Occurrence, Diversity of Listeria spp. Isolates from Food and Food-Contact Surfaces and the Presence of Virulence Genes

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. DNA Extraction
2.3. PCR Tests
2.4. Numerical Analysis of Genotypic Profiles
2.5. Sequencing and Sequence Analyses
3. Results
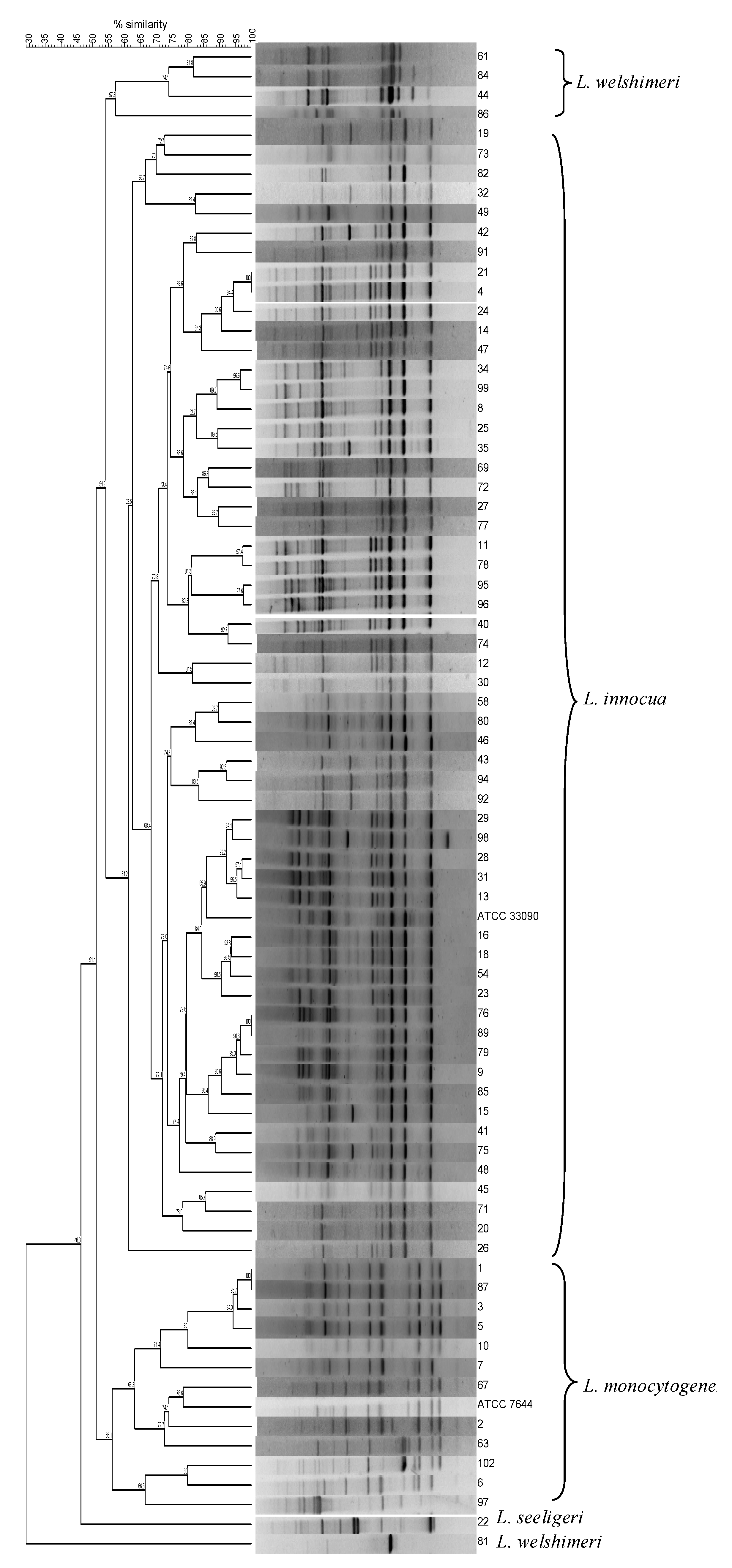
3.1. Typing and Identification of Listeria spp. Isolates
3.2. Pathogenic Potential of the Listeria spp. Isolates
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization/Food and Agriculture Organization (WHO/FAO). Technical Report: Risk As-Sessment of Listeria monocytogenes in Ready-to-Eat Foods; World Health Organization/Food and Agriculture Organization (WHO/FAO): Geneva, Switzerland, 2004; Available online: http://apps.who.int/iris/bitstream/10665/42875/1/9241562625.pdf (accessed on 7 January 2019).
- Pan, Y.; Breidt, F., Jr.; Kathariou, S. Resistance of Listeria monocytogenes biofilms to sanitizing agents in a simulated food processing environment. Appl. Environ. Microbiol. 2006, 72, 7711–7717. [Google Scholar] [CrossRef]
- Commission Regulation (EC). No 2073/2005 of 15 November 2005 on Microbiological Criteria for Food-Stuffs (Text with EEA Relevance); OJ L 338, 22.12.2005; Commission Regulation (EC), 2005; pp. 1–26. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32005R2073 (accessed on 7 December 2019).
- Guillet, C.; Join-Lambert, O.; Le Monnier, A.; Leclercq, A.; Mechaï, F.; Mamzer-Bruneel, M.F.; Bielecka, M.K.; Scortti, M.; Disson, O.; Berche, P.; et al. Human listeriosis caused by Listeria ivanovii. Emerg. Infect. Dis. 2010, 16, 136–138. [Google Scholar] [CrossRef]
- Rocourt, J.; Hof, H.; Schrettenbrunner, A.; Malinverni, R.; Bille, J. Acute purulent Listeria seelingeri meningitis in an immunocompetent adult. Schweiz. Med. Wochenschr. 1986, 116, 248–251. [Google Scholar] [PubMed]
- Rocha, P.R.; Dalmasso, A.; Grattarola, C.; Casalone, C.; Del Piero, F.; Bottero, M.T.; Capucchio, M.T. Atypical cerebral listeriosis associated with Listeria innocua in a beef bull. Res. Vet. Sci. 2013, 94, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Favaro, M.; Sarmati, L.; Sancesario, G. First case of Listeria innocua meningitis in a patient on steroids and eternecept. JMM Case Rep. 2014, 1, e003103. [Google Scholar] [CrossRef]
- Volokhov, D.V.; Duperrier, S.; Neverov, A.A.; George, J.; Buchrieser, C.; Hitchins, A.D. The presence of the internalin gene in natural atypically hemolytic Listeria innocua strains suggests descent from L. monocytogenes. Appl. Environ. Microbiol. 2007, 73, 1928–1939. [Google Scholar] [CrossRef]
- Moura, A.; Disson, O.; Lavina, M.; Thouvenot, P.; Huang, L.; Leclercq, A.; Fredriksson-Ahomaa, M.; Eshwar, A.K.; Stephan, R.; Lecuit, M. Atypical hemolytic Listeria innocua isolates are virulent, albeit less than Listeria monocytogenes. Infect. Immun. 2019, 87, e00758-18. [Google Scholar] [CrossRef]
- Clayton, E.M.; Daly, K.M.; Guinane, C.M.; Hill, C.; Cotter, P.D.; Ross, P.R. Atypical Listeria innocua strains possess an intact LIPI-3. BMC Microbiol. 2014, 14, 58. [Google Scholar] [CrossRef]
- Hadjilouka, A.; Andritsos, N.D.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Listeria monocytogenes serotype prevalence and biodiversity in diverse food products. J. Food Prot. 2014, 77, 2115–2120. [Google Scholar] [CrossRef]
- Chen, Y.; Knabel, S.J. Multiplex PCR for simultaneous detection of bacteria of the genus Listeria, Listeria monocytogenes, and major serotypes and epidemic clones of L. monocytogenes. Appl. Environ. Microbiol. 2007, 73, 6299–6304. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Ainsworth, A.J.; Austin, F.W.; Lawrence, M.L. Characterization of virulent and avirulent Listeria monocytogenes strains by PCR amplification of putative transcriptional regulator and internalin genes. J. Med. Microbiol. 2003, 52, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Curtis, T.D.; Takeuchi, I.; Gram, L.; Knudsen, G.M. The Influence of the toxin/Antitoxin mazEF on growth and survival of Listeria monocytogenes under stress. Toxins 2017, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, N.J.; Hermansson, M.; Wilén, B.M. The choice of PCR primers has great impact on assessments of bacterial community diversity and dynamics in a wastewater treatment plant. PLoS ONE 2013, 8, e76431. [Google Scholar] [CrossRef]
- Gouws, P.A.; Liedemann, I. Evaluation of diagnostic PCR for the detection of Listeria monocytogenes in food products. Food Technol. Biotechnol. 2005, 43, 201–205. [Google Scholar]
- Conficoni, D.; Santagiuliana, M.; Marchesan, M.; Franceschini, F.; Catellani, P.; Ferioli, M.; Giaccone, V. Distribution of Listeria spp. on carcasses of regularly slaughtered swine for Italian dry cured ham. J. Food Prot. 2019, 82, 1104–1109. [Google Scholar] [CrossRef]
- Angelidis, A.S.; Kalamaki, M.S.; Georgiadou, S.S. Identification of non-Listeria spp. bacterial isolates yielding a β-D-glucosidase-positive phenotype on Agar Listeria according to Ottaviani and Agosti (ALOA). Int. J. Food Microbiol. 2015, 193, 114–129. [Google Scholar] [CrossRef]
- Van Coillie, E.; Werbrouck, H.; Heyndrickx, M.; Herman, L.; Rijpens, N. Prevalence and typing of Listeria monocytogenes in ready-to-eat food products on the Belgian market. J. Food Prot. 2004, 67, 2480–2487. [Google Scholar] [CrossRef]
- Gilbreth, S.E.; Call, J.E.; Wallace, F.M.; Scott, V.N.; Chen, Y.; Luchansky, J.B. Relatedness of Listeria monocytogenes isolates recovered from selected ready-to-eat foods and listeriosis patients in the United States. Appl. Environ. Microbiol. 2005, 71, 8115–8122. [Google Scholar] [CrossRef][Green Version]
- Marini, E.; Magi, G.; Vincenzi, C.; Manso, E.; Facinelli, B. Ongoing outbreak of invasive listeriosis due to serotype 1/2a Listeria monocytogenes, Ancona province, Italy, January 2015 to February 2016. Eurosurveillance 2016, 21, 17. [Google Scholar] [CrossRef]
- Amato, E.; Filipello, V.; Gori, M.; Lomonaco, S.; Losio, M.N.; Parisi, A.; Huedo, P.; Knabel, S.J.; Pontello, M. Identification of a major Listeria monocytogenes outbreak clone linked to soft cheese in Northern Italy, 2009–2011. BMC. Infect. Dis. 2017, 17, 342. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority—European Centre for Disease Prevention and Control. Technical Report: Multi-Country Outbreak of Listeria Monocytogenes Sequence Type 8 Infections Linked to Con-Sumption of Salmon Products; EFSA Supporting Publication: Parma, Italy, 2018; p. EN1496. Available online: https://www.efsa.europa.eu/en/supporting/pub/en-1496 (accessed on 7 December 2019).
- Maurella, C.; Gallina, S.; Ru, G.; Adriano, D.; Bellio, A.; Bianchi, D.M.; Chiavacci, L.; Crescio, M.I.; Croce, M.; D’Errico, V.; et al. Outbreak of febrile gastroenteritis caused by Listeria monocytogenes 1/2a in sliced cold beef ham, Italy, May 2016. Eurosurveillance 2018, 23, 10. [Google Scholar] [CrossRef] [PubMed]
- Salata, C.; Lisotto, P.; Boldrin, C.; De Canale, E.; Piccirillo, A.; Calistri, A.; Palù, G. A first molecular characterization of Listeria monocytogenes isolates circulating in humans from 2009 to 2014 in the Italian Veneto region. New Microbiol. 2018, 41, 232–234. [Google Scholar] [PubMed]


{kind=link}
| Sample Category | Lab1 | Lab2 | Lab3 | |||
|---|---|---|---|---|---|---|
| n. Samples | Positive Samples/Isolates | n. Samples | Positive Samples/Isolates | n. Samples | Positive Samples/Isolates | |
| Bovine meat | 102 | L. innocua 8, 9, 11, 12, 13, 16, 18, 21, 23 (hly), 24, 26, 27; L. monocytogenes 10 | 57 | L. innocua 42, 43, 46 | 98 | L. innocua 79, 80, 82, 85, 89, 91, 92, 94, 95 (hly), 96, 98 (hly), 99; L. monocytogenes 87, 97, 102; L. welshimeri 81, 84, 86 (inlA, hly) |
| Pork meat | 31 | L. innocua 4; L. monocytogenes 1, 2, 3, 7 | 9 | 0 | 21 | 0 |
| Fresh fish | 26 | L. innocua 14, 15, 20 | 5 | 0 | 14 | 0 |
| Raw cured pork products | 16 | L. monocytogenes 5, 6 | 13 | L. welshimeri 44 | 14 | L. innocua 73, 75 |
| Frozen pizza | 14 | L. innocua 19, 25 | 13 | 0 | 8 | L. innocua 71, 72, 76, 77 |
| Pizza dough | 5 | 0 | 2 | L. innocua 32, 49 | 22 | L. innocua 78 |
| Frozen pasta | 4 | 0 | 11 | L. innocua 40, 41 | 8 | L. innocua 74 (inlA) |
| Roast peppers | 3 | 12 | L. innocua 34, 35 | 0 | 0 | |
| Swabs from cheese-making plants | 53 | L. innocua 28, 29, 30; L. seeligeri 22 (inlA) | 19 | L. innocua 31, 45 (hly), 47, 48 | 25 | 0 |
| Cheese-aging boards | 22 | 0 | 12 | L. innocua 54, 58, 69; L. monocytogenes 63, 67; L. welshimeri 61 | 14 | 0 |
| Primers | Sequence (5’→3’) | Usage | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| 27f 1492r | AGAGTTTGATCMTGGCTCAG TACGGYTACCTTGTTACGACTT | 16S rRNA gene amplification | 1494 | [16] |
| GTG5 | GTGGTGGTGGTGGTG | Rep-PCR genotyping | n.a. | [12] |
| 4bF 4bR | AGTGGACAATTGATTGGTGAA CATCCATCCCTTACTTTGGAC | identification of the L. monocytogenes serotype 4b | 597 | [13] |
| 12aF 12aR | GAGTAATTATGGCGCAACATC CCAATCGCGTGAATATCGG | identification of the L. monocytogenes serotype 1/2a | 724 | [13] |
| ECIF ECIR | AATAGAAATAAGCGGAAGTGT TTATTTCCTGTCGGCTTAG | identification of the L. monocytogenes epidemic clone ECI | 303 | [13] |
| ECIIF ECIIR | ATTATGCCAAGTGGTTACGGA ATCTGTTTGCGAGACCGTGTC | identification of the L. monocytogenes epidemic clone ECII | 889 | [13] |
| ECIIIF ECIIIR | TTGCTAATTCTGATGCGTTGG GCGCTAGGGAATAGTAAAGG | identification of the L. monocytogenes epidemic clone ECIII | 497 | [13] |
| hlyF hlyR | CATTAGTGGAAAGATGGAATG GTATCCTCCAGAGTGATCGA | detection of the L. monocytogenes hly gene | 730 | [17] |
| inlAF1 inlAR1 | TAACATCAGTCCCCTAGCAGGT TAGCCAACCTGTCACTATTGGA | detection of the L. monocytogenes inlA gene | 516 | [9] |
| lmo2821F lmo2821R | TGTAACCCCGCTTACACAGTT TTACGGCTGGATTGTCTGTG | detection of the L. monocytogenes inlJ gene | 611 | [14] |
| llsAFor llsARev | CGATTTCACAATGTGATAGGATG GCACATGCACCTCATAAC | detection of L. monocytogenes llsA gene | 280 | [11] |
| mazF_qPCR_Fw mazF_qPCR_Rv | ACGGCCTGTTCTCATCATTC CGTTGGCAATTTTGCTTTTT | detection of the Listeria spp. mazF gene | 103 | [15] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, F.; Amadoro, C.; Conficoni, D.; Giaccone, V.; Colavita, G. Occurrence, Diversity of Listeria spp. Isolates from Food and Food-Contact Surfaces and the Presence of Virulence Genes. Microorganisms 2020, 8, 294. https://doi.org/10.3390/microorganisms8020294
Rossi F, Amadoro C, Conficoni D, Giaccone V, Colavita G. Occurrence, Diversity of Listeria spp. Isolates from Food and Food-Contact Surfaces and the Presence of Virulence Genes. Microorganisms. 2020; 8(2):294. https://doi.org/10.3390/microorganisms8020294
Chicago/Turabian StyleRossi, Franca, Carmela Amadoro, Daniele Conficoni, Valerio Giaccone, and Giampaolo Colavita. 2020. "Occurrence, Diversity of Listeria spp. Isolates from Food and Food-Contact Surfaces and the Presence of Virulence Genes" Microorganisms 8, no. 2: 294. https://doi.org/10.3390/microorganisms8020294
APA StyleRossi, F., Amadoro, C., Conficoni, D., Giaccone, V., & Colavita, G. (2020). Occurrence, Diversity of Listeria spp. Isolates from Food and Food-Contact Surfaces and the Presence of Virulence Genes. Microorganisms, 8(2), 294. https://doi.org/10.3390/microorganisms8020294