Ion Channel Properties of a Cation Channelrhodopsin, Gt_CCR4

1
Department of Life Science and Applied Chemistry, Nagoya Institute of Technology, Showa-ku, Nagoya 466-8555, Japan
2
OptoBio Technology Research Center, Nagoya Institute of Technology, Showa-ku, Nagoya 466-8555, Japan
3
PRESTO, Japan Science and Technology Agency, 4-1-8 Honcho, Kawaguchi, Saitama 332-0012, Japan
*
Author to whom correspondence should be addressed.
Appl. Sci. 2019, 9(17), 3440; https://doi.org/10.3390/app9173440
Submission received: 23 July 2019
/
Revised: 16 August 2019
/
Accepted: 18 August 2019
/
Published: 21 August 2019
(This article belongs to the Special Issue The Advances and Applications of Optogenetics)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:We previously reported a cation channelrhodopsin, Gt_CCR4, which is one of the 44 types of microbial rhodopsins from a cryptophyte flagellate, Guillardia theta. Due to the modest homology of amino acid sequences with a chlorophyte channelrhodopsin such as Cr_ChR2 from Chlamydomonas reinhardtii, it has been proposed that a family of cryptophyte channelrhodopsin, including Gt_CCR4, has a distinct molecular mechanism for channel gating and ion permeation. In this study, we compared the photocurrent properties, cation selectivity and kinetics between well-known Cr_ChR2 and Gt_CCR4 by a conventional path clamp method. Large and stable light-induced cation conduction by Gt_CCR4 at the maximum absorbing wavelength (530 nm) was observed with only small inactivation (15%), whereas the photocurrent of Cr_ChR2 exhibited significant inactivation (50%) and desensitization. The light sensitivity of Gt_CCR4 was higher (EC50 = 0.13 mW/mm2) than that of Cr_ChR2 (EC50 = 0.80 mW/mm2) while the channel open life time (photocycle speed) was in the same range as that of Cr_ChR2 (25~30 ms for Gt_CCR4 and 10~15 ms for Cr_ChR2). This observation implies that Gt_CCR4 enables optical neuronal spiking with weak light in high temporal resolution when applied in neuroscience. Furthermore, we demonstrated high Na+ selectivity of Gt_CCR4 in which the selectivity ratio for Na+ was 37-fold larger than that for Cr_ChR2, which primarily conducts H+. On the other hand, Gt_CCR4 conducted almost no H+ and no Ca2+ under physiological conditions. These results suggest that ion selectivity in Gt_CCR4 is distinct from that in Cr_ChR2. In addition, a unique red-absorbing and stable intermediate in the photocycle was observed, indicating a photochromic property of Gt_CCR4.
1. Introduction
Microbial-type rhodopsins are made up of seven or eight transmembrane helices with a covalently bound all-trans retinal as the chromophore [1]. They are found in archaea, bacteria, eukaryota (such as fungi and algae) and viruses, and are physiologically responsible for energy production and the phototaxis reaction. Molecular functions of microbial rhodopsin involve ion transporters, sensors and light-regulated enzymes. As for ion-transporting rhodopsins, they are divided into ion-pumps and channels. Bacteriorhodopsin (BR) was the first identified outward directed proton-pumping rhodopsin [2]. The discovery of a Cl− pump, an Na+ pump and inward-directed proton pumps has been even until recently [3,4,5,6]. Structure-based and spectroscopic studies, when combined with electrophysiology and molecular dynamics studies, revealed the detailed molecular mechanism of bacteriorhodopsin and other pumps.
Channelrhodopsin-1 and -2 (Cr_ChR1 and Cr_ChR2) from Chlamydomonas reinhardtii were the first light-gated ion channels to be discovered [7,8]. These homologous proteins permeate cations in which the permeability ratio of H+, Na+, and K+ is 106, 1, and 0.5, respectively. High-resolution X-ray structures revealed details of their molecular architecture and provided insight into their photoactivation and ion conduction [9,10].
Successful expression of Cr_ChR2 in neurons allowed the action potential to be manipulated by light, which opened up a new field of research, optogenetics [11,12]. A number of variant molecules have been engineered to improve the functionality of ChR, and homologous ChRs were then reported [13]. Color-tuning variants cover almost the entire visible range. Cr_ChR2 displays an action spectrum maximum at 470 nm [8]. ChR variants such as C1V1, which is the chimeric version of ChR1 from Chlamydomonas reinhardtii and Volvox carteri, or C1C2 (a green receiver) absorb light at around 530~545 nm [14,15,16]. Another red-shifted ChR, Chrimson from Chlamydomonas noctigama, exhibits an absorption maximum at 590 nm which allows reliable neuronal stimulation by light exceeds 600 nm [17]. On the other hand, TsChR or PsChR absorb a shorter wavelength, making it possible to excite neurons at 440 nm [18].
The lifetime of an open channel can be extended by mutations at C128 and D156 (DC pair) which form a hydrogen bond bridge in Cr_ChR2. Mutations at C128 to Thr, Ala and Ser slowed the kinetics of channel closing 200, 5000 and 10,000-fold respectively [19]. Cr_ChR2 D156C displayed an even stronger effect, namely higher light sensitivity and prolonged lifetime of the open channel, by as much as 30 min [20].
Converting ion selectivity is challenging. The Cr_ChR2 L132C mutant showed improved Ca2+ permeability [21]. The permeability ratio between H+ and Na+ could be modified by a replacement at E143 to A in Chrimson [22]. Anion channelrhodopsins were engineered or discovered from nature and they have been applied as neuronal silencing tools [23,24,25]. The crystal structures of anion channelrhodopsins revealed their unique features related to the channel gating mechanism [26,27,28].
A novel cation channelrhodopsin family was reported in 2016 and 2017 from Guillardia theta, namely Gt_CCR1-4 [29,30]. These cation channelrhodopsins (CCRs) from cryptophyte algae are more homologous to haloarchaeal rhodopsins, such as proton pumping bacteriorhodopsin, than to chlorophyte CCRs, including Cr_ChR2. Actually, Gt_CCRs conserve the characteristic amino-acid residues involved in unidirectional proton transfer, including the proton acceptor D85 and the proton donor D96 in bacteriorhodopsin (Table S1).
On the other hand, a characteristic glutamic acid in TM2 (E90 in Cr_ChR2) which is crucial for channel gating and ion selectivity, is not conserved in Gt_CCRs [23,31]. Cr_ChR2 possesses a so-called DC pair (C128 and D156 in Cr_ChR2), which is responsible for the channel life time [19,32,33]. This is not found in Gt_CCRs. Thus, overall sequence patterns separate these cryptophyte CCRs form chlorophyte channels. The molecular mechanisms such as channel gating mechanism and ion selectivity could be distinct in chlorophyte CCRs. Sineshchekov and coworkers already revealed that the retinal Schiff-base (SB) in Gt_CCR2 rapidly deprotonates to the D85 homolog, as in BR, upon photoisomerization [34]. Channel-opening requires deprotonation of the D96 homolog. We independently identified photocycle intermediates during the channel function of Gt_CCR4 from electrophysiological and flash photolysis experiments. The M-decay corresponds to channel-closing, implicating tight coupling between retinal dynamics and channel function. However, reprotonation of SB for channel closing was achieved by the direct return of a proton from the D85 homolog. Such proton transfers are not the case with Cr_ChR2. In Cr_ChR2, D156 in TM4 provides the proton [35]. We demonstrated, using an FTIR study, that the secondary structural change in the primary reaction was much smaller than in Cr_ChR2 [30]. These differences in the molecular mechanism place the cryptophyte CCR in a new family of channelrhodopsins, which we described as “DTD channelrhodopsins” or “BR-like cation channelrhodopsins” [29,30]. To further reveal the characteristics of these DTD channelrhodopsins, in this study we performed electrophysiological measurements in parallel with Cr_ChR2.
2. Materials and Methods
2.1. Expression Plasmids
2.2. Cell Culture
The electrophysiological assays of Gt_CCR4 and Cr_ChR2 were performed on ND7/23 cells, which are hybrid cell lines derived from neonatal rat dorsal root ganglia neurons fused with mouse neuroblastoma [36]. ND7/23 cells were grown on a collagen-coated coverslip in Dulbecco’s modified Eagle’s medium (Wako, Osaka, Japan) supplemented with 2.0 µM all-trans retinal and 5% fetal bovine serum, and under a 5% CO2 atmosphere at 37 °C. The expression plasmids were transiently transfected by using the FuGENE HD transfection Reagent (Promega, Fitchburg, WI, USA) according to the manufacturer’s instructions. Electrophysiological recordings were then conducted 24–36 h after transfection. Successfully transfected cells were identified by eGFP or Venus fluorescence under a microscope prior to the measurements.
2.3. Electrophysiology
All experiments were carried out at room temperature (22 ± 2 °C). Photocurrents were recorded as previously described using an Axopatch 200B amplifier (Molecular Devices, Sunnyvale, CA, USA) under a whole-cell patch clamp configuration [12]. Data were filtered at 5 kHz and sampled at 20 kHz (Digdata1550, Molecular Devices, Sunnyvale, CA, USA) and stored in a computer (pClamp10.6, Molecular Devices). Pipette resistance was 3–6 MΩ. The standard internal pipette solution for the whole-cell voltage clamp contained (in mM) 120 KOH, 100 glutamate, 2.5 MgCl2, 2.5 MgATP, 0.01 Alexa568, 50 HEPES, and 5 EGTA, and adjusted to pH 7.2. The standard extracellular solution for the whole-cell voltage clamp contained (in mM) 140 NaCl, 2 KCl, 2 MgCl2, 2 CaCl2, and 10 HEPES, and adjusted to pH 7.2. The ion selectivity internal pipette solution for the whole-cell voltage clamp contained (in mM) 1 NaCl, 1 KCl, 2 CaCl2, 2 MgCl2, 110 N-methyl D-glucamine, 10 CHES, and 10 EGTA, and adjusted to pH 9.0. The ion selectivity extracellular solution for the whole-cell voltage clamp contained (in mM) ExNMG, 9.0 is 1 NaCl, 1 KCl, 2 CaCl2, 2 MgCl2, 140 N-methyl D-glucamine, and 10 CHES, and adjusted to pH 9.0. ExNMG, 6.85 is 1 NaCl, 1 KCl, 2 CaCl2, 2 MgCl2, 140 N-methyl D-glucamine, and 10 MES, and adjusted to pH 6.85. ExNaCl, 9.0 is 140 NaCl, 1 KCl, 2 CaCl2, 2 MgCl2, and 10 CHES, and adjusted to pH 9.0. ExKCl, 9.0 is 1 NaCl, 140 KCl, 2 CaCl2, 2 MgCl2, and 10 CHES, and adjusted to pH 9.0. ExCsCl, 9.0 is 1 NaCl, 1 KCl, 140 CsCl, 2 CaCl2, 2 MgCl2, and 10 CHES, and adjusted to pH 9.0. ExCaCl2, 9.0 is 1 NaCl, 1 KCl, 70 CaCl2, 2 MgCl2, and 10 CHES, and adjusted to pH 9.0. ExMgCl2, 9.0 is 1 NaCl, 1 KCl, 2 CaCl2, 70 MgCl2, and 10 CHES, and adjusted to pH 9.0. All solutions of pH were adjusted with N-methyl D-glucamine or HCl. The liquid junction potential was calculated and compensated by pClamp 10.6 software. Time constants were determined by a single exponential fit unless noted.
2.4. Optics
For whole-cell voltage clamp, irradiation at 470 or 530 or 590 nm was carried out using WheeLED and collimated LED (parts No. WLS-LED-0530-03 or LCS-0530-03-22, WLS-LED-0590-03 Mightex, Toronto, ON, Canada) or an ND YAG flash laser, Mini lite at 532 nm (Continuum, San Jose, CA, USA) controlled by computer software (pCLAMP10.6, Molecular Devices). Light power was measured directly by an objective lens of a microscope by a power meter (LP1, Sanwa Electric Instruments Co., Ltd., Tokyo, Japan).
2.5. Confocal Images
Cell images shown in Figure 1A,B were observed by a Nikon A1 LFOV through an objective lens, Apo 60x Oil λS DIC N2.
2.6. Statistical Analysis
All data in the text and figures are expressed as mean ± SEM.
3. Results
3.1. Basic Characterization of Photocurrent
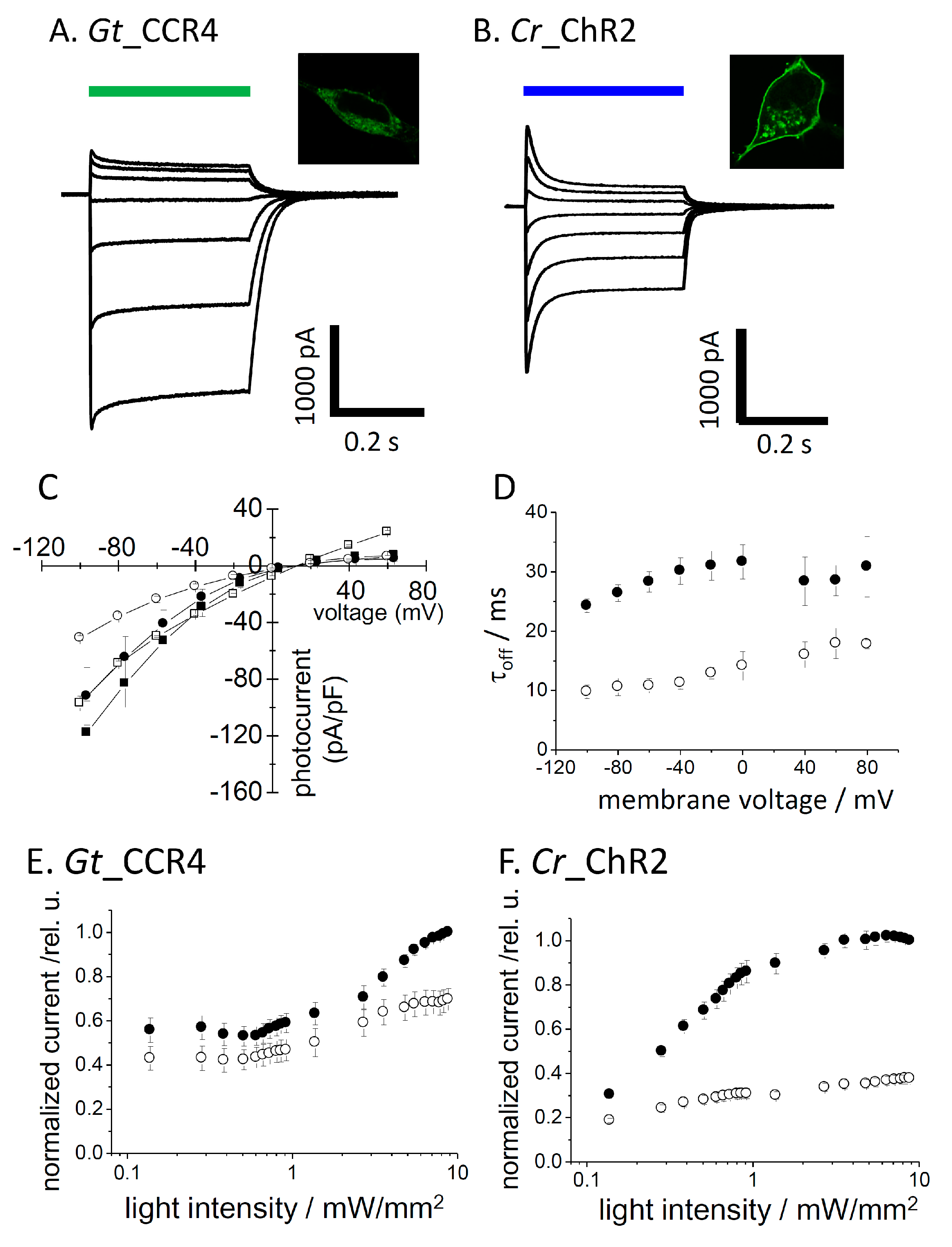
We transiently expressed Gt_CCR4 and Cr_ChR2 in ND7/23 cells by a conventional transfection method (FuGENE). Expression of these channels was visualized by tagged-GFP or Venus fluorescence. Strong membrane expression was confirmed for both channels, although cytoplasmic GFP was observed in Gt_CCR4-expressing cells (Figure 1A microscope images). We illuminated 530 nm light to induce a photocurrent for Gt_CCR4 and 470 nm light for Cr_ChR2 at the same light intensity (6.8 mW/mm2). A large photocurrent was recorded from the Gt_CCR4-expressing cells, reproducing previous studies (Figure 1A, left) [30]. Current amplitude reached −2 nA at −60 mV. The current showed an initial peak component (Ip) which decayed slightly into a steady state level (Iss). However, amplitude of the steady state still retained about 80% of the transient peak component. The photocurrent from Cr_ChR2 showed a large peak component which reached about −2 nA at −60 mV (Figure 1A, right). The current decayed by 50% of the initial peak, suggesting that Cr_ChR2 exhibits a markedly large inactivation compared to Gt_CCR4. Figure 1C depicts the current-voltage relationship of photocurrent from Gt_CCR4 and Cr_ChR2. Both peak component (Ip) and steady state current (Iss) are plotted. The shape of the I-V plot from Gt_CCR4 indicates strong inward-rectification. Iss of Cr_ChR2 displayed similarly inward-rectification, while Ip was weakly rectified, in which a markedly outward-directed current was observed at positive membrane voltages. For both Ip and Iss, Gt_CCR4 showed a larger current density (pA/pF) than Cr_ChR2. For example, the photocurrent (Iss) of Gt_CCR4 at −100 mV exceeded −80 pA/pF, while that of Cr_ChR2 was only about −40 pA/pF.
Kinetics in the photocurrent decay after shutting off the light is shown in Figure 1D. The time constant of Gt_CCR4 is about 25–35 ms under a membrane voltage between −100 and 80 mV, while Cr_ChR2 showed faster kinetics by about 10–20 ms. Next, we compared light sensitivity in the photocurrent of two channels (Figure 1E,F). The photocurrent amplitude from Cr_ChR2 grows as a typical sigmoidal curve for both the initial peak and the steady state components (Figure 1F). EC50 was determined as 0.8 mW/mm2 for Ip and 0.35 mW/mm2 for Iss under the conditions tested. On the other hand, the Gt_CCR4 current showed unique growth in terms of power dependency with two apparent phases in which the current first saturated at 0.1 mW/mm2 at about 50% of full activation, followed by the second phase of growth from 1 to 10 mW/mm2 (Figure 1E). The EC50 was determined as 0.13 mW/mm2 for Ip and 0.18 mW/mm2 for Iss. These results indicate that Gt_CCR4 is more sensitive to light with respect to channel activation.
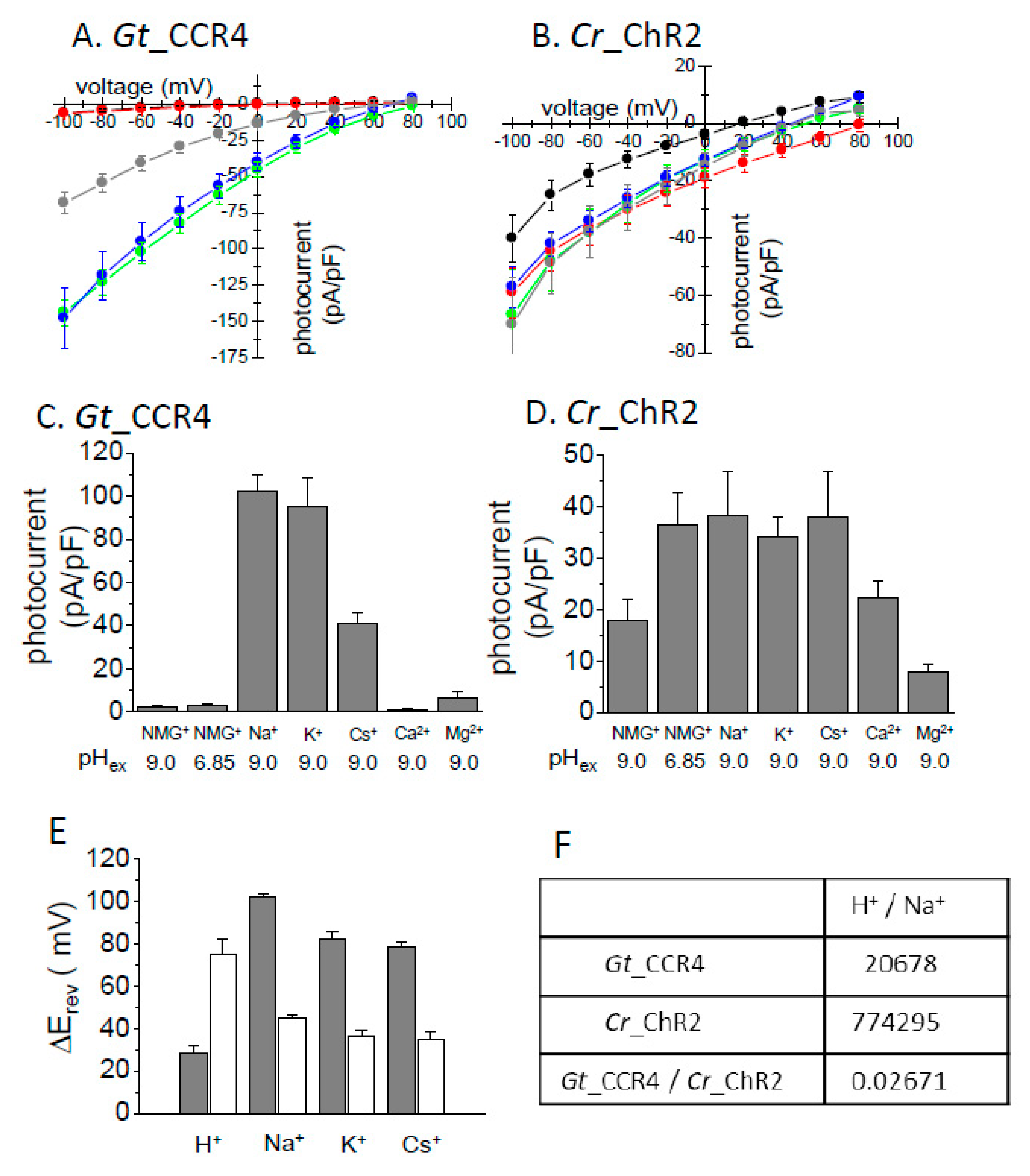
3.2. Ion Selectivity
It was already reported that Gt_CCRs are H+- and Na+-permeable cation channels [29,30]. We here investigated the cation selectivity of Gt_CCR4 in more detail relative to Cr_ChR2. Ionic conditions of the extracellular solution were systematically exchanged with various cations including Na+, K+, Cs+, Ca2+, Mg2+ and NMG. In the presence of NMG, one can assume H+ as the permeated ion. Figure 2A,B show the I-V plot of Gt_CCR4 and Cr_ChR2 under several ionic conditions. Obviously, the reversal potential shift in Na+ solution is larger in Gt_CCR4 than in Cr_ChR2, suggesting that Gt_CCR4 is more permeable to Na+ than Cr_ChR2. Notably, I-V plots of Gt_CCR4 at pH 6.85 and 9.0 in NMG are almost identical (Figure 2A) whereas a large shift of reversal potential was observed in Cr_ChR2 under the same conditions (Figure 2B). These results indicate that Gt_CCR4 has less H+ selectivity than Cr_ChR2. The photocurrent amplitude of Gt_CCR4 and Cr_ChR2 at −60 mV under each condition is summarized in Figure 2C,D. The current amplitude of Gt_CCR4 was significantly larger in the presence of Na+ and K+ close to −100 pA/pF and in the presence of Cs+ at about −40 pA/pF. In contrast, only a negligible current was observed at low pH, or in the presence of Ca2+ or Mg2+. This supports the notion that Gt_CCR4 is more of a monovalent metal cation selective channel. In addition, we also tested measurements under a competitive environment in which both Na+ and Ca2+ were both added to bath solutions (Figure S1). Interestingly, photocurrents by Gt_CCR4 were suppressed at a higher Ca2+ concentration (40 mM), suggesting that Na+ flow is blocked by Ca2+. In contrast, such a large difference in current amplitude was not observed under various conditions in the photocurrent from Cr_ChR2 (Figure 2D). This is due to low ion selectivity in Cr_ChR2 as was reported previously. To assume the selectivity ratio, the reversal potential shift from the condition with NMG at pH 9.0 is depicted in Figure 2E. The shifts (ΔErev) in Gt_CCR4 are larger than in Cr_ChR2 for Na+, K+, and Cs+ indicating that Gt_CCR4 is selective for monovalent cations but less selective for H+. In the initial study of Cr_ChR2, the permeability ratio was calculated based on the current amplitude [8]. The table in Figure 2F summarizes the ratio both for Gt_CCR4 and Cr_ChR2. H+ permeability for Cr_ChR2 is 0.77 × 105, close to the reported value (1 × 106), while that for Gt_CCR4 is 2.1 × 104, indicating about 37-fold less permeability for H+ in Gt_CCR4.
3.3. Flash Laser Electrophysiology
We then measured the photocurrent under a single-turnover condition with a flash laser as the light source (ND-YAG). As shown in Figure 3A,B, 5 ns light evoked a large inward-directed peak current at −60 mV for both Gt_CCR4- and Cr_ChR2-expressing cells. The current amplitude and direction are voltage-dependent for both channels, as was expected from recordings with LED. Current growth was fitted with a single exponential function (Figure 3C). The time constant of Gt_CCR4 seems to be independent of membrane voltage, while that of Cr_ChR2 slowed down slightly as voltage increased. Current decay was determined as about 20 ms for Gt_CCR4 and about 5–10 ms for Cr_ChR2, both of which are smaller than the value obtained from the measurement by LED light (Figure 1C), suggesting two distinct open states with different kinetics.
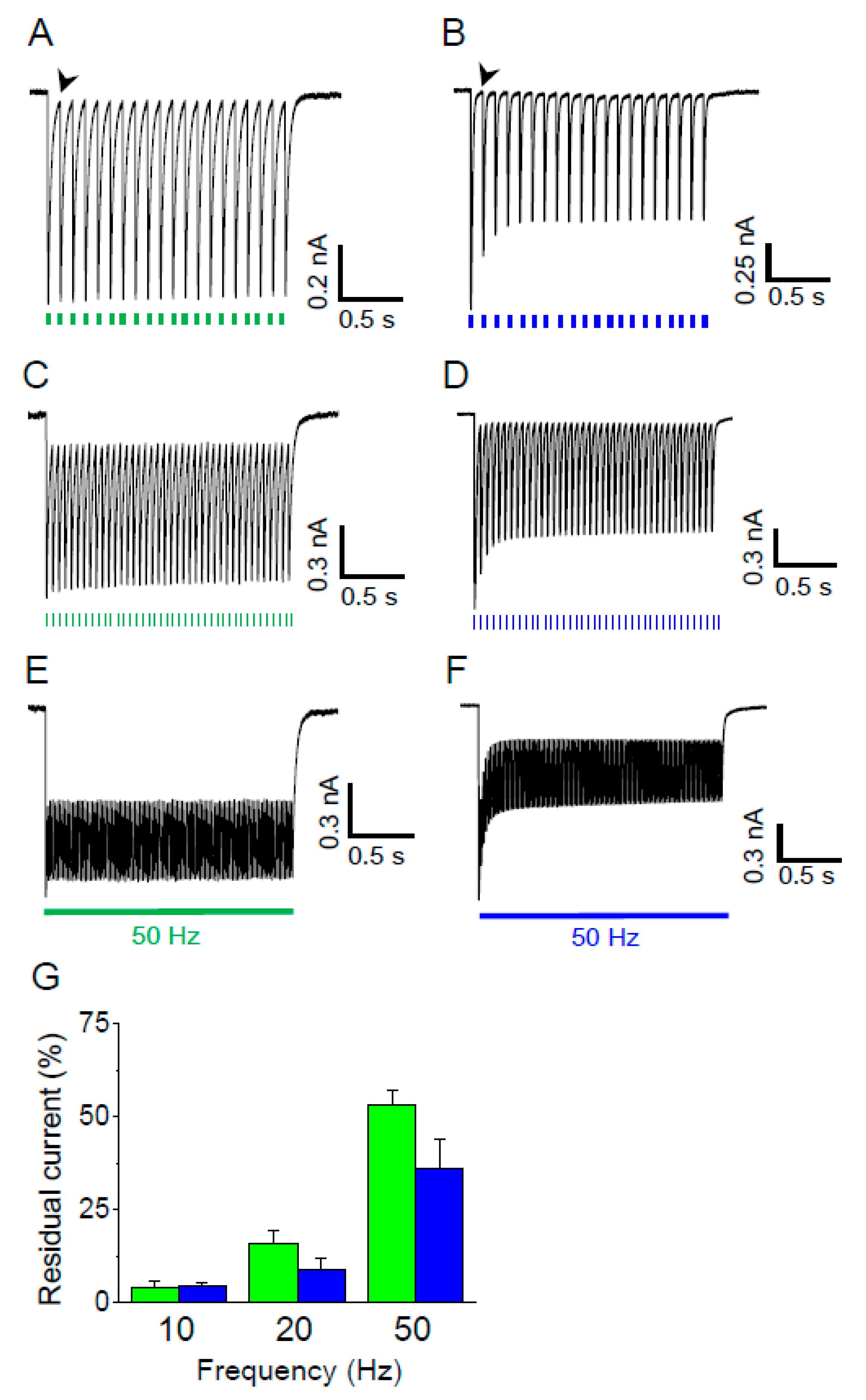
3.4. High Frequency Stimulation
For optogenetics application, reliable neuronal activation would require large and stable activity of the light-gated ion channel. In addition, rapid channel-closing is desired for optical stimulation at a high frequency. Thus, we compared the photocurrent from Gt_CCR4- and Cr_ChR2-expressing cells with three different light frequencies. As shown in Figure 4A, activation of Gt_CCR4 at 10 Hz light at 9.67 mW/mm2 generated high temporal peak currents which almost fully decayed before the next illumination. Peak amplitude remained unchanged because of a small inactivation, as shown in Figure 1A. On the other hand, peak current amplitude immediately decayed less after the initial stimulation in Cr_ChR2-expressing cells (Figure 4B). Stimulation of Gt_CCR4 at 20 and 50 Hz still retained a high level of peak amplitudes, although each peak did not decay completely (Figure 4C,E). Current inactivation was observed in Cr_ChR2-expressing cells at each frequency (20 and 50 Hz) (Figure 4D,F). Figure 4G summarizes the residual current level before the next stimulation at each light frequency. At 10 Hz, both Gt_CCR4 and Cr_ChR2 showed a very low level of the current level, indicating that channels were almost shut off. As frequency increased to 20 Hz and 50 Hz, significant residual currents were observed which exceeded 50% in Gt_CCR4 at 50 Hz whereas Cr_ChR2 showed a slightly lower residual current, probably because of faster channel kinetics.
3.5. Gt_CCR4 Is Inactivated by 590 nm Light But Fully Reactivated by Blue Light
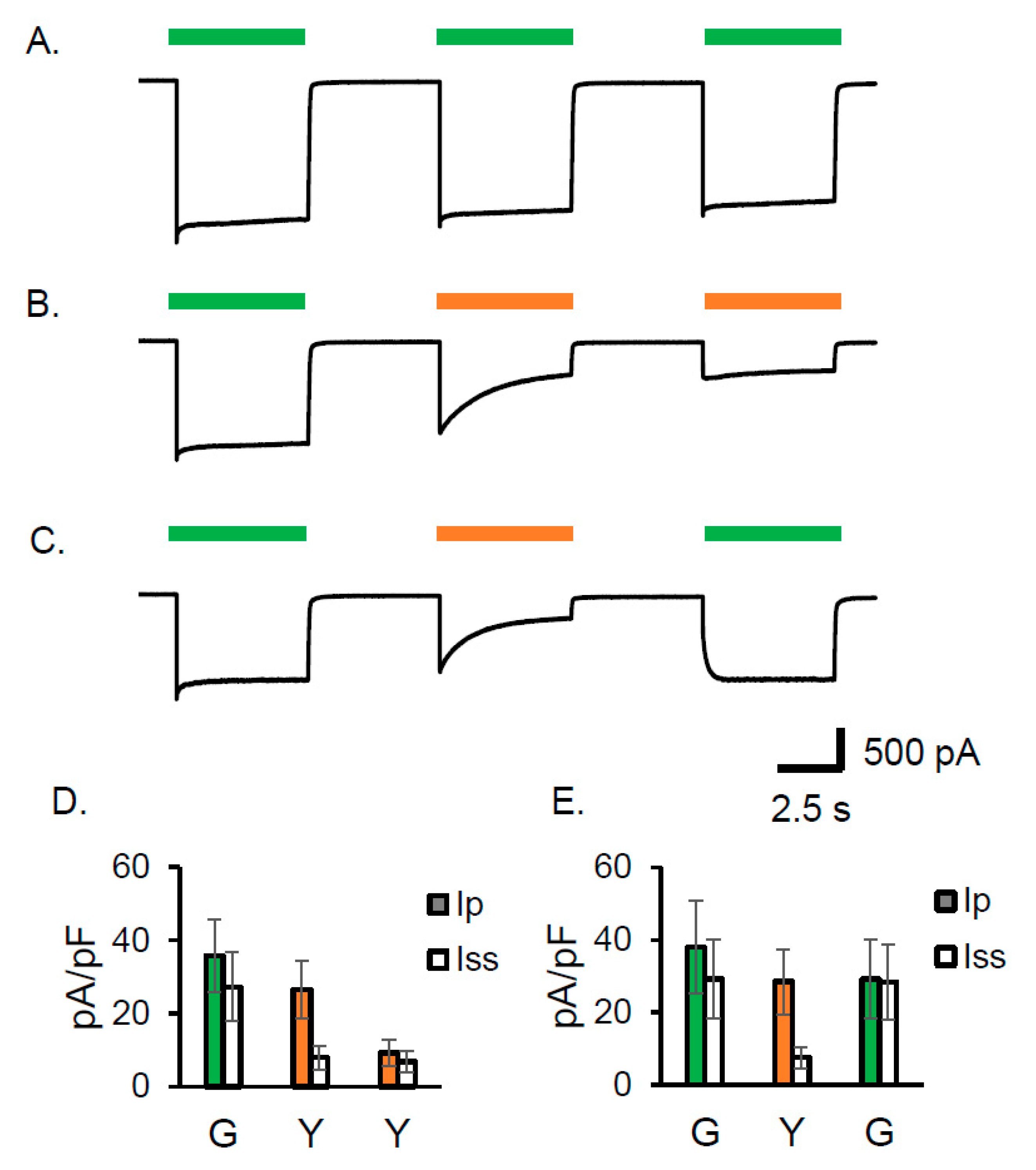
As already shown in Figure 1A, one of the obvious characteristics of Gt_CCR4 is the small inactivation upon 530 nm illumination which is close to λmax (525 nm). Here we compared the current shape by illumination of 530 nm and 590 nm light (Figure 5A–C). Repetitive illuminations of 530 nm light three times gave almost the same current shape, which has a high steady state level (Figure 5A). Upon illumination of 590 nm light, the current slowly decayed into a small steady state level, which was further reduced after a second illumination of 590 nm LED following several seconds of a dark period (Figure 5B,D). Even after 30 sec or 2 min, the inactivated current did not recover (Figure S1A,B). These observations suggest that Gt_CCR4 possesses a long-lived inactivated state which could accumulate in 590 nm light. Only illuminating with 530 nm allowed for a full recovery to the original steady state level (Figure 5C,E).
4. Discussion
In this study, we aimed to elucidate the ion-channel properties of a recently discovered light-gated cation channel Gt_CCR4 from a cryptophyte and compare it to well-known Cr_ChR2 from Chlamydomonas reinhardtii by using an electrophysiological method. In ND7/23 cells, Gt_CCR4 showed a large current density (Figure 1C and Figure 2A,B). Inactivation of the photocurrent obtained from Gt_CCR4 was smaller than in Cr_ChR2. In other words, a large current was observed under constant light (Figure 1A,B). This was also obvious in the current trace after illumination at a high frequency (Figure 4). These characteristics promise stable and reproducible stimulation of neuronal excitability by Gt_CCR4.
The light sensitivity of Gt_CCR4 is higher than that of Cr_ChR2 with a particular steady state component (Iss) (Figure 1E,F). ChR variants with high light sensitivity have already been developed [19,20], but those have a long channel life time with at least two orders of magnitude or even much longer. Therefore, these are inappropriate for high-frequency light stimulation. In contrast, Gt_CCR4 has a short open life time of 25–30 ms, which is about the same range as Cr_ChR2, i.e., 10–15 ms (Figure 1D). Together, Gt_CCR4 is light sensitive and useful as an optogenetics tool with high time resolution. Optical irradiation causes heat and elevates temperature by 0.2~2 °C, especially in cranial nerve experiments [37]. Moreover, it has been demonstrated that the rise in temperature suppressed neuronal spiking in multiple brain regions, serving as a warning of the use of strong light for neuronal stimulation. Such an undesirable artefact has to be avoided by lowering light intensity, while effective depolarization has to be stably maintained. Gt_CCR4 has the potential for overcoming this problem.
H+ permeability is high for Cr_ChR2 [8]. Permeability for Ca2+ has been reported, and not only for monovalent cations such as Na+ and K+ [8,21]. On the other hand, Gt_CCR4 showed high selectivity in monovalent metal cations and low H+ permeability. The permeability of a divalent cation such as Ca2+ seems to be very low or negligible. The position that is important to ion selectivity has been studied in Cr_ChR2. E90 in the central gate is crucial for cation/anion selection [23]. L132 in TM3 influences on Ca2+ permeability [21]. Duan and coworkers recently demonstrated that D156H and D156C mutation increase permeability for Na+ and K+ [38]. The outer gate in Chrimson (E139) on the extracellular side is important for Na+ extrusion [22]. These key residues are not conserved in Gt_CCR4, implying that a different ion selection property resides in DTD channels. It would be necessary to study selectivity based on variant analysis and structural information in the future. Considering its application in optogenetics, Gt_CCR4 would not cause a significant change to pH in the cell membrane because of its very low H+ permeability, which could be advantageous when an unknown effect by pH needs to be prevented. To enable optical stimulation without improper calcium signaling, Gt_CCR4 might work better than Cr_ChR2.
A single turnover photocurrent of Gt_CCR4 by laser irradiation provided a time constant (τoff) of 15–20 ms, which is smaller than that obtained by constant light (25–30 ms). This suggests two processes for channel opening and shutting. A dual photocycle model was indeed proposed for Cr_ChR2 [39]. It is expected that a similar reaction is caused in Gt_CCR4, but more experiments are needed to prove this.
We found a characteristic inactivation of channel activity by long wavelength absorption in Gt_CCR4 (Figure 5). Since inactivation lasted at least a few minutes, formation of a stable intermediate with long wavelength absorption is anticipated. Alternatively, Gt_CCR4 exhibits a photochromic property that is seen in the photocycle of Anabaena sensory rhodopsin [40]. Such photochromism or desensitization was also observed in chlorophyte channelrhodopsins [41,42]. We are now focusing on understanding the reaction mechanism in greater depth via a spectroscopic experiment. In conclusion, we here elucidated the cation channel properties of Gt_CCR4. Its high conductance and cation selectivity without significant inactivation would be an appropriate set of features for optogenetics applications. We are currently assessing the feasibility of Gt_CCR4 as an optical stimulator in cultured neurons.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-3417/9/17/3440/s1, Table S1 Amino acid alignments of bacteriorhodopsin (BR), Cr_ChR2 and Gt_CCR4. Figure S1: Ca2+ photecurrents of Gt_CCR4 and Cr_ChR2. Figure S2 Gt_CCR4 has a long-lived and long wavelength-absorbing inactivated state.
Author Contributions
Conceptualization, All authors; Data Curation, S.S. and S.H.; Validation, S.S., S.H. and S.P.T.; Writing—Original Draft Preparation, S.P.T.; Writing—Review & Editing, S.P.T.; Supervision, H.K. and S.P.T.
Funding
This work was funded by the Japanese Ministry of Education, Culture, Sports, Science and Technology (25104009, 15H02391 to H.K and 18K06109 to S.P.T.), a JST CREST grant (JPMJCR1753 to H.K.), and a JST PRESTO grant (JPMJPR1688 to S.P.T). S.H. is a Research Fellow of the Japan Society for the Promotion of Science (JSPS Research Fellow).
Acknowledgments
We thank Ryoko Nakamura for her excellent technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ernst, O.P.; Lodowski, D.T.; Elstner, M.; Hegemann, P.; Brown, L.S.; Kandori, H. Microbial and animal rhodopsins: Structures, functions, and molecular mechanisms. Chem. Rev. 2014, 114, 126–163. [Google Scholar] [CrossRef] [PubMed]
- Oesterhelt, D.; Stoeckenius, W. Rhodopsin-like protein from the purple membrane of Halobacterium halobium. Nat. New Biol. 1971, 233, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Matsuno-Yagi, A.; Mukohata, Y. Two Possible Roles of Bacteriorhodopsin; a Comparative Study of Strains of Halobacz’eril’m Halobium Differing in Pigmentation. Biochem. Biophys. Res. Commun. 1977, 78, 237–243. [Google Scholar] [CrossRef]
- Inoue, K.; Ono, H.; Abe-Yoshizumi, R.; Yoshizawa, S.; Ito, H.; Kogure, K.; Kandori, K. A light-driven sodium ion pump in marine bacteria. Nat. Commun. 2013, 4, 1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Ito, S.; Kato, Y.; Nomura, Y.; Shibata, M.; Uchihashi, T.; Tsunoda, S.P.; Kandori, H. A natural light-driven inward proton pump. Nat. Commun. 2016, 7, 13415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polovinkin, V.; Rodriguez-Valera, F.; Bueldt, G.; Kovalev, K.; Alekseev, A.; Gordeliy, V.; Chizhov, I.; Juettner, J.; Bamann, C.; Manstein, D.J.; et al. Inward H + pump xenorhodopsin: Mechanism and alternative optogenetic approach. Sci. Adv. 2017, 3, e1603187. [Google Scholar]
- Nagel, G.; Ollig, D.; Fuhrmann, M.; Kateriya, S.; Musti, A.M.; Bamberg, E.; Hegemann, P. Channelrhodopsin-1: A Light-Gated Proton Channel in Green Algae. Science 2002, 296, 2395–2398. [Google Scholar] [CrossRef]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.E.; Zhang, F.; Yizhar, O.; Ramakrishnan, C.; Nishizawa, T.; Hirata, K.; Ito, J.; Aita, Y.; Tsukazaki, T.; Hayashi, S.; et al. Crystal structure of the channelrhodopsin light-gated cation channel. Nature 2012, 482, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Volkov, O.; Kovalev, K.; Polovinkin, V.; Borshchevskiy, V.; Bamann, C.; Astashkin, R.; Marin, E.; Popov, A.; Balandin, T.; Willbold, D.; et al. Structural insights into ion conduction by channelrhodopsin 2. Science 2017, 358, pii: eaan8862. [Google Scholar] [CrossRef]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef]
- Ishizuka, T.; Kakuda, M.; Araki, R.; Yawo, H. Kinetic evaluation of photosensitivity in genetically engineered neurons expressing green algae light-gated channels. Neurosci. Res. 2006, 54, 85–94. [Google Scholar] [CrossRef]
- Schneider, F.; Grimm, C.; Hegemann, P. Biophysics of Channelrhodopsin. Annu. Rev. Biophys. 2015, 44, 167–186. [Google Scholar] [CrossRef] [Green Version]
- Prigge, M.; Schneider, F.; Tsunoda, S.P.; Shilyansky, C.; Wietek, J.; Deisseroth, K.; Hegemann, P. Color-tuned channelrhodopsins for multiwavelength optogenetics. J. Biol. Chem. 2012, 287, 31804–31812. [Google Scholar] [CrossRef]
- Wang, H.; Sugiyama, Y.; Hikima, T.; Sugano, E.; Tomita, H.; Takahashi, T.; Ishizuka, T.; Yawo, H. Molecular determinants differentiating photocurrent properties of two channelrhodopsins from Chlamydomonas. J. Biol. Chem. 2009, 284, 5685–5696. [Google Scholar] [CrossRef]
- Tsunoda, S.P.; Hegemann, P. Glu 87 of channelrhodopsin-1 causes pH-dependent color tuning and fast photocurrent inactivation. Photochem. Photobiol. 2009, 85, 564–569. [Google Scholar] [CrossRef]
- Klapoetke, N.C.; Murata, Y.; Kim, S.S.; Pulver, S.R.; Birdsey-Benson, A.; Cho, Y.K.; Morimoto, T.K.; Chuong, A.S.; Carpenter, E.J.; Tian, Z.; et al. Independent optical excitation of distinct neural populations. Nat. Methods 2014, 11, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Govorunova, E.G.; Sineshchekov, O.A.; Li, H.; Janz, R.; Spudich, J.L. Characterization of a highly efficient blue-shifted channelrhodopsin from the marine alga platymonas subcordiformis. J. Biol. Chem. 2013, 288, 29911–29922. [Google Scholar] [CrossRef]
- Berndt, A.; Yizhar, O.; Gunaydin, L.A.; Hegemann, P.; Deisseroth, K. Bi-stable neural state switches. Nat. Neurosci. 2009, 12, 229–234. [Google Scholar] [CrossRef]
- Yizhar, O.; Fenno, L.E.; Prigge, M.; Schneider, F.; Davidson, T.J.; Ogshea, D.J.; Sohal, V.S.; Goshen, I.; Finkelstein, J.; Paz, J.T.; et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 2011, 477, 171–178. [Google Scholar] [CrossRef]
- Kleinlogel, S.; Feldbauer, K.; Dempski, R.E.; Fotis, H.; Wood, P.G.; Bamann, C.; Bamberg, E. Ultra light-sensitive and fast neuronal activation with the Ca2+-permeable channelrhodopsin CatCh. Nat. Neurosci. 2011, 14, 513–518. [Google Scholar] [CrossRef]
- Vierock, J.; Grimm, C.; Nitzan, N.; Hegemann, P. Molecular determinants of proton selectivity and gating in the red-light activated channelrhodopsin Chrimson. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Wietek, J.; Wiegert, J.S.; Adeishvili, N.; Schneider, F.; Watanabe, H.; Tsunoda, S.P.; Vogt, A.; Elstner, M.; Oertner, T.G.; Hegemann, P. Conversion of channelrhodopsin into a light-gated chloride channel. Science 2014, 344, 409–412. [Google Scholar] [CrossRef]
- Berndt, A.; Lee, S.Y.; Ramakrishnan, C.; Deisseroth, K. Structure-guided transformation of channelrhodopsin into a light-activated chloride channel. Science 2014, 344, 420–424. [Google Scholar] [CrossRef]
- Govorunova, E.G.; Sineshchekov, O.A.; Janz, R.; Liu, X.; Spudich, J.L. NEUROSCIENCE. Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics. Science 2015, 349, 647–650. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kato, H.E.; Yamashita, K.; Ito, S.; Inoue, K.; Ramakrishnan, C.; Fenno, L.E.; Evans, K.E.; Paggi, J.M.; Dror, R.O.; et al. Crystal structure of the natural anion-conducting channelrhodopsin GtACR1. Nature 2018, 561, 343–348. [Google Scholar] [CrossRef]
- Kato, H.E.; Kim, Y.S.; Paggi, J.M.; Evans, K.E.; Allen, W.E.; Richardson, C.; Inoue, K.; Ito, S.; Ramakrishnan, C.; Fenno, L.E.; et al. Structural mechanisms of selectivity and gating in anion channelrhodopsins. Nature 2018, 561, 349–371. [Google Scholar] [CrossRef]
- Spudich, J.L.; Li, H.; Huang, C.-Y.; Wang, M.; Schafer, C.T.; Sineshchekov, O.A.; Zheng, L.; Govorunova, E.G. Crystal structure of a natural light-gated anion channelrhodopsin. Elife 2019, 8, 1–21. [Google Scholar]
- Govorunova, E.G.; Sineshchekov, O.A.; Spudich, J.L. Structurally Distinct Cation Channelrhodopsins from Cryptophyte Algae. Biophys. J. 2016, 110, 2302–2304. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, Y.; Konno, M.; Ito, S.; Tsunoda, S.P.; Inoue, K.; Kandori, H. Molecular properties of a DTD channelrhodopsin from Guillardia theta. Biophys. Physicobiol. 2017, 14, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, Y.; Wang, H.; Hikima, T.; Sato, M.; Kuroda, J.; Takahashi, T.; Ishizuka, T.; Yawo, H. Photocurrent attenuation by a single polar-to-nonpolar point mutation of channelrhodopsin-2. Photochem. Photobiol. Sci. 2009, 8, 328–336. [Google Scholar] [CrossRef]
- Dawydow, A.; Gueta, R.; Ljaschenko, D.; Ullrich, S.; Hermann, M.; Ehmann, N.; Gao, S.; Fiala, A.; Langenhan, T.; Nagel, G.; et al. Channelrhodopsin-2-XXL, a powerful optogenetic tool for low-light applications. Proc. Natl. Acad. Sci. USA 2014, 111, 13972–13977. [Google Scholar] [CrossRef]
- Nack, M.; Radu, I.; Gossing, M.; Bamann, C.; Bamberg, E.; von Mollard, G.F.; Heberle, J.; Hofkens, J.; Pozzo, J.L.; Tosic, O.; et al. The DC gate in Channelrhodopsin-2: Crucial hydrogen bonding interaction between C128 and D156. Photochem. Photobiol. Sci.USA 2010, 9, 194–198. [Google Scholar] [CrossRef]
- Sineshchekov, O.A.; Govorunova, E.G.; Li, H.; Spudich, J.L. Bacteriorhodopsin-like channelrhodopsins: Alternative mechanism for control of cation conductance. Proc. Natl. Acad. Sci. 2017, 114, E9512–E9519. [Google Scholar] [CrossRef] [Green Version]
- Lorenz-Fonfria, V.A.; Resler, T.; Krause, N.; Nack, M.; Gossing, M.; Fischer von Mollard, G.; Bamann, C.; Bamberg, E.; Schlesinger, R.; Heberle, J. Transient protonation changes in channelrhodopsin-2 and their relevance to channel gating. Proc. Natl. Acad. Sci. 2013, 110, E1273–E1281. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.N.; Bevan, S.J.; Coote, P.R.; Dunn, P.M.; Harmar, A.; Hogan, P.; Latchman, D.S.; Morrison, C.; Rougon, G.; Theveniau, M.; et al. Novel Cell Lines Display Properties of Nociceptive Sensory Neurons. Proc. R. Soc. B Biol. Sci. 1990, 241, 187–194. [Google Scholar]
- Owen, S.F.; Liu, M.H.; Kreitzer, A.C. Thermal constraints on in vivo optogenetic manipulations. Nat. Neurosci. 2019, 22, 1061–1065. [Google Scholar] [CrossRef]
- Duan, X.; Nagel, G.; Gao, S. Mutated Channelrhodopsins with Increased Sodium and Calcium Permeability. Appl. Sci. 2019, 9, 664. [Google Scholar] [CrossRef]
- Kuhne, J.; Vierock, J.; Tennigkeit, S.A.; Dreier, M.-A.; Wietek, J.; Petersen, D.; Gavriljuk, K.; El-Mashtoly, S.F.; Hegemann, P.; Gerwert, K. Unifying photocycle model for light adaptation and temporal evolution of cation conductance in channelrhodopsin-2. Proc. Natl. Acad. Sci USA 2019, 116, 9380–9389. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, A.; Furutani, Y.; Jung, K.H.; Kandori, H. Photochromism of Anabaena sensory rhodopsin. J. Am. Chem. Soc. 2007, 129, 8644–8649. [Google Scholar] [CrossRef]
- Zamani, A.; Sakuragi, S.; Ishizuka, T.; Yawo, H. Kinetic characteristics of chimeric channelrhodopsins implicate the molecular identity involved in desensitization. Biophys. Physicobiol. 2017, 14, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.Y. A User’s Guide to Channelrhodopsin Variants: Features, Limitations and Future Developments. Exp. Physiol. 2011, 96, 19–25. [Google Scholar] [CrossRef]
Figure 1.
Basic properties of Gt_CCR4 and Cr_ChR2. (A,B) Each cation channelrhodopsin expressed in ND7/23 cells was stimulated by green (530 nm) or blue (470 nm) LED light (6.8 mW/mm2). Standard solutions were used. Membrane potentials were clamped from −60 mV to +60 mV in +20 mV steps. Fluorescence images were taken using a confocal microscope. (C) Current-voltage relationship (I-V plot) of Gt_CCR4 (filled symbol) and Cr_ChR2 (empty symbol). Current peak component (square) and steady state amplitude (circle) of two channels are depicted. (D) Current-decay kinetics of Gt_CCR4 and Cr_ChR2. τoff is plotted as a function of membrane voltage. Filled circle; Gt_CCR4, empty circle; Cr_ChR2. (E,F) Light power dependency of photocurrents from Gt_CCR4 and Cr_ChR2 at −60 mV. Each channel was stimulated by 530 nm (Gt_CCR4) and 470 nm (Cr_ChR2). Photocurrent values are normalized. Current peak component (filled circle) and steady state amplitude (empty circle) are depicted. (n = 4–8 cells).
Figure 1.
Basic properties of Gt_CCR4 and Cr_ChR2. (A,B) Each cation channelrhodopsin expressed in ND7/23 cells was stimulated by green (530 nm) or blue (470 nm) LED light (6.8 mW/mm2). Standard solutions were used. Membrane potentials were clamped from −60 mV to +60 mV in +20 mV steps. Fluorescence images were taken using a confocal microscope. (C) Current-voltage relationship (I-V plot) of Gt_CCR4 (filled symbol) and Cr_ChR2 (empty symbol). Current peak component (square) and steady state amplitude (circle) of two channels are depicted. (D) Current-decay kinetics of Gt_CCR4 and Cr_ChR2. τoff is plotted as a function of membrane voltage. Filled circle; Gt_CCR4, empty circle; Cr_ChR2. (E,F) Light power dependency of photocurrents from Gt_CCR4 and Cr_ChR2 at −60 mV. Each channel was stimulated by 530 nm (Gt_CCR4) and 470 nm (Cr_ChR2). Photocurrent values are normalized. Current peak component (filled circle) and steady state amplitude (empty circle) are depicted. (n = 4–8 cells).

Figure 2.
Ion selectivity of Gt_CCR4 and Cr_ChR2. Each channelrhodopsin expressed in ND7/23 cells was stimulated by green and blue LED light. I-V plot of Gt_CCR4 (A) and Cr_ChR2 (B) are depicted. Steady-state current density (pA/pF) in 20 mV steps from −100 mV to +80 mV was plotted. The present liquid junction potential was considered. The pipette solution contained 110 mM NMG-Cl at pH 9.0, and the bath solution varied: black, NMG-Cl at pH 9.0; red, NMG-Cl at pH 6.85; green, NaCl at pH 9.0; blue, KCl at pH 9.0; grey, CsCl at pH 9.0. See “Materials and Methods” for details about the solutions. (C,D) Comparison of current density of Gt_CCR4 (C) and Cr_ChR2 (D) in the presence of various cations at −60 mV. (E) Reversal potential shift (ΔErev) for each condition for Gt_CCR4 and Cr_ChR2. Erev was determined from the I-V plot shown in A,B. Each Erev value was subtracted from the Erev at NMG-Cl at pH 9.0. (F) Permeability ratio for H+ and Na+ in Gt_CCR4 and Cr_ChR2 as estimated from current value and ionic concentration. (n = 6–9 cells).
Figure 2.
Ion selectivity of Gt_CCR4 and Cr_ChR2. Each channelrhodopsin expressed in ND7/23 cells was stimulated by green and blue LED light. I-V plot of Gt_CCR4 (A) and Cr_ChR2 (B) are depicted. Steady-state current density (pA/pF) in 20 mV steps from −100 mV to +80 mV was plotted. The present liquid junction potential was considered. The pipette solution contained 110 mM NMG-Cl at pH 9.0, and the bath solution varied: black, NMG-Cl at pH 9.0; red, NMG-Cl at pH 6.85; green, NaCl at pH 9.0; blue, KCl at pH 9.0; grey, CsCl at pH 9.0. See “Materials and Methods” for details about the solutions. (C,D) Comparison of current density of Gt_CCR4 (C) and Cr_ChR2 (D) in the presence of various cations at −60 mV. (E) Reversal potential shift (ΔErev) for each condition for Gt_CCR4 and Cr_ChR2. Erev was determined from the I-V plot shown in A,B. Each Erev value was subtracted from the Erev at NMG-Cl at pH 9.0. (F) Permeability ratio for H+ and Na+ in Gt_CCR4 and Cr_ChR2 as estimated from current value and ionic concentration. (n = 6–9 cells).
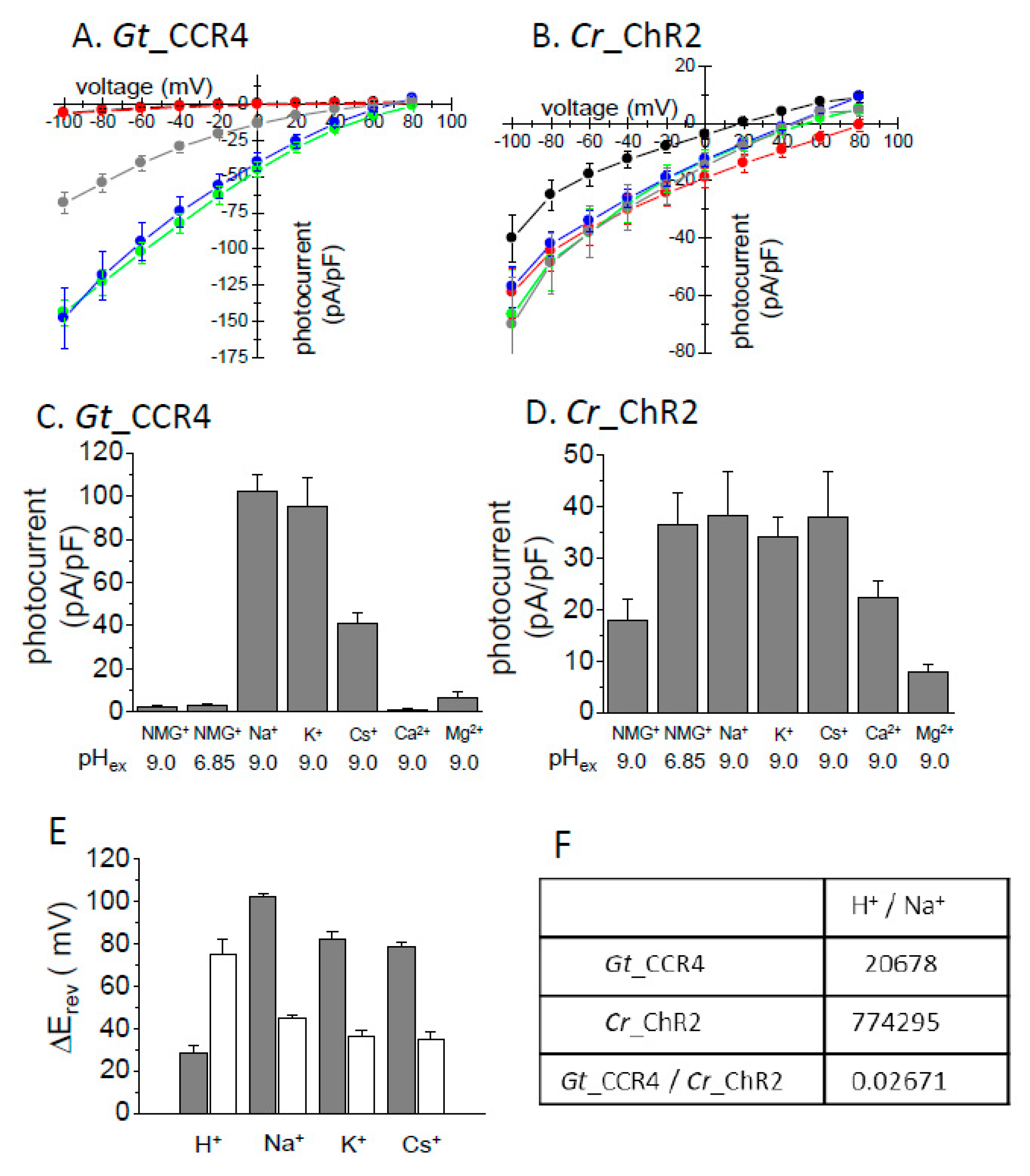
Figure 3.
Flash laser stimulation. Standard solutions were used. Each channelrhodopsin in ND7/23 cells was stimulated by 5 ns by a green flash laser. Representative trace generated by Gt_CCR4 (A) and Cr_ChR2 (B) in 20 mV steps from −100 mV to +80 mV. (C) τon-voltage relationship from Gt_CCR4 (filled circle) and Cr_ChR2 (empty circle). Current rise was fitted by a single exponential function. The time constant was plotted. (D) τoff-voltage relationship from Gt_CCR4 (filled circle) and Cr_ChR2 (empty circle). Current decay was fitted by a single exponential function. (n = 5–7 cells).
Figure 3.
Flash laser stimulation. Standard solutions were used. Each channelrhodopsin in ND7/23 cells was stimulated by 5 ns by a green flash laser. Representative trace generated by Gt_CCR4 (A) and Cr_ChR2 (B) in 20 mV steps from −100 mV to +80 mV. (C) τon-voltage relationship from Gt_CCR4 (filled circle) and Cr_ChR2 (empty circle). Current rise was fitted by a single exponential function. The time constant was plotted. (D) τoff-voltage relationship from Gt_CCR4 (filled circle) and Cr_ChR2 (empty circle). Current decay was fitted by a single exponential function. (n = 5–7 cells).
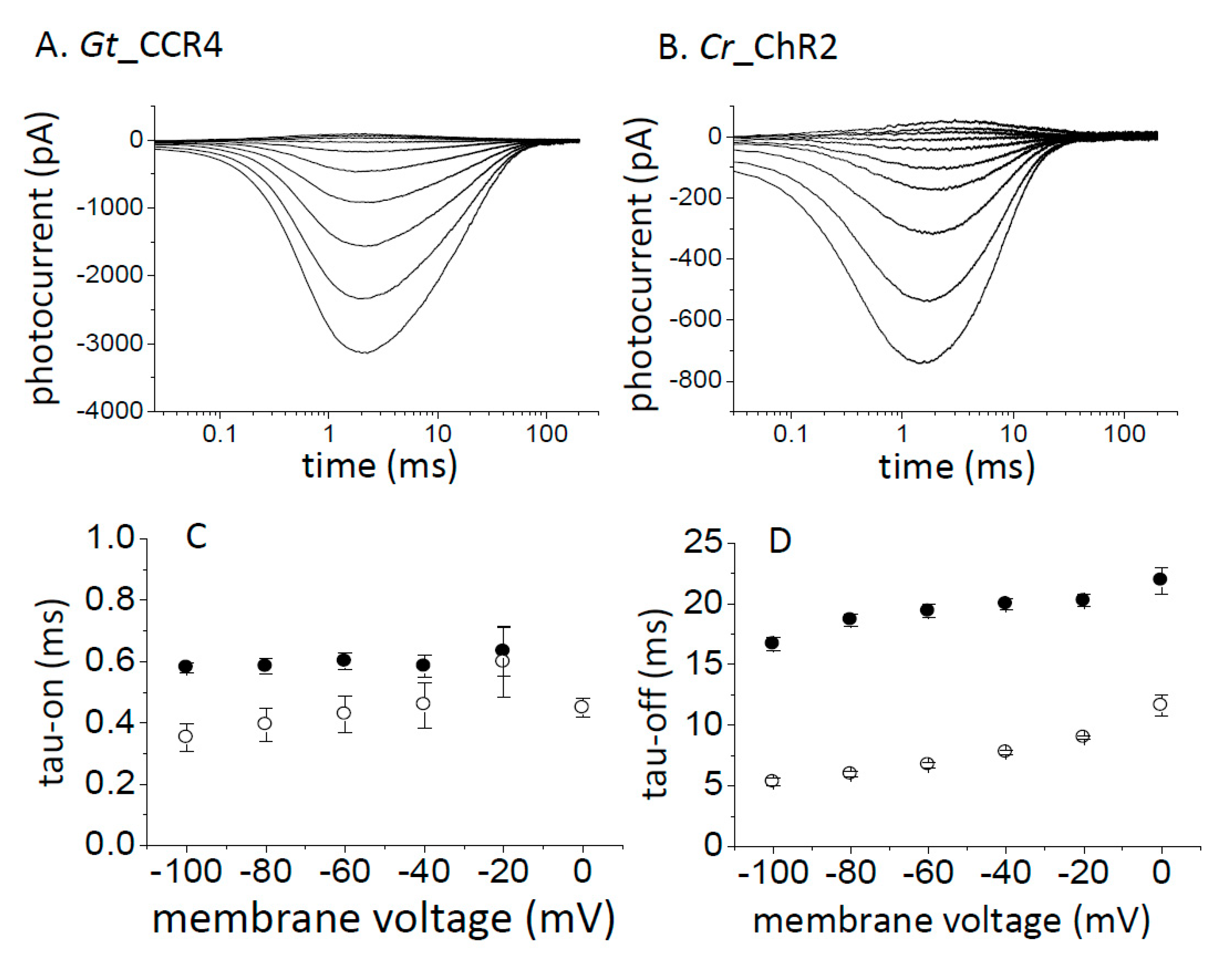
Figure 4.
High frequency stimulation of Gt_CCR4 and Cr_ChR2. (A,C,E) Gt_CCR4 in ND7/23 cells was stimulated by green LED light with a frequency of 10, 20 or 50 Hz as indicated by colored dots or a line under each trace. Membrane voltage was clamped at −60 mV. (B,D,F) Similarly, Cr_ChR2 was stimulated by blue LED light with a frequency of 10, 20 or 50 Hz as indicated by colored dots or a line under each trace. (G) Residual tail current at three different light frequencies is summarized. Residual tail current is the current amplitude right before the next photo stimulation indicated by an arrowhead in A,B. The current value was normalized to the peak amplitude as 100%. A standard solution was used. (n = 3 cells).
Figure 4.
High frequency stimulation of Gt_CCR4 and Cr_ChR2. (A,C,E) Gt_CCR4 in ND7/23 cells was stimulated by green LED light with a frequency of 10, 20 or 50 Hz as indicated by colored dots or a line under each trace. Membrane voltage was clamped at −60 mV. (B,D,F) Similarly, Cr_ChR2 was stimulated by blue LED light with a frequency of 10, 20 or 50 Hz as indicated by colored dots or a line under each trace. (G) Residual tail current at three different light frequencies is summarized. Residual tail current is the current amplitude right before the next photo stimulation indicated by an arrowhead in A,B. The current value was normalized to the peak amplitude as 100%. A standard solution was used. (n = 3 cells).

Figure 5.
Gt_CCR4 is inactivated by light with a longer wavelength. Standard solutions were used. Membrane voltage was clamped at −60 mV. Photocurrent by 530 nm light (7.44 mW/mm2) in (A) reached a steady state level after transient peak current. Repetitive stimulations gave almost identical current shapes. (B) Slow inactivation of the photocurrent was observed by illumination with 590 nm light (7.44 mW/mm2). The current was further reduced when illuminated twice with 590 nm light. (C) After 590 nm inactivation, 530 nm light fully reactivated the photocurrent to the original steady state level. (D) Photocurrent density from the measurement shown in (B). The photocurrent density upon exposure to the first 530 nm light (shown in G), and the second and third 590 nm light (Y), is shown. Ip; transient peak component. Iss; steady state component. (E) Photocurrent density from the measurement shown in C. The photocurrent density upon exposure to the first 530 nm light (shown in G), the second 590 nm light (Y), and the third 530 nm light (shown in G), is shown. (n = 4 cells).
Figure 5.
Gt_CCR4 is inactivated by light with a longer wavelength. Standard solutions were used. Membrane voltage was clamped at −60 mV. Photocurrent by 530 nm light (7.44 mW/mm2) in (A) reached a steady state level after transient peak current. Repetitive stimulations gave almost identical current shapes. (B) Slow inactivation of the photocurrent was observed by illumination with 590 nm light (7.44 mW/mm2). The current was further reduced when illuminated twice with 590 nm light. (C) After 590 nm inactivation, 530 nm light fully reactivated the photocurrent to the original steady state level. (D) Photocurrent density from the measurement shown in (B). The photocurrent density upon exposure to the first 530 nm light (shown in G), and the second and third 590 nm light (Y), is shown. Ip; transient peak component. Iss; steady state component. (E) Photocurrent density from the measurement shown in C. The photocurrent density upon exposure to the first 530 nm light (shown in G), the second 590 nm light (Y), and the third 530 nm light (shown in G), is shown. (n = 4 cells).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shigemura, S.; Hososhima, S.; Kandori, H.; Tsunoda, S.P. Ion Channel Properties of a Cation Channelrhodopsin, Gt_CCR4. Appl. Sci. 2019, 9, 3440. https://doi.org/10.3390/app9173440
AMA Style
Shigemura S, Hososhima S, Kandori H, Tsunoda SP. Ion Channel Properties of a Cation Channelrhodopsin, Gt_CCR4. Applied Sciences. 2019; 9(17):3440. https://doi.org/10.3390/app9173440
Chicago/Turabian StyleShigemura, Shunta, Shoko Hososhima, Hideki Kandori, and Satoshi P. Tsunoda. 2019. "Ion Channel Properties of a Cation Channelrhodopsin, Gt_CCR4" Applied Sciences 9, no. 17: 3440. https://doi.org/10.3390/app9173440
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.