

Examination of the Bactericidal and Fungicidal Activity of Bacillus amyloliquefaciens M Isolated from Spring Waters in Bulgaria
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Media
2.3. Cultivation of Bacillus amyloliquefaciens M
2.4. Examination of the Antimicrobial Activity of Bacillus amyloliquefaciens M
2.5. Examination of the Antiphytopathogenic Activity of Bacillus amyloliquefaciens M at Different Temperatures and pH
2.6. Thin Layer Chromatography (TLC)
2.7. Biochromatography
2.8. SDS-PAGE
2.9. Gel Chromatography
2.10. Determination of the Number of Viable Cells and Spores of Bacillus amyloliquefaciens M
2.11. Modeling of the Process Kinetics and Identification of the Parameters in the Logistic Curve Model
2.12. Processing of the Results
3. Results and Discussion
3.1. Antibacterial and Antifungal Activity of Bacillus amyloliquefaciens M
3.2. Determination of the Rf Value of the Antibiotic Substances Produced by B. amyloliquefaciens M
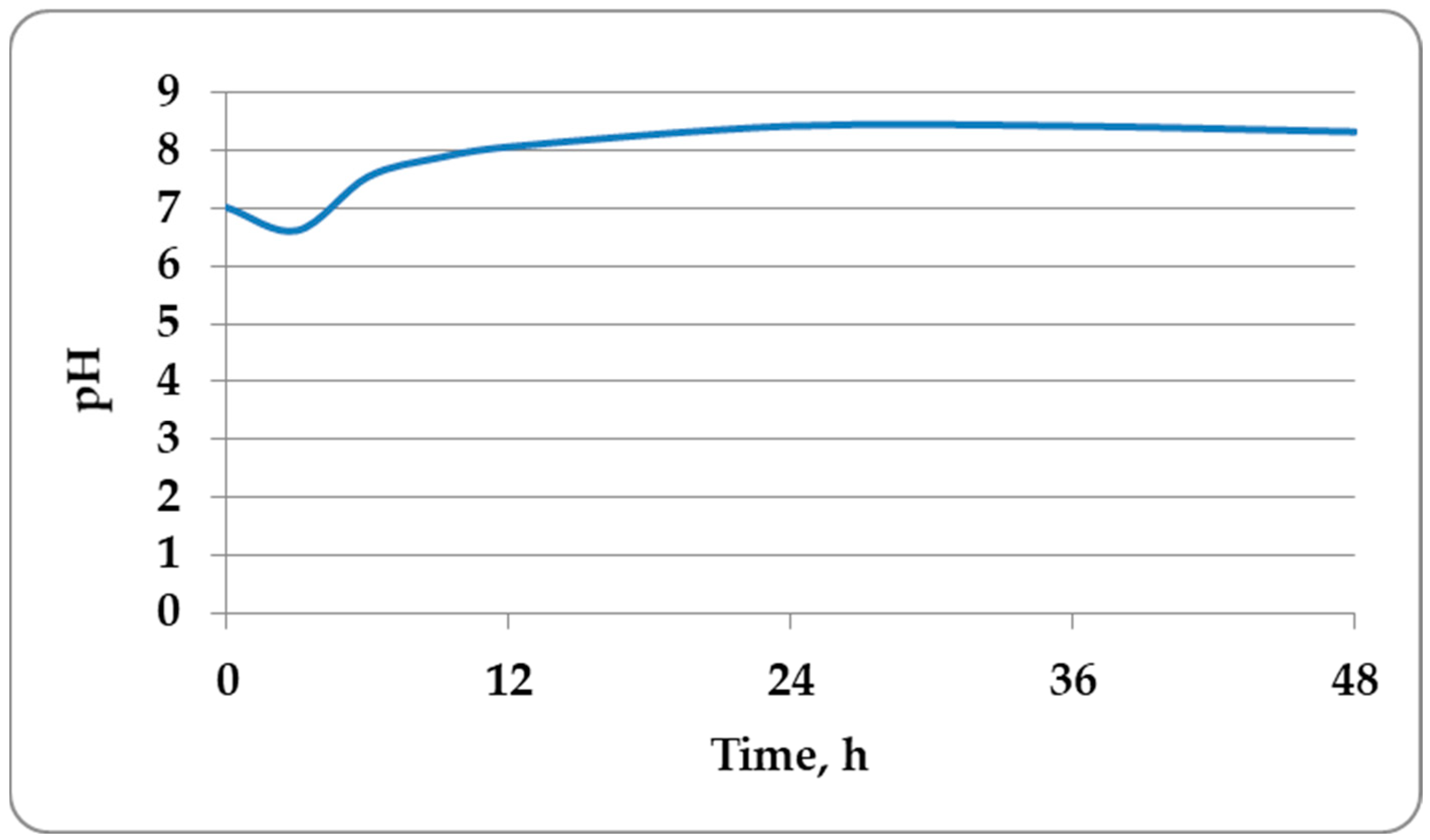
3.3. Cultivation of B. amyloliquefaciens M in a Laboratory Bioreactor and Determination of the Kinetic Parameters of the Process
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kheiri, F.; Faghani, M.; Landy, N. Evaluation of thyme and ajwain as antibiotic growth promoter substitutions on growth performance, carcass characteristics and serum biochemistry in Japanese quails (Coturnix japonica). Anim. Nutr. 2018, 4, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Gheisari, A.; Shahrvand, S.; Landy, N. Effect of ethanolic extract of propolis as an alternative to antibiotics as a growth promoter on broiler performance, serum biochemistry and immune responses. Vet. World 2017, 10, 249–254. [Google Scholar] [CrossRef]
- Andremont, A. Consequences of antibiotic therapy to the intestinal ecosystem. Ann. Françaises D‘Anesthésie Réanimation 2000, 19, 395–402. [Google Scholar] [CrossRef]
- Henning, S.; Marianne, S. Resistence to antibiotics in the normal flora of animals. Vet. Res. 2001, 32, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Denkova-Kostova, R.; Teneva, D.; Tomova, T.; Goranov, B.; Denkova, Z.; Shopska, V.; Slavchev, A.; Hristova-Ivanova, Y. Chemical composition, antioxidant and antimicrobial activity of essential oils from tangerine (Citrus reticulata L.), grapefruit (Citrus paradisi L.), lemon (Citrus lemon L.) and cinnamon (Cinnamomum zeylanicum Blume). Z. Naturforschung-Sect. C J. Biosci. 2021, 76, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Goranov, B.; Teneva, D.; Denkova-Kostova, R.; Shopska, V.; Oulahal, N.; Denkova, Z.; Kostov, G.; Degraeve, P.; Pagan, R. Biopreservation of chocolate mousse with Lactobacillus helveticus 2/20: Microbial Challenge Test. Molecules 2022, 27, 5631. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Working Group Report; Food and Health Agricultural Organisation of the United Nations—World Health Organisation: London, ON, Canada, 2002. [Google Scholar]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infect. Dis. 2008, 46, S58–S61. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Rodjan, P.; Soisuwan, K.; Thongprajukaew, K.; Theapparat, Y.; Khongthong, S.; Jeenkeawpieam, J.; Salaeharae, T. Effect of organic acids or probiotics alone or in combination on growth performance, nutrient digestibility, enzyme activities, intestinal morphology and gut microflora in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2018, 102, e931–e940. [Google Scholar] [CrossRef]
- Abudabos, A.M.; Alhouri, H.A.A.; Alhidary, I.A.; Nassan, M.A.; Swelum, A.A. Ameliorative effect of Bacillus subtilis, Saccharomyces boulardii, oregano, and calcium montmorillonite on growth, intestinal histology, and blood metabolites on Salmonella-infected broiler chicken. Environ. Sci. Pollut. Res. 2019, 26, 16274–16278. [Google Scholar] [CrossRef]
- Guo, M.; Li, M.; Zhang, C.; Zhang, X.; Wu, Y. Dietary administration of the Bacillus subtilis enhances immune responses and disease resistance in chickens. Front. Microbiol. 2020, 11, 1768. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, E. Antimicrobial Peptide and Plant Disease Control. FEMS Microbiol. Lett. 2007, 270, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; Pacheco, S.; Gómez, I.; Garcia-Gómez, B.; Onofre, J.; Soberón, M. Insecticidal Proteins from Bacillus thuringiensis and Their Mechanism of Action. In Bacillus thuringiensis and Lysinibacillus sphaericus; Springer International Publishing: Cham, Switzerland, 2017; pp. 53–66. [Google Scholar]
- Nayak, S.K. Multifaceted applications of probiotic Bacillus species in aquaculture with special reference to Bacillus subtilis. Rev. Fish. Sci. Aquac. 2020, 13, 862–906. [Google Scholar] [CrossRef]
- Jahan, R.; Siddique, S.S.; Jannat, R.; Hossain, M.M. Cosmos white rot: First characterization, physiology, host range, disease resistance, and chemical control. J. Basic Microbiol. 2022, 62, 911–929. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Akanda, A.M.; Hossain, M.M.; Hossain, M.M. First characterization of a newly emerging phytopathogen, Sclerotinia sclerotiorum causing white mold in pea. J. Basic Microbiol. 2021, 61, 923–939. [Google Scholar] [CrossRef] [PubMed]
- Sumi, C.D.; Yang, B.W.; Yeo, I.-C.; Hahm, Y.T. Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Can. J. Microbiol. 2015, 61, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and their mode of action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef]
- Yoo, J.-S.; Zheng, C.-J.; Lee, S.; Kwak, J.-H.; Kim, W.-G. Macrolactin N, a new peptide deformylase inhibitor produced by Bacillus subtilis. Bioorg. Med. Chem. Lett. 2006, 16, 4889–4892. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xue, Y.; Gao, W.; Li, J.; Zu, X.; Fu, D.; Bai, X.; Zuo, Y.; Hu, Z.; Zhang, F. Bacillaceae-derived peptide antibiotics since 2000. Peptides 2018, 101, 10–16. [Google Scholar] [CrossRef]
- Malviya, D.; Sahu, P.K.; Singh, U.B.; Paul, S.; Gupta, A.; Gupta, A.R.; Singh, S.; Kumar, M.; Paul, D.; Rai, J.P.; et al. Lesson from ecotoxity: Revisiting the microbial lipopeptides for the management of emerging diseases for crop protection. Int. J. Environ. Res. Public Health 2020, 17, 1434. [Google Scholar] [CrossRef]
- Stefanello, C.; Vieira, S.L.; Rios, H.V.; Simões, C.T.; Ferzola, P.H.; Sorbara, J.O.B.; Cowieson, A.J. Effects of energy, α-amylase, and β-xylanase on growth performance of broiler chickens. Anim. Feed Sci. Technol. 2017, 225, 205–212. [Google Scholar] [CrossRef]
- Sinnelä, M.T.; Park, Y.K.; Lee, J.H.; Jeong, K.C.C.; Kim, Y.-W.; Hwang, H.-J.; Mah, J.-H. Effects of Calcium and Manganese on Sporulation of Bacillus Species Involved in Food Poisoning and Spoilage. Foods 2019, 8, 119. [Google Scholar] [CrossRef]
- Baharudin, M.M.A.; Ngalimat, M.S.; Shariff, F.M.; Yusof, Z.N.B.; Karim, M.; Baharum, S.N.; Sabri, S. Antimicrobial activities of Bacillus velezensis strains isolated from stingless bee products against methicillin-resistant Staphylococcus aureus. PLoS ONE 2021, 16, e0251514. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 435128. [Google Scholar] [CrossRef]
- Leifert, C.; Li, H.; Chidburee, S.; Hampson, S.; Workman, S.; Sigee, D.; Epton, H.A.S.; Harbour, A. Antibiotic production and biocontrol activity by Bacillus subtilis CL27 and Bacillus pumilus CL45. J. Appl. Bacteriol. 1995, 78, 97–108. [Google Scholar] [CrossRef]
- Arx, E.V.; Faupel, M.; Brugger, M. Das 4,4′-Tetramethyldiamino-diphenylmethan Reagens (TDM) Eine Modifikation der Chlor-o-Tolidin Farbereaktion für die Dünnschichtchromatographie. J. Chromatogr. 1976, 120, 224–228. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Aitken, A.; Learmonth, P.M. The Protein Protocols Handbook, 2nd ed.; Walker, J.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2002. [Google Scholar]
- Massawe, V.C.; Hanif, A.; Farzand, A.; Mburu, D.K.; Ochola, S.O.; Wu, L.; Samad Tahir, H.A.; Gu, Q.; Wu, H.; Gao, X. Volatile compounds of endophytic Bacillus spp. have biocontrol activity against Sclerotinia sclerotiorum. Phytopathology 2018, 108, 1373–1385. [Google Scholar] [CrossRef]
- Birnie, C.; Malamud, D.; Schnaare, R. Antimicrobial evaluation of N-Alkyl Betaines and N-Alkyl-N,N-Dimethylamine oxides with variations in chain length. Antimicrob. Agents Chemother. 2000, 44, 2514–2517. [Google Scholar] [CrossRef]
- Martins, C.; Correia, V.G.; Aguiar-Ricardo, A.; Cunha, Â.; Moutinho, M.G.M. Antimicrobial activity of new green-functionalized oxazoline-based oligomers against clinical isolates. Springer Plus 2015, 4, 382. [Google Scholar] [CrossRef]
- Soliman, S.A.; Khaleil, M.M.; Metwally, R.A. Evaluation of the Antifungal Activity of Bacillus amyloliquefaciens and B. velezensis and characterization of the bioactive secondary metabolites produced against plant pathogenic fungi. Biology 2022, 11, 1390. [Google Scholar] [CrossRef]
- Shobier, A.H.S.; Abdel Ghani, S.A.; Barakat, K.M. GC/MS spectroscopic approach and antifungal potential of bioactive extracts produced by marine macroalgae. Egypt. J. Aquat. Res. 2016, 42, 289–299. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Senthilkumar, A.; Venkatesalu, V. Antibacterial and antifungal efficacy of fatty acid methyl esters from the leaves of Sesuvium portulacastrum L. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 775–780. [Google Scholar]
- Mensah-Agyei, G.O.; Ayeni, K.I.; Ezeamagu, C.O. GC-MS analysis of bioactive compounds and evaluation of antimicrobial activity of the extracts of Daedalea elegans: A Nigerian mushroom. Afr. J. Microbiol. Res. 2020, 14, 204–210. [Google Scholar] [CrossRef]
- Shanga, B.M.; Lyon, G.D. Bacillus subtilis BS107as an antagonist in potato blackieg and soft rod bacteria. Can. J. Microbiol. 1998, 44, 777–783. [Google Scholar]
- McKeen, C.D.; Reilly, C.C.; Pusey, P.L. Production and Partial Characterization of Antifungal Substances Antagonistic to Monilinia fructicola from Bacillus subtilis. Phytopathology 1986, 76, 136–139. [Google Scholar] [CrossRef]
- Besson, F.; Michel, G. Influence of divalent ions on the solubility of iturin and bacillomycin L, antifungal peptidolipids of Bacillus subtilis. Microbios 1991, 65, 15–21. [Google Scholar]
- Zhao, R.; Wang, H.-H.; Gao, J.; Zhang, W.-J.; Li, X.; Zhou, J.-J.; Liang, P.; Gao, X.-Y.; Gu, S.-H. Plant volatile compound methyl benzoate is highly effective against Spodoptera frugiperda and safe to non-target organisms as an eco-friendly botanical-insecticide. Ecotoxicol. Environ. Saf. 2022, 245, 114101. [Google Scholar] [CrossRef]
- Tzeng, Y.-M.; Rao, Y.K.; Tsay, K.-J.; Wu, W.-S. Effect of cultivation conditions on spore production from Bacillus amyloliquefaciens B128 and its antagonism to Botrytis elliptica. J. Appl. Microbiol. 2008, 104, 1275–1282. [Google Scholar] [CrossRef]
- Díaz-García, A.; García-Riaño, J.; Zapata-Narvaez, J. Improvement of sporulation conditions of a new strain of Bacillus amyloliquefaciens in liquid fermentation. Adv. Biosci. Biotechnol. 2015, 6, 302–310. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candida utilis ATCC 42402, 2 × 107 cfu/cm3 | Saccharomyces cerevisiae ATCC 9763, 2.4 × 107 cfu/cm3 | Aspergillus niger ATCC 1015, 1.2 × 107 cfu/cm3 | Aspergillus flavus ATCC 9643, 2.8 × 107 cfu/cm3 | Fusarium moniliforme ATCC 38932, 1.0 × 106 cfu/cm3 | Penicillium chrysogenum ATCC 28089, 2 × 107 cfu/cm3 | |
|---|---|---|---|---|---|---|
| Fermentation medium with molasses | ||||||
| B | 37.33 ± 0.47 | 39.50 ± 0.41 | 25.33 ± 0.47 | 40.50 ± 0.41 | 20.67 ± 0.47 | 32 17 ± 0.24 |
| ASN | 39.83 ± 0.24 | 25.83 ± 0.24 | 20.50 ± 0.41 | 30.33 ± 0.47 | 18.17 ± 0.24 | 28.33 ± 0.47 |
| Fermentation medium with malt | ||||||
| B | 30.50 ± 0.41 | 25.50 ± 0.41 | 30.50 ± 0.41 | 30.67 ± 0.47 | 18.17 ± 0.24 | 20.50 ± 0.41 |
| ASN | 26.33 ± 0.47 | 27.67 ± 0.47 | 20.83 ± 0.24 | 32.33 ± 0.47 | 20.33 ± 0.47 | 18.67 ± 0.47 |
| Fermentation medium MPB with salts | ||||||
| B | 37.33 ± 0.47 | 30.17 ± 0.24 | 22.33 ± 0.47 | 37.83 ± 0.24 | 20.17 ± 0.24 | 15.33 ± 0.47 |
| ASN | 32.50 ± 0.41 | 24.17 ± 0.24 | 22.50 ± 0.41 | 25.33 ± 0.47 | 12.67 ± 0.47 | 12.83 ± 0.24 |
| Bacillus amyloliquefaciens M | dinhibition zone, mm | |
|---|---|---|
| Sclerotinia sclerotium var scleroria ATCC 18687, 5.4 × 107 cfu/cm3 | ||
| 24 h | 72 h | |
| B | - | 25.33 ± 0.47 |
| ASN | 27.17 ± 0.24 | 27.50 ± 0.41 |
| CS | 30.33 ± 0.47 | 30.67 ± 0.47 |
| Escherichia coli ATCC 25922, 9 × 1010 cfu/cm3 | Salmonella enterica ssp. enterica serovar Enteritidis ATCC 25928, 2.5 × 1011 cfu/cm3 | Salmonella abony NTCC 6017, 5.4 × 1011 cfu/cm3 | Staphylococcus aureus ATCC 6538P, 3 × 1011 cfu/cm3 | Pseudomonas aeruginosa NBIMCC 1390, 1.4 × 1011 cfu/cm3 | Listeria monocytogenes ATCC 19111, 2.0 × 1010 cfu/cm3 | Bacillus cereus ATCC 14579, 5 × 109 cfu/cm3 | |
|---|---|---|---|---|---|---|---|
| Fermentation medium with molasses | |||||||
| B | 18.33 ± 0.47 | - | 9.50 ± 0.41 | 26.50 ± 0.41 | 20.17 ± 0.24 | 32.50 ± 0.41 | 18.50 ± 0.41 |
| ASN | 18.17 ± 0.24 | - | 9.33 ± 0.47 | 23.17 ± 0.24 | 18.17 ± 0.24 | 28.17 ± 0.24 | 20.50 ± 0.41 |
| Fermentation medium with malt | |||||||
| B | 18.33 ± 0.47 | - | 9.17 ± 0.24 | 15.50 ± 0.41 | 20.17 ± 0.24 | 33.33 ± 0.47 | 13.50 ± 0.41 |
| ASN | 20.67 ± 0.47 | - | 9.33 ± 0.47 | 28.17 ± 0.24 | 28.33 ± 0.47 | 26.67 ± 0.47 | 17.17 ± 0.24 |
| Fermentation medium MPB with salts | |||||||
| B | 12.17 ± 0.24 | - | 9.17 ± 0.24 | 25.33 ± 0.47 | 20.50 ± 0.41 | 20.33 ± 0.47 | 13.17 ± 0.24 |
| ASN | 10.33 ± 0.47 | - | - | 12.33 ± 0.47 | 9.17 ± 0.24 | 18.67 ± 0.47 | 17.33 ± 0.47 |
| Bacillus amyloliquefaciens M Static Cultivation 30 °C 48 h | Test Microorganisms | |||
|---|---|---|---|---|
| Aspergillus niger ATCC 1015, 3.0 × 105 cfu/cm3 | Aspergillus awamori ATCC 22342, 1.2 × 105 cfu/cm3 | Aspergillus flavus ATCC 9643, 1.1 × 105 cfu/cm3 | ||
| LBG broth pH = 4.5 | ASN | 23.17 ± 0.24 | 10.17 ± 0.24 | 10.17 ± 0.24 |
| CS | 23.33 ± 0.47 | 12.33 ± 0.47 | 10.33 ± 0.47 | |
| B | 26.33 ± 0.47 | 10.17 ± 0.24 | 10.33 ± 0.47 | |
| LBG broth pH = 5 | ASN | 25.67 ± 0.47 | 40.67 ± 0.47 | 25.17 ± 0.24 |
| CS | 28.33 ± 0.47 | 20.50 ± 0.41 | 24.17 ± 0.24 | |
| B | 28.17 ± 0.24 | 30.33 ± 0.47 | 28.50 ± 0.41 | |
| LBG broth pH = 6 | ASN | 28.17 ± 0.24 | 37.67 ± 0.47 | 34.17 ± 0.24 |
| CS | 30.33 ± 0.47 | 23.33 ± 0.47 | 32.17 ± 0.24 | |
| B | 31.67 ± 0.47 | 32.50 ± 0.41 | 35.17 ± 0.24 | |
| LBG broth pH = 7 | ASN | 37.67 ± 0.47 | 35.33 ± 0.47 | 34.67 ± 0.47 |
| CS | 25.33 ± 0.47 | 25.50 ± 0.41 | 22.33 ± 0.47 | |
| B | 32.17 ± 0.24 | 43.67 ± 0.47 | 36.50 ± 0.41 | |
| LBG broth pH = 8 | ASN | 38.33 ± 0.47 | 40.67 ± 0.47 | 34.33 ± 0.47 |
| CS | 31.17 ± 0.24 | 30.50 ± 0.41 | 25.67 ± 0.47 | |
| B | 32.33 ± 0.47 | 42.17 ± 0.24 | 35.17 ± 0.24 | |
| MPB broth pH = 4.5 | ASN | 12.17 ± 0.24 | 12.50 ± 0.41 | 20.33 ± 0.47 |
| CS | 15.33 ± 0.47 | 10.17 ± 0.24 | 12.67 ± 0.47 | |
| B | 13.17 ± 0.24 | 13.33 ± 0.47 | 20.50 ± 0.41 | |
| MPB broth pH = 5 | ASN | 23.67 ± 0.47 | 10.17 ± 0.24 | 18.33 ± 0.47 |
| CS | 13.33 ± 0.47 | 10.17 ± 0.24 | 22.17 ± 0.24 | |
| B | 27.50 ± 0.41 | 15.33 ± 0.47 | 22.50 ± 0.41 | |
| MPB broth pH = 6 | ASN | 28.67 ± 0.47 | 25.50 ± 0.41 | 23.17 ± 0.24 |
| CS | 35.67 ± 0.47 | 33.50 ± 0.41 | 25.17 ± 0.24 | |
| B | 30.33 ± 0.47 | 30.33 ± 0.47 | 28.33 ± 0.47 | |
| MPB broth pH = 7 | ASN | 30.50 ± 0.41 | 12.17 ± 0.24 | 22.33 ± 0.47 |
| CS | 28.17 ± 0.24 | 10.17 ± 0.24 | 12.17 ± 0.24 | |
| B | 32.33 ± 0.47 | 14.50 ± 0.41 | 26.67 ± 0.47 | |
| MPB broth pH = 8 | ASN | 20.17 ± 0.24 | 12.17 ± 0.24 | 15.17 ± 0.24 |
| CS | 25.33 ± 0.47 | 13.50 ± 0.41 | 14.33 ± 0.47 | |
| B | 25.50 ± 0.41 | 14.67 ± 0.47 | 13.17 ± 0.24 | |
| Bacillus amyloliquefaciens M Dynamic Cultivation 30 °C 48 h | Test Microorganisms | |||
|---|---|---|---|---|
| Aspergillus niger ATCC 1015, 3.0 × 105 cfu/cm3 | Aspergillus awamori ATCC 22342, 1.2 × 105 cfu/cm3 | Aspergillus flavus ATCC 9643, 1.1 × 105 cfu/cm3 | ||
| LBG broth pH = 4.5 | ASN | 30.17 ± 0.24 | 12.17 ± 0.24 | 13.17 ± 0.24 |
| CS | 25.50 ± 0.41 | 12.33 ± 0.47 | 10.33 ± 0.47 | |
| B | 27.33 ± 0.47 | 12.33 ± 0.47 | 15.33 ± 0.47 | |
| LBG broth pH = 5 | ASN | 28.33 ± 0.47 | 33.33 ± 0.47 | 34.50 ± 0.41 |
| CS | 35.17 ± 0.24 | 40.67 ± 0.47 | 20.17 ± 0.24 | |
| B | 30.50 ± 0.41 | 32.50 ± 0.41 | 32.67 ± 0.47 | |
| LBG broth pH = 6 | ASN | 25.33 ± 0.47 | 33.17 ± 0.24 | 35.17 ± 0.24 |
| CS | 32.17 ± 0.24 | 28.67 ± 0.47 | 35.17 ± 0.24 | |
| B | 34.50 ± 0.41 | 33.33 ± 0.47 | 35.33 ± 0.47 | |
| LBG broth pH = 7 | ASN | 42.67 ± 0.47 | 32.33 ± 0.47 | 30.17 ± 0.24 |
| CS | 30.17 ± 0.24 | 23.50 ± 0.41 | 23.17 ± 0.24 | |
| B | 43.17 ± 0.24 | 37.50 ± 0.41 | 35.33 ± 0.47 | |
| LBG broth pH = 8 | ASN | 43.33 ± 0.47 | 38.17 ± 0.24 | 35.17 ± 0.24 |
| CS | 34.50 ± 0.41 | 37.17 ± 0.24 | 30.50 ± 0.41 | |
| B | 43.50 ± 0.41 | 43.67 ± 0.47 | 37.50 ± 0.41 | |
| MPB broth pH = 4.5 | ASN | 22.33 ± 0.47 | 22.67 ± 0.47 | 20.17 ± 0.24 |
| CS | 12.17 ± 0.24 | 10.50 ± 0.41 | 15.17 ± 0.24 | |
| B | 20.33 ± 0.47 | 20.67 ± 0.47 | 27.33 ± 0.47 | |
| MPB broth pH = 5 | ASN | 20.33 ± 0.47 | 13.17 ± 0.24 | 13.17 ± 0.24 |
| CS | 15.17 ± 0.24 | 14.33 ± 0.47 | 18.33 ± 0.47 | |
| B | 20.50 ± 0.41 | 15.67 ± 0.47 | 20.50 ± 0.41 | |
| MPB broth pH = 6 | ASN | 25.33 ± 0.47 | 24.50 ± 0.41 | 28.17 ± 0.24 |
| CS | 35.33 ± 0.47 | 30.17 ± 0.24 | 30.50 ± 0.41 | |
| B | 28.33 ± 0.47 | 30.67 ± 0.47 | 30.67 ± 0.47 | |
| MPB broth pH = 7 | ASN | 20.17 ± 0.24 | 15.17 ± 0.24 | 28.17 ± 0.24 |
| CS | 22.33 ± 0.47 | 15.50 ± 0.41 | 25.33 ± 0.47 | |
| B | 28.67 ± 0.47 | 18.33 ± 0.47 | 28.50 ± 0.41 | |
| MPB broth pH = 8 | ASN | 18.17 ± 0.24 | 15.33 ± 0.47 | 25.17 ± 0.24 |
| CS | 15.17 ± 0.24 | 17.67 ± 0.47 | 20.67 ± 0.47 | |
| B | 23.50 ± 0.41 | 25.50 ± 0.41 | 25.33 ± 0.47 | |
| Bacillus amyloliquefaciens M Static Cultivation 37 °C 24 h | Test Microorganisms | |||
|---|---|---|---|---|
| Aspergillus niger ATCC 1015, 3 × 105 cfu/cm3 | Aspergillus awamori ATCC 22342, 1.2 × 105 cfu/cm3 | Aspergillus flavus ATCC 9643, 1.12 × 105 cfu/cm3 | ||
| LBG broth pH = 4.5 | ASN | 18.17 ± 0.24 | 10.17 ± 0.24 | 25.33 ± 0.47 |
| CS | 20.33 ± 0.47 | 12.17 ± 0.24 | 25.67 ± 0.47 | |
| B | 15.50 ± 0.41 | 15.50 ± 0.41 | 30.17 ± 0.24 | |
| LBG broth pH = 5 | ASN | 30.50 ± 0.41 | 30.50 ± 0.41 | 37.17 ± 0.24 |
| CS | 32.33 ± 0.47 | 34.50 ± 0.41 | 37.33 ± 0.47 | |
| B | 32.67 ± 0.47 | 35.33 ± 0.47 | 35.50 ± 0.41 | |
| LBG broth pH = 6 | ASN | 35.33 ± 0.47 | 28.17 ± 0.24 | 35.17 ± 0.24 |
| CS | 40.67 ± 0.47 | 32.50 ± 0.41 | 40.33 ± 0.47 | |
| B | 40.33 ± 0.47 | 33.33 ± 0.47 | 43.33 ± 0.47 | |
| LBG broth pH = 7 | ASN | 30.50 ± 0.41 | 30.17 ± 0.24 | 37.67 ± 0.47 |
| CS | 35.17 ± 0.24 | 32.33 ± 0.47 | 37.50 ± 0.41 | |
| B | 37.33 ± 0.47 | 43.33 ± 0.47 | 43.50 ± 0.41 | |
| LBG broth pH = 8 | ASN | 35.17 ± 0.24 | 35.50 ± 0.41 | 37.50 ± 0.41 |
| CS | 37.67 ± 0.47 | 40.17 ± 0.24 | 35.33 ± 0.47 | |
| B | 34.33 ± 0.47 | 37.17 ± 0.24 | 40.67 ± 0.47 | |
| MPB broth pH = 4.5 | ASN | 30.17 ± 0.24 | 34.33 ± 0.47 | 35.33 ± 0.47 |
| CS | 37.17 ± 0.24 | 32.17 ± 0.24 | 34.67 ± 0.47 | |
| B | 32.33 ± 0.47 | 34.67 ± 0.47 | 39.17 ± 0.24 | |
| MPB broth pH = 5 | ASN | 30.33 ± 0.47 | 34.33 ± 0.47 | 35.17 ± 0.24 |
| CS | 37.17 ± 0.24 | 40.17 ± 0.24 | 34.67 ± 0.47 | |
| B | 37.50 ± 0.41 | 35.50 ± 0.41 | 37.33 ± 0.47 | |
| MPB broth pH = 6 | ASN | 32.17 ± 0.24 | 32.33 ± 0.47 | 40.50 ± 0.41 |
| CS | 35.17 ± 0.24 | 35.67 ± 0.47 | 34.17 ± 0.24 | |
| B | 37.50 ± 0.41 | 34.33 ± 0.47 | 35.33 ± 0.47 | |
| MPB broth pH = 7 | ASN | 35.33 ± 0.47 | 30.17 ± 0.24 | 34.33 ± 0.47 |
| CS | 33.67 ± 0.47 | 39.50 ± 0.41 | 25.67 ± 0.47 | |
| B | 35.17 ± 0.24 | 39.50 ± 0.41 | 32.67 ± 0.47 | |
| MPB broth pH = 8 | ASN | 30.17 ± 0.24 | 30.67 ± 0.47 | 42.33 ± 0.47 |
| CS | 30.50 ± 0.41 | 37.17 ± 0.24 | 35.67 ± 0.47 | |
| B | 39.33 ± 0.47 | 37.50 ± 0.41 | 37.50 ± 0.41 | |
| Bacillus amyloliquefaciens M Dynamic Cultivation 37 °C 24 h | Test Microorganisms | |||
|---|---|---|---|---|
| Aspergillus niger ATCC 1015, 3 × 105 cfu/cm3 | Aspergillus awamori ATCC 22342, 1.2 × 105 cfu/cm3 | Aspergillus flavus ATCC 9643, 1.12 × 105 cfu/cm3 | ||
| LBG broth pH = 4.5 | ASN | 15.17 ± 0.24 | 15.17 ± 0.24 | 28.50 ± 0.41 |
| CS | 25.33 ± 0.47 | 20.50 ± 0.41 | 37.17 ± 0.24 | |
| B | 22.33 ± 0.47 | 17.50 ± 0.41 | 30.33 ± 0.47 | |
| LBG broth pH = 5 | ASN | 43.67 ± 0.47 | 35.17 ± 0.24 | 42.67 ± 0.47 |
| CS | 32.33 ± 0.47 | 35.50 ± 0.41 | 32.17 ± 0.24 | |
| B | 28.17 ± 0.24 | 33.33 ± 0.47 | 35.50 ± 0.41 | |
| LBG broth pH = 6 | ASN | 32.33 ± 0.47 | 26.17 ± 0.24 | 43.17 ± 0.24 |
| CS | 45.33 ± 0.47 | 43.33 ± 0.47 | 52.17 ± 0.24 | |
| B | 43.67 ± 0.47 | 35.50 ± 0.41 | 40.33 ± 0.47 | |
| LBG broth pH = 7 | ASN | 27.17 ± 0.24 | 33.67 ± 0.47 | 40.33 ± 0.47 |
| CS | 40.50 ± 0.41 | 40.17 ± 0.24 | 49.17 ± 0.24 | |
| B | 38.33 ± 0.47 | 37.33 ± 0.47 | 45.50 ± 0.41 | |
| LBG broth pH = 8 | ASN | 32.33 ± 0.47 | 40.50 ± 0.41 | 43.17 ± 0.24 |
| CS | 43.17 ± 0.24 | 42.33 ± 0.47 | 45.33 ± 0.47 | |
| B | 35.17 ± 0.24 | 39.17 ± 0.24 | 45.33 ± 0.47 | |
| MPB broth pH = 4.5 | ASN | 28.17 ± 0.24 | 30.17 ± 0.24 | 35.50 ± 0.41 |
| CS | 27.33 ± 0.47 | 32.17 ± 0.24 | 40.33 ± 0.47 | |
| B | 37.67 ± 0.47 | 37.50 ± 0.41 | 37.17 ± 0.24 | |
| MPB broth pH = 5 | ASN | 40.17 ± 0.24 | 37.50 ± 0.41 | 40.17 ± 0.24 |
| CS | 32.33 ± 0.47 | 42.33 ± 0.47 | 35.17 ± 0.24 | |
| B | 43.17 ± 0.24 | 30.50 ± 0.41 | 40.17 ± 0.24 | |
| MPB broth pH = 6 | ASN | 35.33 ± 0.47 | 34.50 ± 0.41 | 37.33 ± 0.47 |
| CS | 45.17 ± 0.24 | 43.50 ± 0.41 | 45.67 ± 0.47 | |
| B | 43.33 ± 0.47 | 41.17 ± 0.24 | 40.17 ± 0.24 | |
| MPB broth pH = 7 | ASN | 25.17 ± 0.24 | 25.17 ± 0.24 | 25.17 ± 0.24 |
| CS | 35.33 ± 0.47 | 32.67 ± 0.47 | 32.33 ± 0.47 | |
| B | 35.33 ± 0.47 | 30.50 ± 0.41 | 35.50 ± 0.41 | |
| MPB broth pH = 8 | ASN | 30.17 ± 0.24 | 28.17 ± 0.24 | 35.17 ± 0.24 |
| CS | 37.50 ± 0.41 | 37.33 ± 0.47 | 40.33 ± 0.47 | |
| B | 37.33 ± 0.47 | 32.33 ± 0.47 | 44.50 ± 0.41 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goranov, B.; Gaytanska, Y.; Denkova-Kostova, R.; Ivanova, P.; Denkova, Z.; Kostov, G. Examination of the Bactericidal and Fungicidal Activity of Bacillus amyloliquefaciens M Isolated from Spring Waters in Bulgaria. Appl. Sci. 2024, 14, 3612. https://doi.org/10.3390/app14093612
Goranov B, Gaytanska Y, Denkova-Kostova R, Ivanova P, Denkova Z, Kostov G. Examination of the Bactericidal and Fungicidal Activity of Bacillus amyloliquefaciens M Isolated from Spring Waters in Bulgaria. Applied Sciences. 2024; 14(9):3612. https://doi.org/10.3390/app14093612
Chicago/Turabian StyleGoranov, Bogdan, Yordanka Gaytanska, Rositsa Denkova-Kostova, Petya Ivanova, Zapryana Denkova, and Georgi Kostov. 2024. "Examination of the Bactericidal and Fungicidal Activity of Bacillus amyloliquefaciens M Isolated from Spring Waters in Bulgaria" Applied Sciences 14, no. 9: 3612. https://doi.org/10.3390/app14093612