Transcription of IVIAT and Virulence Genes in Photobacterium damselae subsp. piscicida Infecting Solea senegalensis

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
2.2. Growth of Phdp under Iron Limiting Conditions and Oxidative Stress
2.3. Solea senegalensis Infection with Phdp
2.4. Bacteria Gene Expression Analysis
2.5. Bacteria Gene Expression Analysis under In Vivo Conditions
2.6. Statistical Analysis
2.7. Ethical Statements
3. Results
3.1. In Vitro Transcription of Virulence Genes
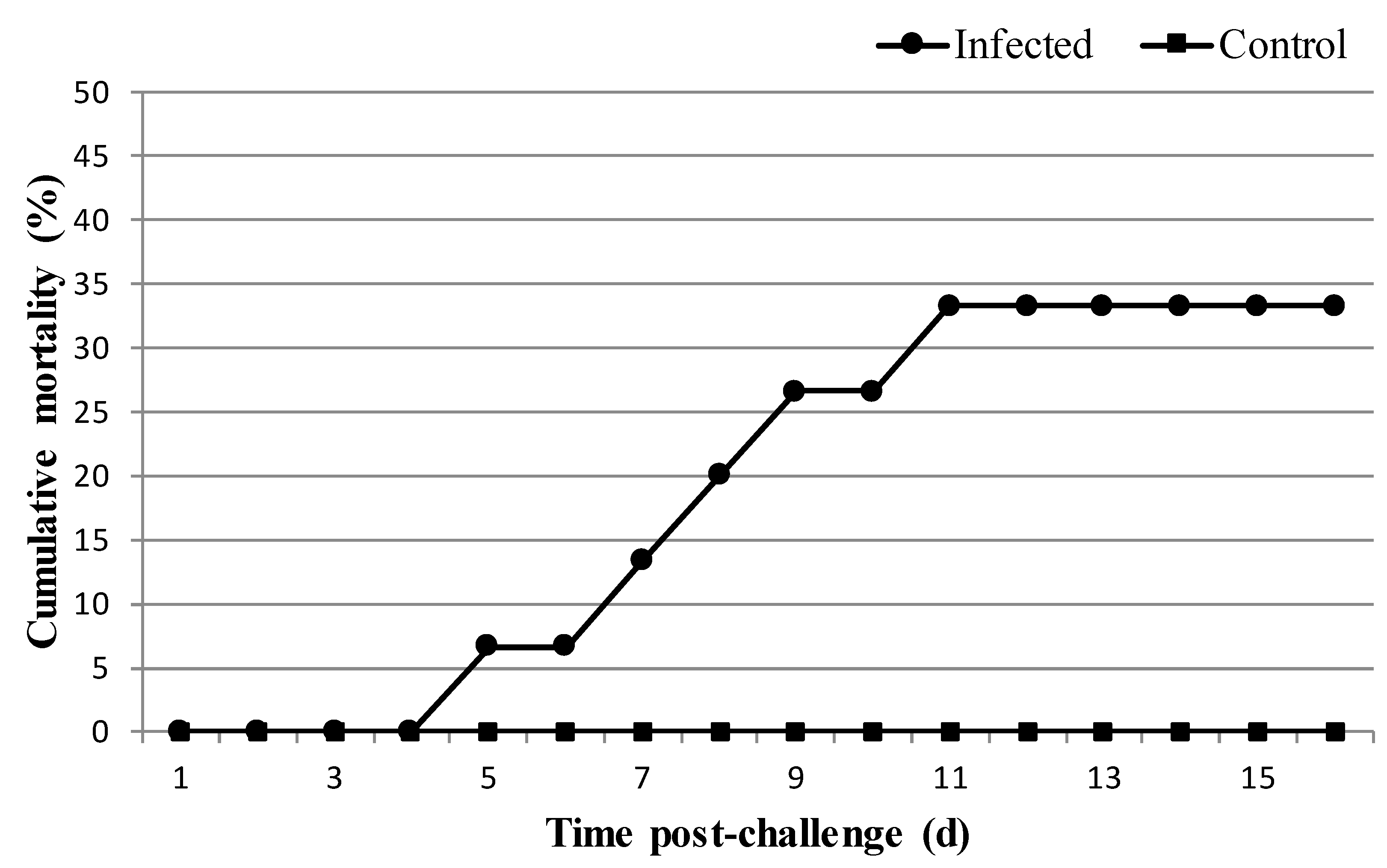
3.2. Experimental Infection
3.3. In Vivo Transcription of Phdp Virulence Genes
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Andreoni, F.; Magnani, M. Photobacteriosis: Prevention and Diagnosis. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Romalde, J.L. Photobacterium damselae subsp. piscicida: An integrated view of a bacterial fish pathogen. Int. Microbiol. 2002, 5, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Feng, J.; Su, Y.; Ye, L.; Wang, J. Studies on the isolation of Photobacterium damselae subsp. piscicida from diseased golden pompano (Trachinotus ovatus Linnaeus) and antibacterial agents sensitivity. Vet. Microbiol. 2013, 162, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Do Vale, A.; Silva, M.T.; dos Santos, N.M.S.; Nascimento, D.S.; Reis-Rodrigues, P.; Costa-Ramos, C.; Ellis, A.E.; Azevedo, J.E. AIP56, a novel plasmid-encoded virulence factor of Photobacterium damselae subsp. piscicida with apoptogenic activity against sea bass macrophages and neutrophils. Mol. Microbiol. 2005, 58, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Do Vale, A.; Costa-Ramos, C.; Silva, A.; Silva, D.S.P.; Gärtner, F.; dos Santos, N.M.S.; Silva, M.T. Systemic macrophage and neutrophil destruction by secondary necrosis induced by a bacterial exotoxin in a Gram-negative septicaemia. Cell Microbiol. 2007, 9, 988–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, D.S.; Pereira, L.M.G.; Moreira, A.R.; Ferreira-da-Silva, F.; Brito, R.M.; Faria, T.Q.; Zornetta, I.; Montecucco, C.; Oliveira, P.; Azevedo, J.E.; et al. The Apoptogenic Toxin AIP56 Is a Metalloprotease A-B Toxin that Cleaves NF-κb P65. PLoS Pathog. 2013, 9, e1003128. [Google Scholar] [CrossRef] [PubMed]
- Do Vale, A.; Cabanes, D.; Sousa, S. Bacterial toxins as pathogen weapons against phagocytes. Front. Microbiol. 2016, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.T.; Do Vale, A.; dos Santos, N.M.S. Secondary necrosis in multicellular animals: An outcome of apoptosis with pathogenic implications. Apoptosis 2008, 13, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Acosta, F.; Vivas, J.; Padilla, D.; Vega, J.; Bravo, J.; Grasso, V.; Real, F. Invasion and survival of Photobacterium damselae subsp. piscicida in non-phagocytic cells of gilthead sea bream, Sparus aurata L. J. Fish Dis. 2009, 32, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Doriga, M.V.; Barnes, A.C.; dos Santos, N.M.S.; Ellis, A.E. Invasion of fish epithelial cells by Photobacterium damselae subsp. piscicida: Evidence for receptor specificity, and effect of capsule and serum. Microbiology 2000, 146 Pt 1, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Elkamel, A.A.; Hawke, J.P.; Henk, W.G.; Thune, R.L. Photobacterium damselae subsp. piscicida is capable of replicating in hybrid striped bass macrophages. J. Aquat. Anim. Health 2003, 15, 175–183. [Google Scholar]
- Magariños, B.; Romalde, J.L.; Noya, M.; Barja, J.L.; Toranzo, A.E. Adherence and invasive capacities of the fish pathogen Pasteurella piscicida. FEMS Microbiol. Lett. 1996, 138, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Andreoni, F.; Boiani, R.; Serafini, G.; Amagliani, G.; Dominici, S.; Riccioni, G.; Zaccone, R.; Mancuso, M.; Scapigliati, G.; Magnani, M. Isolation of a novel gene from Photobacterium damselae subsp. piscicida and analysis of the recombinant antigen as promising vaccine candidate. Vaccine 2013, 31, 820–826. [Google Scholar] [PubMed]
- Magariños, B.; Romalde, J.L.; Lemos, M.L.; Barja, J.L.; Toranzo, A.E. Iron uptake by Pasteurella piscicida and its role in pathogenicity for fish. Appl. Environ. Microbiol. 1994, 60, 2990–2998. [Google Scholar] [PubMed]
- Do Vale, A.; Magariños, B.; Romalde, J.L.; Lemos, M.L.; Ellis, A.E.; Toranzo, A.E. Binding of haemin by the fish pathoghen Photobacterium damselae subsp. piscicida. Dis. Aquat. Organ. 2002, 48, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Juiz-Río, S.; Osorio, C.R.; Lemos, M.L. Heme uptake genes in human and fish isolates of Photobacterium damselae: Existence of hutA pseudogenes. Arch. Microbiol. 2005, 185, 347–358. [Google Scholar]
- Andreoni, F.; Boiani, R.; Serafini, G.; Bianconi, I.; Dominici, S.; Gorini, F.; Magnani, M. Expression, purification, and characterization of the recombinant putative periplasmic hemin-binding protein (hutB) of Photobacterium damselae subsp. piscicida. Biosci. Biotechnol. Biochem. 2009, 73, 1180–1183. [Google Scholar] [CrossRef] [PubMed]
- Osorio, C.R.; Rivas, A.J.; Balado, M.; Fuentes-Monteverde, J.C.; Rodríguez, J.; Jiménez, C.; Lemos, M.L.; Waldor, M.K. A transmissible plasmid-borne pathogenicity island confers piscibactin biosynthesis in the fish pathogen Photobacterium damselae subsp. piscicida. Appl. Environ. Microbiol. 2015, 81, 5867–5879. [Google Scholar] [CrossRef] [PubMed]
- Souto, A.; Montaos, M.A.; Rivas, A.J.; Balado, M.; Osorio, C.R.; Rodríguez, J.; Lemos, M.L.; Jiménez, C. Structure and biosynthetic assembly of Piscibactin, a new siderophore from Photobacterium damselae subsp. piscicida, predicted from genome analysis. Eur. J. Org. Chem. 2012, 29, 5693–5700. [Google Scholar] [CrossRef]
- Osorio, C.R.; Juiz-Río, S.; Lemos, M.L. A siderophore biosynthesis gene cluster from the fish pathogen Photobacterium damselae subsp. piscicida is structurally and functionally related to the Yersinia high-pathogenicity island. Microbiology 2006, 152, 3327–3341. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Díaz, J.A.; Fumanal, M.; Viguera, E.; Moriñigo, M.A.; Balebona, M.C. Use of in vivo induced technology to identify antigens expressed by Photobacterium damselae subsp. piscicida during infection of Senegalese sole (Solea senegalensis). Fish Shellfish Immunol. 2017, 64, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Goh, E.; Yim, G.; Tsui, W.; McClure, J.; Surette, M.G.; Davies, J. Transcriptional modulation of bacterial gene expression by subinhibitory concentrations of antibiotics. Proc. Natl. Acad. Sci. USA 2002, 99, 17025–17030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Song, J.; Huang, H.; Chen, W.; Li, M.; Zhao, Y.; Cong, Y.; Zhu, J.; Rao, X.; Hu, X.; et al. Identification of in-vivo induced genes of Streptococcus suis serotype 2 specially expressed in infected human. Microb. Pathog. 2013, 63, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.C.; Balebona, M.C.; Horne, M.T.; Ellis, A.E. Superoxide dismutase and catalase in Photobacterium damselae subsp. piscicida and their roles in resistance to reactive oxygen species. Microbiology 1999, 145, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rosales, P.; Chabrillon, M.; Morinigo, M.A.; Balebona, M.C. Survival against exogenous hydrogen peroxide of Photobacterium damselae subsp. piscicida under different culture conditions. J. Fish Dis. 2003, 26, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rosales, P.; Chabrillon, M.; Arijo, S.; Martinez-Manzanares, E.; Moriñigo, M.A.; Balebona, M.C. Superoxide dismutase and catalase activities in Photobacterium damselae ssp. piscicida. J. Fish Dis. 2006, 29, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Osorio, C.R.; Collins, M.D.; Toranzo, A.E.; Barja, J.L.; Romalde, J.L. 16S rRNA gene sequence analysis of Photobacterium damselae and nested PCR method for rapid detection of the causative agent of fish pasteurellosis. Appl. Environ. Microbiol. 1999, 65, 2942–2946. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Touati, D. Iron and oxidative stress in bacteria. Arch. Biochem. Biophys. 2000, 373, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 2011, 3, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Díaz, J.A.; Fumanal, M.; Mancera, J.M.; Moriñigo, M.A.; Balebona, M.C. Two routes of infection with Photobacterium damselae subsp. piscicida are effective in the modulation of the transcription of immune related genes in Solea senegalensis. Vet. Immunol. Immunopathol. 2016, 179, 8–17. [Google Scholar] [PubMed]
- Neves, J.V.; Wilson, J.M.; Rodrigues, P.N. Transferrin and ferritin response to bacterial infection: The role of the liver and brain in fish. Dev. Comp. Immunol. 2009, 33, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Mosca, F.; Ciulli, S.; Volpatti, D.; Romano, N.; Volpe, E.; Bulfon, C.; Massimini, M.; Caccia, E.; Galeotti, M.; Tiscar, P.G. Defensive response of European sea bass (Dicentrarchus labrax) against Listonella anguillarum or Photobacterium damselae subsp. piscicida experimental infection. Vet. Immunol. Immunopathol. 2014, 162, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Thode, S.; Rojeck, E.; Kozlowski, M.; Ahmad, R.; Haugen, P. Distribution of siderophore gene systems on a Vibrionaceae phylogeny: Database searches, phylogenetic analyses and evolutionary perspectives. PLoS ONE 2018, 13, e0191860. [Google Scholar] [CrossRef] [PubMed]
- Ernst, F.D.; Homuth, G.; Stoof, J.; Mader, U.; Waidner, B.; Kuipers, E.J.; Kist, M.; Kusters, J.G.; Bereswill, S.; van Vliet, A.H. Iron-responsive regulation of the Helicobacter pylori iron-cofactored superoxide dismutase SodB is mediated by Fur. J. Bacteriol. 2005, 187, 3687–3692. [Google Scholar] [CrossRef] [PubMed]
- Baillon, M.A.; van Vliet, A.H.; Ketley, J.M.; Constantinidou, C.; Penn, C.W. An iron-regulated alkyl hydroperoxide reductase (AhpC) confers aerotolerance and oxidative stress resistance to the microaerophilic pathogen Campylobacter jejuni. J. Bacteriol. 1999, 181, 4798–4804. [Google Scholar] [PubMed]
- Costa-Ramos, C.; Do Vale, A.; Ludovico, P.; Dos Santos, N.M.S.; Silva, M.T. The bacterial exotoxin AIP56 induces fish macrophage and neutrophil apoptosis using mechanisms of the extrinsic and intrinsic pathways. Fish Shellfish Immunol. 2011, 30, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Do Vale, A.; Marques, F.; Silva, M.T. Apoptosis of sea bass (Dicentrarchus labrax L.) neutrophils and macrophages induced by experimental infection with Photobacterium damselae subsp. piscicida. Fish Shellfish Immunol. 2003, 15, 129–144. [Google Scholar] [CrossRef]
- Rico-Perez, G.; Pezza, A.; Pucciarelli, M.G.; de Pedro, M.A; Soncini, F.C.; Garcia-Del Portillo, F. A novel peptidoglycan d,l-endopeptidase induced by Salmonella inside eukaryotic cells contributes to virulence. Mol. Microbiol. 2016, 99, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, G.; Lun, S.; Guo, H.; Ammerman, N.C.; Geiman, D.E.; Bishai, W.R. Rv2190c, an NlpC/P60 family protein, is required for full virulence of Mycobacterium tuberculosis. PLoS ONE 2012, 7, e43429. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Lingle, C.K.; Adam, P.R.; Ramyar, K.X.; McWhorter, W.J.; Stabel, J.R.; Picking, W.D.; Geisbrecht, B.V. NlpC/P60 domain-containing proteins of Mycobacterium avium subspecies paratuberculosis that differentially bind and hydrolyze peptidoglycan. Protein Sci. 2016, 25, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Pak, M.; Kish, R.; Kajihara, K.; Brown, E.J. A mycobacterial operon essential for virulence in vivo and invasion and intracellular persistence in macrophages. Infect. Immun. 2006, 74, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Elkamel, A.A.; Thune, R.L. Invasion and replication of Photobacterium damselae subsp. piscicida in fish cell lines. J. Aquat. Anim. Health 2003, 15, 167–174. [Google Scholar] [CrossRef]
- Osorio, C.R.; Juiz-Río, S.; Lemos, M.L. The ABC-transporter hutCD genes of Photobacterium damselae subsp. piscicida are essential for haem utilization as iron source and are expressed during infection in fish. J. Fish Dis. 2010, 33, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Kovacs-Simon, A.; Titball, R.W.; Michell, S.L. Lipoproteins of bacterial pathogens. Infect. Immun. 2011, 79, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, H.; Matsuyama, S. Sorting of lipoproteins to the outer membrane in E. coli. Biochim. Biophys. Acta 2004, 1693, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, T.; Raynal, B.; Munier-Lehmann, H. Two classes of bacterial IMPDHs according to their quaternary structures and catalytic properties. PLoS ONE 2015, 10, e0116578. [Google Scholar] [CrossRef]
- Chiu, Y.; Kuo, T.Y.; Lin, C.C.; Chen, W.J. Proteomic analysis reveals responsive proteins of Vibrio parahaemolyticus on exposure to cationic antimicrobial peptides. J. Appl. Microbiol. 2011, 110, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Keiler, K.C. Mechanisms of ribosome rescue in bacteria. Nat. Rev. Microbiol. 2015, 13, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Szebenyi, D.M.; Musayev, F.N.; di Salvo, M.L.; Safo, M.K.; Schirch, V. Serine hydroxymethyltransferase: Role of glu75 and evidence that serine is cleaved by a retroaldol mechanism. Biochemistry 2004, 43, 6868–6876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, K.; Duan, Z.; Zhou, J.; Yu, Z.; Ni, Y.; Lu, C. Identification and characterization of inosine 5-monophosphate dehydrogenase in Streptococcus suis type 2. Microb. Pathog. 2009, 47, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Merrell, D.S.; Hava, D.L.; Camilli, A. Identification of novel factors involved in colonization and acid tolerance of Vibrio cholerae. Mol. Microbiol. 2002, 43, 1471–1491. [Google Scholar] [CrossRef] [PubMed]
- Jelsbak, L.; Hartman, H.; Schroll, C.; Rosenkrantz, J.T.; Lemire, S.; Wallrodt, I.; Thomnsen, L.E.; Poolman, M.; Kilstrup, M.; Jensen, P.R.; et al. Identification of metabolic pathways essential for fitness of Salmonella typhimurium in vivo. PLoS ONE 2014, 9, e101869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, M.A.; Hayes, S.; Deadman, M.E.; Tang, C.M.; Hood, D.W.; Moxon, E.R. Signature tagged mutagenesis of Haemophilus infuenzae identifies genes required for in vivo survival. Microb. Pathog. 2002, 33, 211–223. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Code | Sequence (5’→3’) | Amplicon Size (bp) | Source |
|---|---|---|---|---|
| Apoptosis induced protein 56 kD | aip56 | GGTCGAAGCGATACAAGAGC (F) CCGTTGAAATCATCATCGTG (R) | 207 | This study |
| Adhesion lipoprotein | pdp-0080 | TGCAGGCCAACATCTAACAG (F) TTAGCTCAGCAGGGAATGGT (R) | 158 | This study |
| periplasmic hemin binding protein | hutB | ACGGAGCATCGTTCTCAACT (F) TGGCATTGTTTTGATGGTTG (R) | 264 | This study |
| ABC transporter ATPase | hutD | TGAACCCACATCTGCTCTTG (F) GCGGTTGGGGTTAGTACTTG (R) | 201 | This study |
| Protein 55 kD | p55 | GGATTTGGCTACCTCGTTCA (F) CCCACGGAGCATTAAACATT (R) | 249 | This study |
| Alkyl hydroperoxide reductase | ahpC | ATGGTGGTATTGGCCCTGTT (F) CATTGAGCTGGGCACACTTC (R) | 250 | [21] |
| Inosine-5’-monophosphate dehydrogenase | impdh | TGCTGATGGTGGTATCCGTT (F) GACATCGCACCAAGAGAACC (R) | 177 | [21] |
| Superoxide dismutase | sod | AGACGCACTAGAACCACACA (F) GGGCTTAGACAGTGCCAGTA (R) | 213 | [21] |
| Non-ribosomal peptide synthetase involved in siderophore biosynthesis 1 | irp1 | GCTACAGAGGCCGCTATTTG (F) CTTCATCTTGCCAGTAGCCA (R) | 202 | [21] |
| Non-ribosomal peptide synthetase involved in siderophore biosynthesis 2 | irp2 | AGGCAGCATTTCAGCAGATT (F) CGTTGTTCTCGGTCGGTATT (R) | 226 | [21] |
| Serine hydroxymethyl transferase | shmt | CGGAACTTTATGCAGCCATT (F) CAATGGCAAGTTGTTCTGCT (R) | 201 | [21] |
| Alanyl-trna synthethase | alars | GTGTTAAGCATGGGCGATTT (F) CCTTGTTCACCACAGAAGCA (R) | 232 | [21] |
| 16S ribosomal RNA | 16S rRNA | AACTGGCAGGCTAGAGTCTT (F) CACAACCTCCAAGTAGACAT (R) | 198 | [21] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez-Díaz, J.A.; Fumanal, M.; Do Vale, A.; Fernández-Díaz, C.; Moriñigo, M.Á.; Balebona, M.C. Transcription of IVIAT and Virulence Genes in Photobacterium damselae subsp. piscicida Infecting Solea senegalensis. Microorganisms 2018, 6, 67. https://doi.org/10.3390/microorganisms6030067
Núñez-Díaz JA, Fumanal M, Do Vale A, Fernández-Díaz C, Moriñigo MÁ, Balebona MC. Transcription of IVIAT and Virulence Genes in Photobacterium damselae subsp. piscicida Infecting Solea senegalensis. Microorganisms. 2018; 6(3):67. https://doi.org/10.3390/microorganisms6030067
Chicago/Turabian StyleNúñez-Díaz, José Alberto, Milena Fumanal, Ana Do Vale, Catalina Fernández-Díaz, Miguel Ángel Moriñigo, and María Carmen Balebona. 2018. "Transcription of IVIAT and Virulence Genes in Photobacterium damselae subsp. piscicida Infecting Solea senegalensis" Microorganisms 6, no. 3: 67. https://doi.org/10.3390/microorganisms6030067