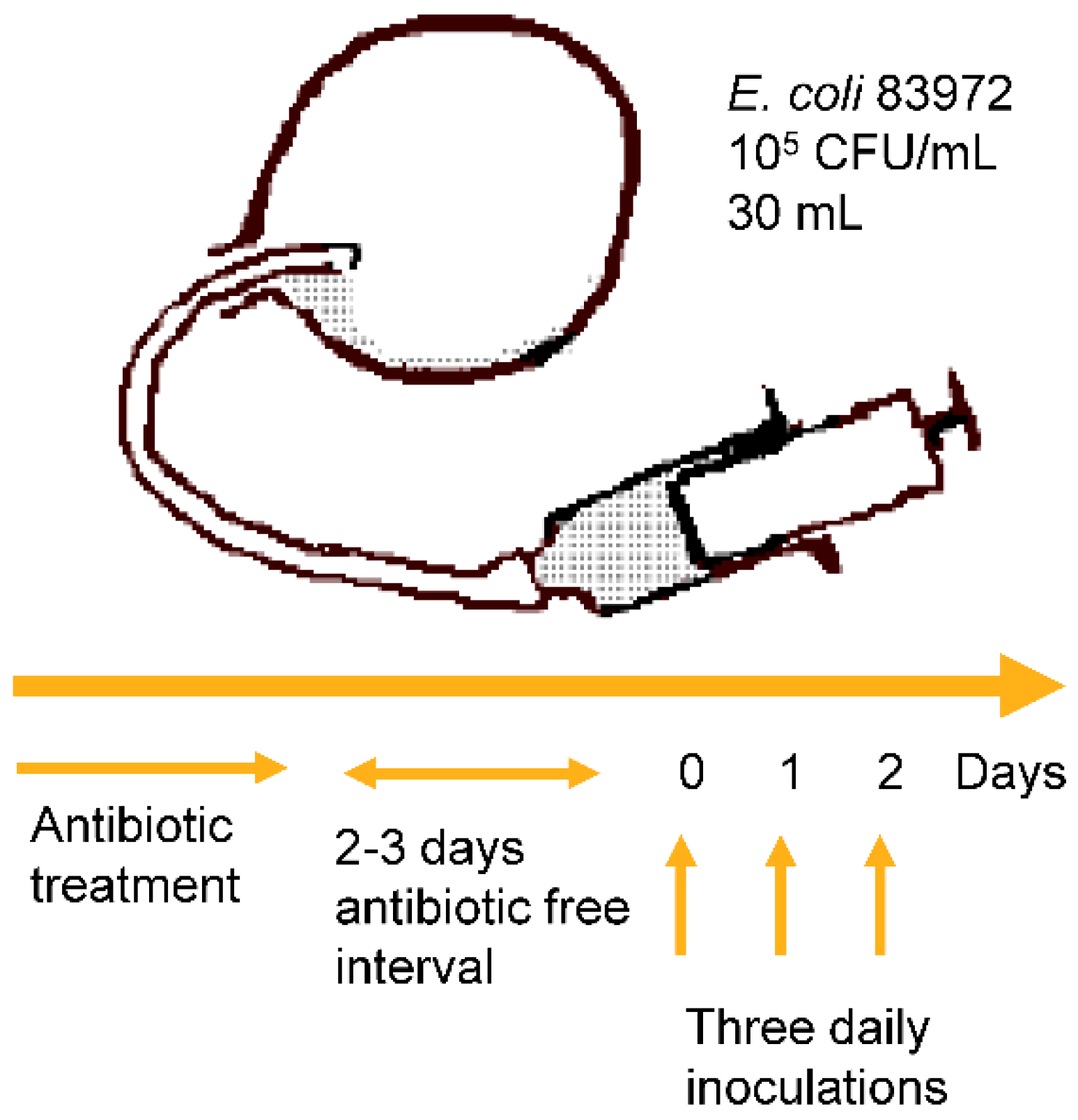
2.1. E. coli 83972 and the Inoculation Protocol
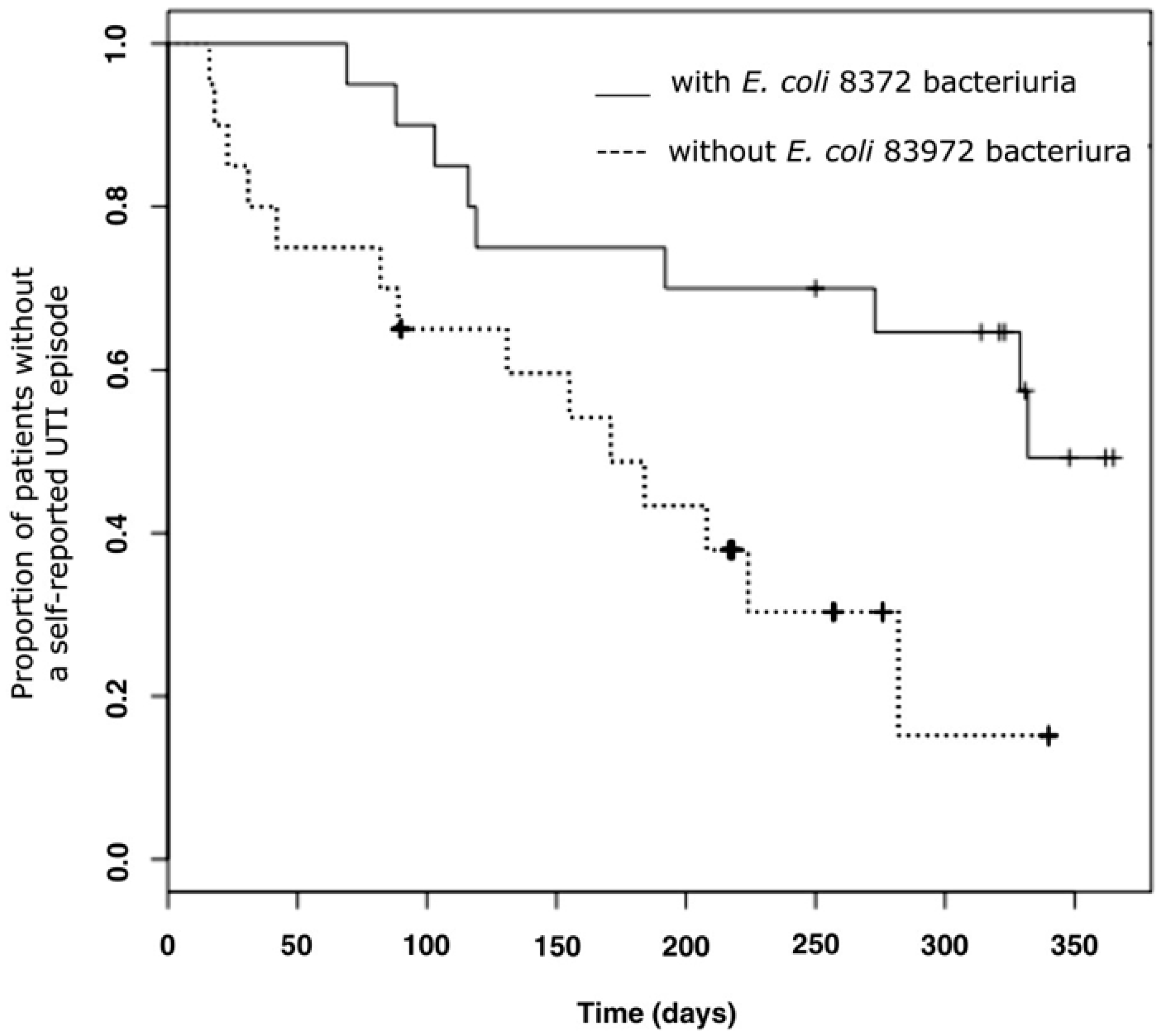
2.2. E. coli 83972 Inoculation Protects against Symptomatic UTI
3. Studies of Bacteria and Their Infected Hosts
3.1. The Role of the P and Type 1 Fimbriae
3.1.1. Fimbriated Transformants of E. coli 83972
3.1.2. Reconstituted Chromosomal Fimbrial Gene Clusters
3.2. Host Response to E. coli 83972 in Inoculated Human Hosts
3.3. Genetic Control of the Innate Immune Response during E. coli 83972 Inoculation
4. Assessment of Risk that E. coli 83972 Regains Virulence in the Human Urinary Tract
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
References
- Wullt, B.; Bergsten, G.; Fischer, H.; Godaly, G.; Karpman, D.; Leijonhufvud, I.; Lundstedt, A.C.; Samuelsson, P.; Samuelsson, M.; Svensson, M.L.; et al. The host response to urinary tract infection. Infect. Dis. Clin. North Am. 2003, 17, 279–301. [Google Scholar] [CrossRef]
- Kass, E.H. Asymptomatic infections of the urinary tract. Trans. Assoc. Am. Physicians 1956, 69, 56–64. [Google Scholar] [PubMed]
- Lindberg, U. Asymptomatic bacteriuria in schoolgirls. V. The clinical course and response to treatment. Acta Paediatr. Scand. 1975, 64, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Stenqvist, K.; Sandberg, T.; Lidin-Janson, G.; Orskov, F.; Orskov, I.; Svanborg-Eden, C. Virulence factors of Escherichia coli in urinary isolates from pregnant women. J. Infect. Dis. 1987, 156, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Nordenstam, G.; Sundh, V.; Lincoln, K.; Svanborg, A.; Eden, C.S. Bacteriuria in representative population samples of persons aged 72–79 years. Am. J. Epidemiol. 1989, 130, 1176–1186. [Google Scholar] [PubMed]
- Nicolle, L.E. Consequences of asymptomatic bacteriuria in the elderly. Int. J. Antimicrob. Agents 1994, 4, 107–111. [Google Scholar] [CrossRef]
- Nicolle, L.E.; Bradley, S.; Colgan, R.; Rice, J.C.; Schaeffer, A.; Hooton, T.M.; Infectious Diseases Society of America; American Society of Nephrology; American Geriatric Society. Infectious diseases society of america guidelines for the diagnosis and treatment of asymptomatic bacteriuria in adults. Clin. Infect. Dis. 2005, 40, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Foxman, B.; Brown, P. Epidemiology of urinary tract infections: Transmission and risk factors, incidence, and costs. Infect. Dis. Clin. North Am. 2003, 17, 227–241. [Google Scholar] [CrossRef]
- Hansson, S.; Jodal, U.; Lincoln, K.; Svanborg-Eden, C. Untreated asymptomatic bacteriuria in girls: Ii--effect of phenoxymethylpenicillin and erythromycin given for intercurrent infections. BMJ 1989, 298, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Mazzoli, S.; Mondaini, N.; Meacci, F.; Nesi, G.; D'Elia, C.; Malossini, G.; Boddi, V.; Bartoletti, R. The role of asymptomatic bacteriuria in young women with recurrent urinary tract infections: To treat or not to treat? Clin. Infect. Dis. 2012, 55, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, U.; Claesson, I.; Hanson, L.A.; Jodal, U. Asymptomatic bacteriuria in schoolgirls. Viii. Clinical course during a 3-year follow-up. J. Pediatr. 1978, 92, 194–199. [Google Scholar] [CrossRef]
- Hansson, S.; Jodal, U.; Norén, L. Treatment vs. Non-treatment of asymptomatic bacteriuria in girlas with renal scarring. In Host-parasite Interactions in Urinary Tract Infections; Kass, E.H., Svanborg Edén, C., Eds.; University of Chicago Press: Chicago, IL, USA, 1989; pp. 289–291. [Google Scholar]
- Claesson, I.; Lindberg, U. Asymptomatic bacteriuria in schoolgirls. Vii. A follow-up study of the urinary tract in treated and untreated schoolgirls with asymptomatic bacteriuria. Radiology 1977, 124, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Grabe, M.; Bjerklund-Johansen, T.E.; Botto, H.; Wullt, B.; Çek, M.; Naber, K.G.; Pickard, R.S.; Tenke, P.; Wagenlehner, F. Individual Guidelines. 2014. Available online: Http://www.Uroweb.Org/professional-resources/guidelines/ (accessed on 20 July 2016).
- Hagberg, L.; Bruce, A.W.; Reid, G.; Svanborg-Eden, C.; Lincoln, K.; Lidin-Janson, G. Colonisation of the urinary tract with live bacteria from the normal fecal and urethral flora in patients with recurrent symptomatic urinary tract infections. In Host-parasite Interactions in Urinary Tract Infections; Kass, E.H., Svanborg Eden, C., Eds.; University of Chicago Press: Chicago, IL, USA, 1989; pp. 194–197. [Google Scholar]
- Sunden, F.; Hakansson, L.; Ljunggren, E.; Wullt, B. Escherichia coli 83972 bacteriuria protects against recurrent lower urinary tract infections in patients with incomplete bladder emptying. J. Urol. 2010, 184, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Darouiche, R.O.; Hull, R.A. Bacterial interference for prevention of urinary tract infection. Clin. Infect. Dis. 2012, 55, 1400–1407. [Google Scholar] [CrossRef] [PubMed]
- Darouiche, R.O.; Thornby, J.I.; Cerra-Stewart, C.; Donovan, W.H.; Hull, R.A. Bacterial interference for prevention of urinary tract infection: A prospective, randomized, placebo-controlled, double-blind pilot trial. Clin. Infect. Dis. 2005, 41, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Wullt, B.; Connell, H.; Rollano, P.; Mansson, W.; Colleen, S.; Svanborg, C. Urodynamic factors influence the duration of Escherichia coli bacteriuria in deliberately colonized cases. J. Urol. 1998, 159, 2057–2062. [Google Scholar] [CrossRef]
- Hull, R.A.; Rudy, D.C.; Donovan, W.H.; Wieser, I.E.; Stewart, C.; Darouiche, R.O. Virulence properties of Escherichia coli 83972, a prototype strain associated with asymptomatic bacteriuria. Infect. Immun. 1999, 67, 429–432. [Google Scholar] [PubMed]
- Mabbett, A.N.; Ulett, G.C.; Watts, R.E.; Tree, J.J.; Totsika, M.; Ong, C.L.; Wood, J.M.; Monaghan, W.; Looke, D.F.; Nimmo, G.R.; et al. Virulence properties of asymptomatic bacteriuria Escherichia coli. Int. J. Med. Microbiol. 2009, 299, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Svanborg-Edén, C.; Hanson, L.A.; Jodal, U.; Lindberg, U.; Sohl-Åkerlund, A. Variable adherence to normal urinary tract epithelial cells of Escherichia coli strains associated with various forms of urinary tract infections. Lancet 1976, II, 490–492. [Google Scholar] [CrossRef]
- Kaijser, B.; Hanson, L.A.; Jodal, U.; Lidin-Janson, G.; Robbins, J.B. Frequency of E. coli k antigens in urinary-tract infections in children. Lancet 1977, 1, 663–666. [Google Scholar] [CrossRef]
- Kallenius, G.; Mollby, R.; Svenson, S.B.; Helin, I.; Hultberg, H.; Cedergren, B.; Winberg, J. Occurrence of p-fimbriated Escherichia coli in urinary tract infections. Lancet 1981, 2, 1369–1372. [Google Scholar] [CrossRef]
- Lomberg, H.; Hellstrom, M.; Jodal, U.; Svanborg-Eden, C. Renal scarring and non-attaching Escherichia coli. Lancet 1986, 2, 1341. [Google Scholar] [CrossRef]
- Vaisanen-Rhen, V.; Elo, J.; Vaisanen, E.; Siitonen, A.; Orskov, I.; Orskov, F.; Svenson, S.B.; Makela, P.H.; Korhonen, T.K. P-fimbriated clones among uropathogenic Escherichia coli strains. Infect. Immun. 1984, 43, 149–155. [Google Scholar] [PubMed]
- Caugant, D.A.; Levin, B.R.; Lidin-Janson, G.; Whittam, T.S.; Svanborg Eden, C.; Selander, R.K. Genetic diversity and relationships among strains of Escherichia coli in the intestine and those causing urinary tract infections. Prog. Allergy 1983, 33, 203–227. [Google Scholar] [PubMed]
- Zdziarski, J.; Svanborg, C.; Wullt, B.; Hacker, J.; Dobrindt, U. Molecular basis of commensalism in the urinary tract: Low virulence or virulence attenuation? Infect. Immun. 2008, 76, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Lidin-Janson, G.; Lindberg, U. Asymptomatic bacteriuria in schoolgirls. Vi. The correlation between urinary and faecal Escherichia coli. Relation to the duration of the bacteriuria and the sampling technique. Acta Paediatr. Scand. 1977, 66, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Klemm, P.; Roos, V.; Ulett, G.C.; Svanborg, C.; Schembri, M.A. Molecular characterization of the Escherichia coli 83972 asymptomatic bacteriuria strain: The taming of a pathogen. Infect. Immun. 2006, 74, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Zdziarski, J.; Brzuszkiewicz, E.; Wullt, B.; Liesegang, H.; Biran, D.; Voigt, B.; Grönberg-Hernandez, J.; Ragnarsdottir, B.; Hecker, M.; Ron, E.Z.; et al. Host imprints on bacterial genomes--rapid, divergent evolution in individual patients. PLoS Pathog. 2010, 6, e1001078. [Google Scholar] [CrossRef] [PubMed]
- Andersson, P.; Engberg, I.; Lidin-Janson, G.; Lincoln, K.; Hull, R.; Hull, S.; Svanborg, C. Persistence of Escherichia coli bacteriuria is not determined by bacterial adherence. Infect. Immun. 1991, 59, 2915–2921. [Google Scholar] [PubMed]
- Wullt, B.; Bergsten, G.; Connell, H.; Rollano, P.; Gebretsadik, N.; Hull, R.; Svanborg, C. P fimbriae enhance the early establishment of Escherichia coli in the human urinary tract. Mol. Microbiol. 2000, 38, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Sunden, F.; Hakansson, L.; Ljunggren, E.; Wullt, B. Bacterial interference--is deliberate colonization with Escherichia coli 83972 an alternative treatment for patients with recurrent urinary tract infection? Int. J. Antimicrob. Agents 2006, 28 (Suppl 1), S26–S29. [Google Scholar] [CrossRef] [PubMed]
- Wullt, B.; Bergsten, G.; Connell, H.; Rollano, P.; Gebratsedik, N.; Hang, L.; Svanborg, C. P-fimbriae trigger mucosal responses to Escherichia coli in the human urinary tract. Cell. Microbiol. 2001, 3, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Leffler, H.; Svanborg-Edén, C. Glycolipid receptors for uropathogenic Escherichia coli on human erythrocytes and uroepithelial cells. Infect Immun. 1981, 34, 920–929. [Google Scholar] [PubMed]
- Hagberg, L.; Hull, R.; Hull, S.; Falkow, S.; Freter, R.; Svanborg Eden, C. Contribution of adhesion to bacterial persistence in the mouse urinary tract. Infect Immun. 1983, 40, 265–272. [Google Scholar] [PubMed]
- Schaeffer, A.J.; Schwan, W.R.; Hultgren, S.J.; Duncan, J.L. Relationship of type 1 pilus expression in Escherichia coli to ascending urinary tract infections in mice. Infect Immun. 1987, 55, 373–380. [Google Scholar] [PubMed]
- Connell, H.; Agace, W.; Klemm, P.; Schembri, M.; Mårild, S.; Svanborg, C. Type 1 fimbrial adhesion enhances Escherichia coli virulence for the urinary tract. Proc. Natl. Acad. Sci. USA 1996, 93, 9827–9832. [Google Scholar] [CrossRef] [PubMed]
- Bergsten, G.; Samuelsson, M.; Wullt, B.; Leijonhufvud, I.; Fischer, H.; Svanborg, C. Papg-dependent adherence breaks mucosal inertia and triggers the innate host response. J. Infect. Dis. 2004, 189, 1734–1742. [Google Scholar] [CrossRef] [PubMed]
- Wullt, B. The role of p fimbriae for Escherichia coli establishment and mucosal inflammation in the human urinary tract. Int. J. Antimicrob. Agents 2003, 21, 605–621. [Google Scholar] [CrossRef]
- Bergsten, G.; Wullt, B.; Schembri, M.A.; Leijonhufvud, I.; Svanborg, C. Do type 1 fimbriae promote inflammation in the human urinary tract? Cell. Microbiol. 2007, 9, 1766–1781. [Google Scholar] [CrossRef] [PubMed]
- Ambite, I.; Grönberg-Hernández, J.; Stork, C.; Köves, B.; Zdziarski, J.; Dobrindt, U.; Wullt, B.; Svanborg, C. Fimbriae reprogram host gene expression. A case of “virulence gene addiction”? 2016; submitted. [Google Scholar]
- Hernandez, J.G.; Sunden, F.; Connolly, J.; Svanborg, C.; Wullt, B. Genetic control of the variable innate immune response to asymptomatic bacteriuria. PLoS ONE 2011, 6, e28289. [Google Scholar] [CrossRef] [PubMed]
- Ragnarsdottir, B.; Samuelsson, M.; Gustafsson, M.C.; Leijonhufvud, I.; Karpman, D.; Svanborg, C. Reduced toll-like receptor 4 expression in children with asymptomatic bacteriuria. J. Infect. Dis. 2007, 196, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.; Lutay, N.; Ragnarsdottir, B.; Yadav, M.; Jonsson, K.; Urbano, A.; Al Hadad, A.; Ramisch, S.; Storm, P.; Dobrindt, U.; et al. Pathogen specific, irf3-dependent signaling and innate resistance to human kidney infection. PLoS Pathog. 2010, 6, e1001109. [Google Scholar] [CrossRef] [PubMed]
- Koves, B.; Salvador, E.; Gronberg-Hernandez, J.; Zdziarski, J.; Wullt, B.; Svanborg, C.; Dobrindt, U. Rare emergence of symptoms during long-term asymptomatic E. coli 83972 carriage, without altered virulence factor repertoire. J. Urol. 2013, 191, 519–528. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wullt, B.; Svanborg, C. Deliberate Establishment of Asymptomatic Bacteriuria—A Novel Strategy to Prevent Recurrent UTI. Pathogens 2016, 5, 52. https://doi.org/10.3390/pathogens5030052
Wullt B, Svanborg C. Deliberate Establishment of Asymptomatic Bacteriuria—A Novel Strategy to Prevent Recurrent UTI. Pathogens. 2016; 5(3):52. https://doi.org/10.3390/pathogens5030052
Chicago/Turabian StyleWullt, Björn, and Catharina Svanborg. 2016. "Deliberate Establishment of Asymptomatic Bacteriuria—A Novel Strategy to Prevent Recurrent UTI" Pathogens 5, no. 3: 52. https://doi.org/10.3390/pathogens5030052
APA StyleWullt, B., & Svanborg, C. (2016). Deliberate Establishment of Asymptomatic Bacteriuria—A Novel Strategy to Prevent Recurrent UTI. Pathogens, 5(3), 52. https://doi.org/10.3390/pathogens5030052