Functional Near-Infrared Spectroscopy Recordings of Visuospatial Working Memory Processes. Part II: A Replication Study in Children on Sensitivity and Mental-Ability-Induced Differences in Functional Activation

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Assessment of Psychometric Intelligence
2.3. Quantification of Individual Visuospatial WM Spans
2.4. Experimental and Active Control Conditions of the fNIRS Study
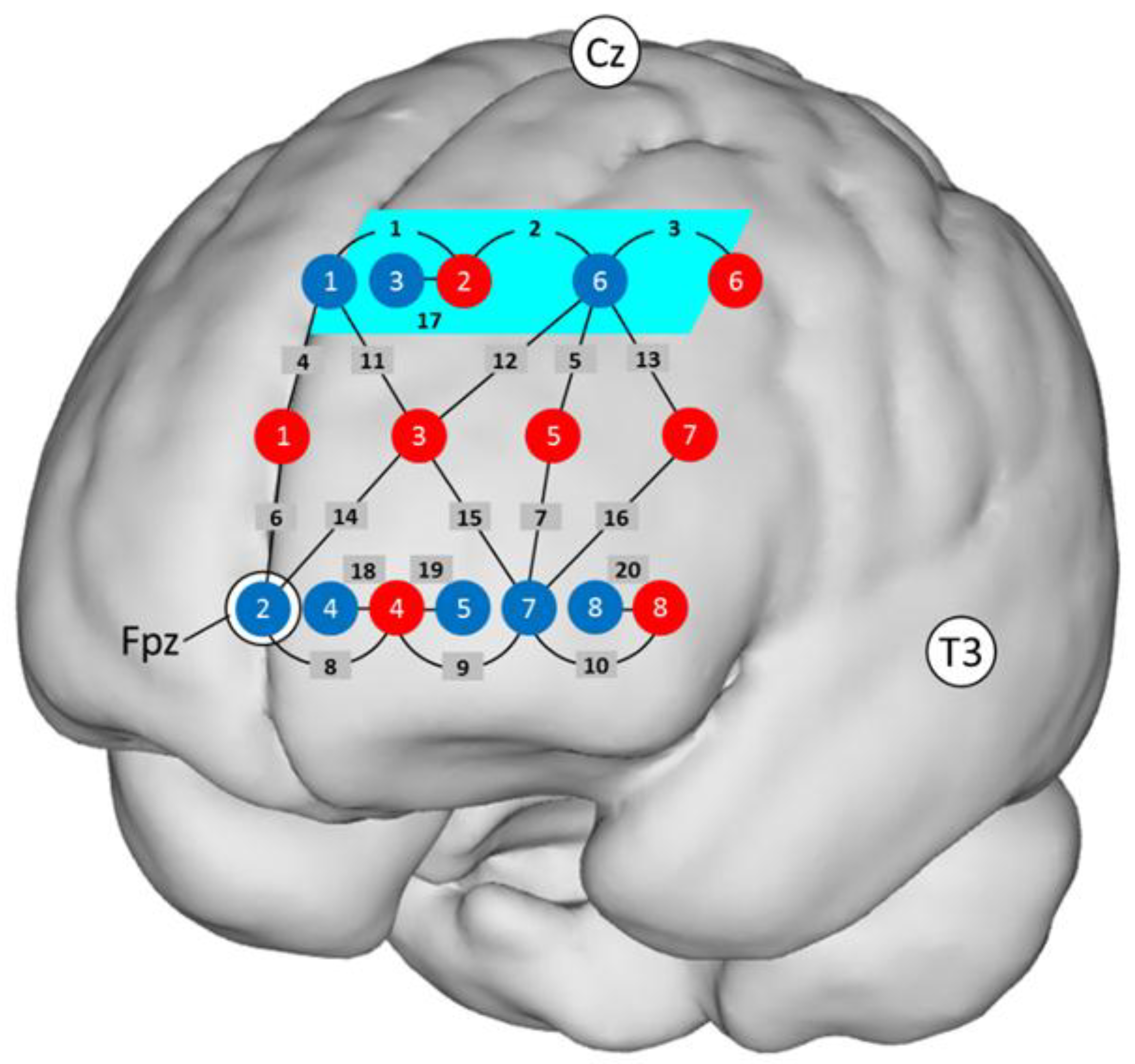
2.5. fNIRS Recordings
2.6. Time Course of the Study
3. Results
3.1. Behavioral Data
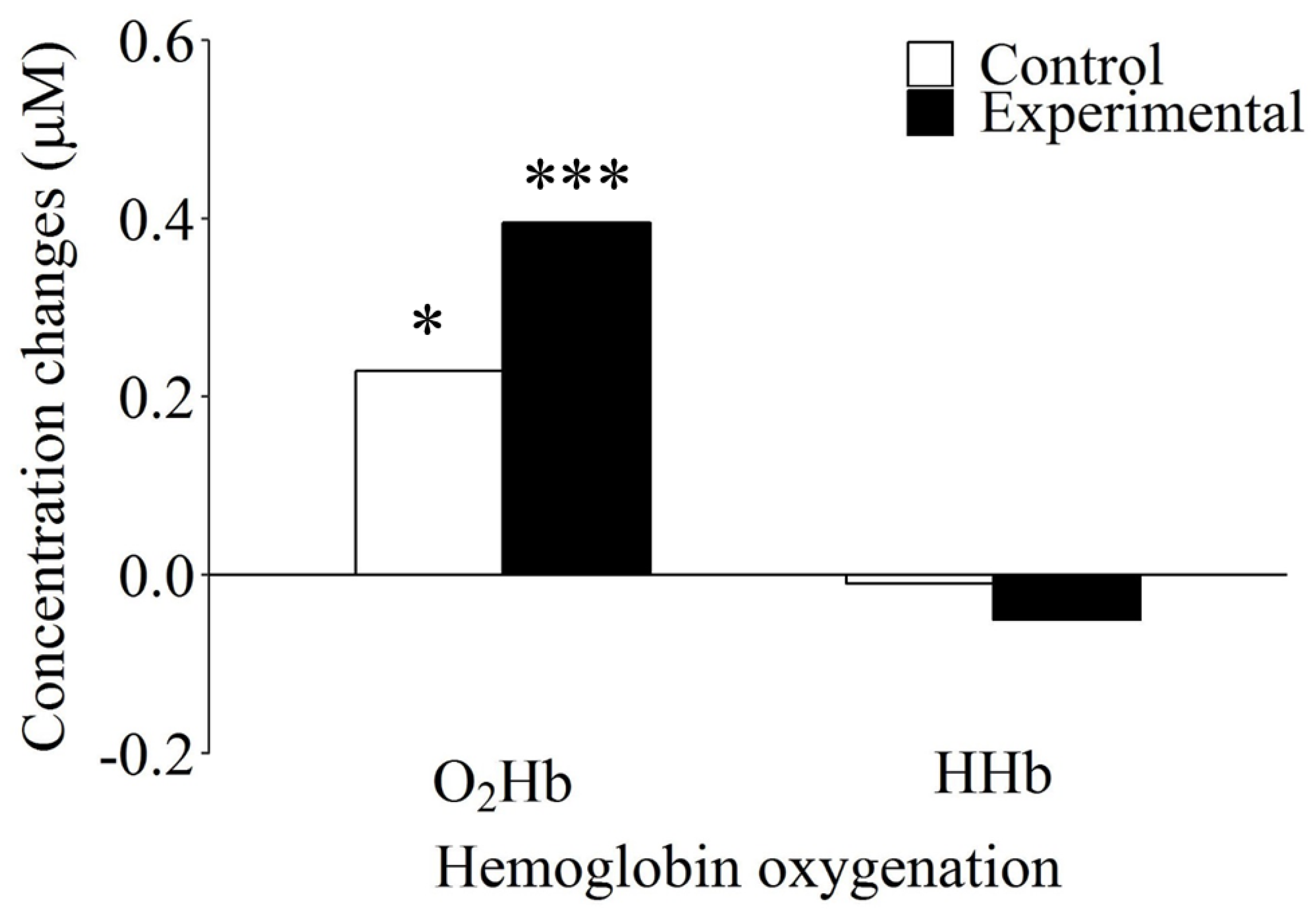
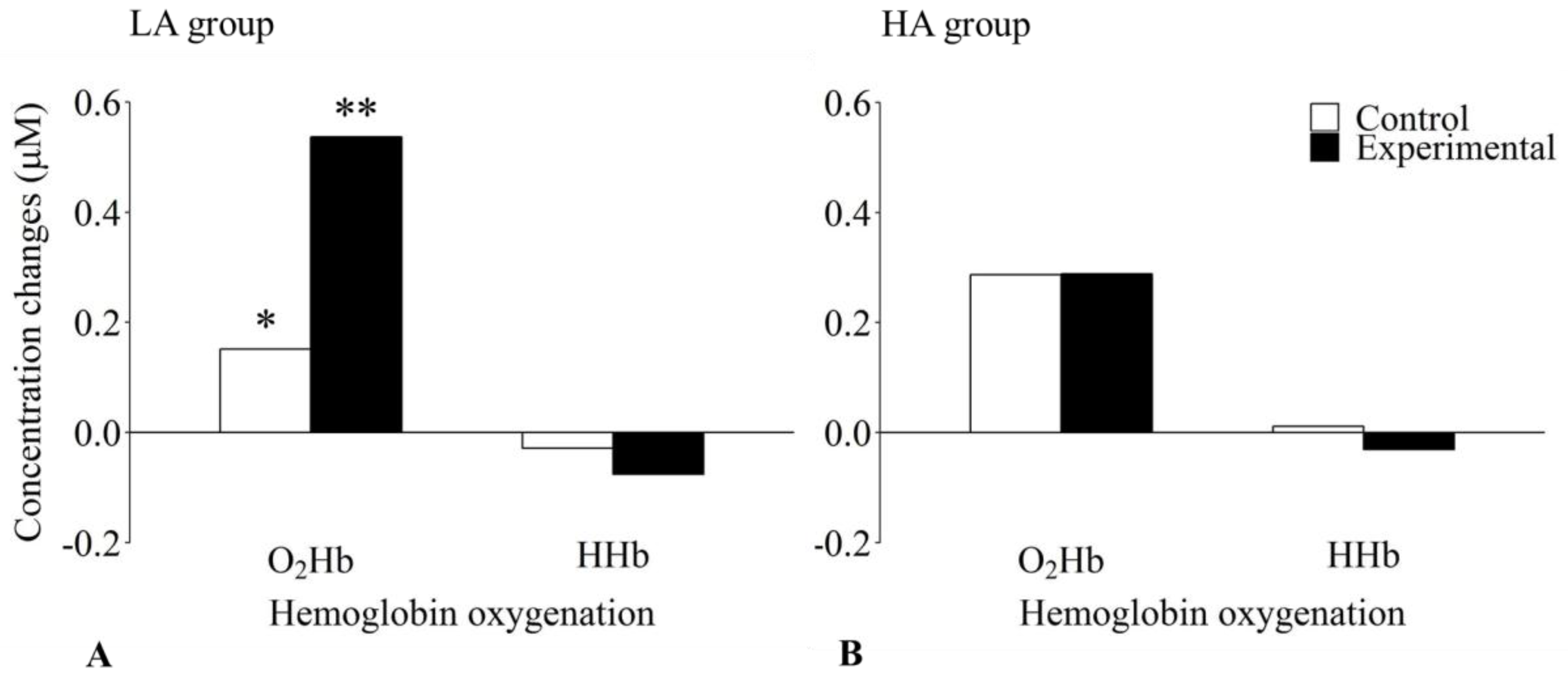
3.2. fNIRS Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chance, B.; Zhuang, Z.; Unah, C.; Alter, C.; Lipton, L. Cognition-activated low-frequency modulation of light absorption in human brain. Proc. Natl. Acad. Sci. USA 1993, 90, 3770–3774. [Google Scholar] [CrossRef] [PubMed]
- Villringer, A.; Planck, J.; Hock, C.; Schleinkofer, L.; Dirnagl, U. Near infrared spectroscopy (NIRS): A new tool to study hemodynamic changes during activation of brain function in human adults. Neurosci. Lett. 1993, 154, 101–104. [Google Scholar] [CrossRef]
- Tak, S.; Ye, J.C. Statistical analysis of fNIRS data: A comprehensive review. Neuroimage 2014, 85, 72–91. [Google Scholar] [CrossRef] [PubMed]
- Kirilina, E.; Jelzow, A.; Heine, A.; Niessing, M.; Wabnitz, H.; Brühl, R.; Ittermann, B.; Jacobs, A.M.; Tachtsidis, I. The physiological origin of task-evoked systemic artefacts in functional near infrared spectroscopy. Neuroimage 2012, 61, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villringer, A.; Chance, B. Non-invasive optical spectroscopy and imaging of human brain function. Trends Neurosci. 1997, 20, 435–442. [Google Scholar] [CrossRef]
- Tachtsidis, I.; Scholkmann, F. False positives and false negatives in functional near-infrared spectroscopy: Issues, challenges, and the way forward. Neurophotonics 2016, 3, 39801. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Bray, S.; Reiss, A.L. Functional near infrared spectroscopy (NIRS) signal improvement based on negative correlation between oxygenated and deoxygenated hemoglobin dynamics. Neuroimage 2010, 49, 3039–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malonek, D.; Grinvald, A. Interactions between electrical activity and cortical microcirculation revealed by imaging spectroscopy: Implications for functional brain mapping. Science 1996, 272, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Sheth, S.A.; Nemoto, M.; Guiou, M.; Walker, M.; Pouratian, N.; Toga, A.W. Linear and nonlinear relationships between neuronal activity, oxygen metabolism, and hemodynamic responses. Neuron 2004, 42, 347–355. [Google Scholar] [CrossRef]
- Boas, D.A.; Strangman, G.; Culver, J.P.; Hoge, R.D.; Jasdzewski, G.; Poldrack, R.A.; Rosen, B.R.; Mandeville, J.B. Can the cerebral metabolic rate of oxygen be estimated with near-infrared spectroscopy? Phys. Med. Biol. 2003, 48, 2405–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Fuchino, Y.; Kiguchi, M.; Katura, T.; Maki, A.; Yoro, T.; Koizumi, H. Intersubject variability of near-infrared spectroscopy signals during sensorimotor cortex activation. J. Biomed. Opt. 2005, 10, 44001. [Google Scholar] [CrossRef] [PubMed]
- Boas, D.A.; Elwell, C.E.; Ferrari, M.; Taga, G. Twenty years of functional near-infrared spectroscopy: Introduction for the special issue. Neuroimage 2014, 85. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, H.; Shewokis, P.A.; Bunce, S.; Izzetoglu, K.; Willems, B.; Onaral, B. Optical brain monitoring for operator training and mental workload assessment. Neuroimage 2012, 59, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Molteni, E.; Contini, D.; Caffini, M.; Baselli, G.; Spinelli, L.; Cubeddu, R.; Cerutti, S.; Bianchi, A.M.; Torricelli, A. Load-dependent brain activation assessed by time-domain functional near-infrared spectroscopy during a working memory task with graded levels of difficulty. J. Biomed. Opt. 2012, 17, 56005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishburn, F.A.; Norr, M.E.; Medvedev, A.V.; Vaidya, C.J. Sensitivity of fNIRS to cognitive state and load. Front. Hum. Neurosci. 2014, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Herff, C.; Heger, D.; Fortmann, O.; Hennrich, J.; Putze, F.; Schultz, T. Mental workload during n-back task—Quantified in the prefrontal cortex using fNIRS. Front. Hum. Neurosci. 2014, 7, 935. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Luo, Q.; Gong, H. Gender-specific hemodynamics in prefrontal cortex during a verbal working memory task by near-infrared spectroscopy. Behav. Brain Res. 2010, 209, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Chua, Z.; Peysakhovich, V.; Del Campo, N.; Matton, N. Mental workload and neural efficiency quantified in the prefrontal cortex using fNIRS. Sci. Rep. 2017, 7, 5222. [Google Scholar] [CrossRef] [PubMed]
- Huppert, T.J.; Hoge, R.D.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. A temporal comparison of BOLD, ASL, and NIRS hemodynamic responses to motor stimuli in adult humans. Neuroimage 2006, 29, 368–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinschmidt, A.; Obrig, H.; Requardt, M.; Merboldt, K.D.; Dirnagl, U.; Villringer, A.; Frahm, J. Simultaneous recording of cerebral blood oxygenation changes during human brain activation by magnetic resonance imaging and near-infrared spectroscopy. J. Cereb. Blood Flow Metab. 1996, 16, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Mehagnoul-Schipper, D.J.; Van Der Kallen, B.F.W.; Colier, W.N.J.M.; Van Der Sluijs, M.C.; Van Erning, L.J.T.O.; Thijssen, H.O.M.; Oeseburg, B.; Hoefnagels, W.H.L.; Jansen, R.W.M.M. Simultaneous measurements of cerebral oxygenation changes during brain activation by near-infrared spectroscopy and functional magnetic resonance imaging in healthy young and elderly subjects. Hum. Brain Map. 2002, 16, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Yahata, N.; Funane, T.; Takizawa, R.; Katura, T.; Atsumori, H.; Nishimura, Y.; Kinoshita, A.; Kiguchi, M.; Koizumi, H.; et al. A NIRS-fMRI investigation of prefrontal cortex activity during a working memory task. Neuroimage 2013, 83, 158–173. [Google Scholar] [CrossRef] [PubMed]
- Strangman, G.; Culver, J.P.; Thompson, J.H.; Boas, D.A. A quantitative comparison of simultaneous BOLD fMRI and NIRS recordings during functional brain activation. Neuroimage 2002, 17, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Toronov, V.; Webb, A.; Choi, J.H.; Wolf, M.; Michalos, A.; Gratton, E.; Hueber, D. Investigation of human brain hemodynamics by simultaneous near-infrared spectroscopy and functional magnetic resonance imaging. Med. Phys. 2001, 28, 521. [Google Scholar] [CrossRef] [PubMed]
- Funane, T.; Sato, H.; Yahata, N.; Takizawa, R.; Nishimura, Y.; Kinoshita, A.; Katura, T.; Atsumori, H.; Fukuda, M.; Kasai, K.; et al. Concurrent fNIRS-fMRI measurement to validate a method for separating deep and shallow fNIRS signals by using multidistance optodes. Neurophotonics 2015, 2, 15003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Bray, S.; Bryant, D.M.; Glover, G.H.; Reiss, A.L. A quantitative comparison of NIRS and fMRI across multiple cognitive tasks. Neuroimage 2011, 54, 2808–2821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witmer, J.S.; Aeschlimann, E.A.; Metz, A.J.; Troche, S.J.; Rammsayer, T.H. The validity of functional near-infrared spectroscopy recordings of visuospatial working memory processes in humans. Brain Sci. 2018, 8, 62. [Google Scholar] [CrossRef] [PubMed]
- Du Boisgueheneuc, F.; Levy, R.; Volle, E.; Seassau, M.; Duffau, H.; Kinkingnehun, S.; Samson, Y.; Zhang, S.; Dubois, B. Functions of the left superior frontal gyrus in humans: A lesion study. Brain 2006, 129, 3315–3328. [Google Scholar] [CrossRef] [PubMed]
- Rottschy, C.; Langner, R.; Dogan, I.; Reetz, K.; Laird, A.R.; Schulz, J.B.; Fox, P.T.; Eickhoff, S.B. Modelling neural correlates of working memory: A coordinate-based meta-analysis. Neuroimage 2012, 60, 830–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasdzewski, G.; Strangman, G.; Wagner, J.; Kwong, K.K.; Poldrack, R.A.; Boas, D.A. Differences in the hemodynamic response to event-related motor and visual paradigms as measured by near-infrared spectroscopy. Neuroimage 2003, 20, 479–488. [Google Scholar] [CrossRef]
- Diener, E.; Biswas-Diener, R. The replication crisis in psychology. In Noba textbook series: Psychology; Biwas-Diener, R., Diener, E., Eds.; DEF Publishers: Champaign, IL, USA, 2017. [Google Scholar]
- Maxwell, S.E.; Lau, M.Y.; Howard, G.S. Is psychology suffering from a replication crisis? What does “failure to replicate” really mean? Am. Psychol. 2015, 70, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Open Science Collaboration Estimating the reproducibility of psychological science. Science 2015. [CrossRef]
- Simons, D.J. The value of direct replication. Perspect. Psychol. Sci. 2014, 9, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, M.S.; Beurlot, M.R.; Fava, E.; Nath, A.R.; Parikh, N.A.; Saad, Z.S.; Bortfeld, H.; Oghalai, J.S. The developmental trajectory of brain-scalp distance from birth through childhood: Implications for functional neuroimaging. PLoS ONE 2011, 6, e24981. [Google Scholar] [CrossRef] [PubMed]
- Strangman, G.E.; Zhang, Q.; Li, Z. Scalp and skull influence on near infrared photon propagation in the Colin27 brain template. Neuroimage 2014, 85, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Charlot, V.; Tzourio, N.; Zilbovicius, M.; Mazoyer, B.; Denis, M. Different mental imagery abilities result in different regional cerebrral blood flow activation patterns during cognitive tasks. Neuropsychologia 1992, 30, 565–580. [Google Scholar] [CrossRef]
- Haier, R.J.; Siegel, B.V.; Nuechterlein, K.H.; Hazlett, E.; Wu, J.C.; Paek, J.; Browning, H.L.; Buchsbaum, M.S. Cortical glucose metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography. Intelligence 1988, 12, 199–217. [Google Scholar] [CrossRef]
- Parks, R.W.; Loewenstein, D.A.; Dodrill, K.L.; Barker, W.W.; Yoshii, F.; Chang, J.Y.; Emran, A.; Apicella, A.; Sheramata, W.A.; Duara, R. Cerebral metabolic effects of a verbal fluency test: A PET scan study. J. Clin. Exp. Neuropsychol. 1988, 10, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.Y.; Eaves, E.L.; Ng, J.C.; Carpenter, D.M.; Mai, X.; Schroeder, D.H.; Condon, C.A.; Colom, R.; Haier, R.J. Brain networks for working memory and factors of intelligence assessed in males and females with fMRI and DTI. Intelligence 2010, 38, 293–303. [Google Scholar] [CrossRef]
- Dunst, B.; Benedek, M.; Jauk, E.; Bergner, S.; Koschutnig, K.; Sommer, M.; Ischebeck, A.; Spinath, B.; Arendasy, M.; Bühner, M.; et al. Neural efficiency as a function of task demands. Intelligence 2014, 42, 22–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preusse, F.; van der Meer, E.; Deshpande, G.; Krueger, F.; Wartenburger, I. Fluid intelligence allows flexible recruitment of the parieto-frontal network in analogical reasoning. Front. Hum. Neurosci. 2011, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Rypma, B.; Berger, J.S.; Prabhakaran, V.; Bly, B.M.; Kimberg, D.Y.; Biswal, B.B.; D’Esposito, M. Neural correlates of cognitive efficiency. Neuroimage 2006, 33, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.R.; Chabris, C.F.; Braver, T.S. Neural mechanisms of general fluid intelligence. Nat. Neurosci. 2003, 6, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, S.I.; Rodrigo, A.H.; Ayaz, H.; Fournier, M.A.; Ruocco, A.C. Decision-making conflict and the neural efficiency hypothesis of intelligence: A functional near-infrared spectroscopy investigation. Neuroimage 2015, 109, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Weiß, R.H. CFT 20-R—Grundintelligenztest Skala 2; Hogrefe: Göttingen, Germany, 2006. [Google Scholar]
- Cattell, R.B. Theory of fluid and crystallized intelligence: A critical experiment. J. Educ. Psychol. 1963, 54. [Google Scholar] [CrossRef]
- Ang, S.Y.; Lee, K. Exploring developmental differences in visual short-term memory and working memory. Dev. Psychol. 2010, 46, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Hasselhorn, M.; Schumann-Hengsteler, R.; Gronauer, J.; Grube, D.; Mähler, C.; Schmid, I.; Seitz-Stein, K.; Zoelch, C. Arbeitsgedächtnistestbatterie für Kinder von Fünf Bis Zwölf Jahren (AGTB 5-12); Hogrefe: Göttingen, Germany, 2012. [Google Scholar]
- Friston, K.J.; Price, C.J.; Fletcher, P.; Moore, C.; Frackowiak, R.S.J.; Dolan, R.J. The trouble with cognitive subtraction. Neuroimage 1996, 4, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Jonides, J.; Marshuetz, C.; Koeppe, R.A. Components of verbal working memory: Evidence from neuroimaging. Proc. Natl. Acad. Sci. USA 1998, 95, 876–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yücel, M.A.; Selb, J.; Aasted, C.M.; Petkov, M.P.; Becerra, L.; Borsook, D.; Boas, D.A. Short separation regression improves statistical significance and better localizes the hemodynamic response obtained by near-infrared spectroscopy for tasks with differing autonomic responses. Neurophotonics 2015, 2, 35005. [Google Scholar] [CrossRef] [PubMed]
- Saager, R.B.; Berger, A.J. Direct characterization and removal of interfering absorption trends in two-layer turbid media. J. Opt. Soc. Am. A. Opt. Image Sci. Vis. 2005, 22, 1874–1882. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, L.; Yücel, M.A.; Boas, D.A.; Cooper, R.J. Further improvement in reducing superficial contamination in NIRS using double short separation measurements. Neuroimage 2014, 85, 127–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholkmann, F.; Spichtig, S.; Muehlemann, T.; Wolf, M. How to detect and reduce movement artifacts in near-infrared imaging using moving standard deviation and spline interpolation. Physiol. Meas. 2010, 31, 649–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnon, L.; Perdue, K.; Greve, D.N.; Goldenholz, D.; Kaskhedikar, G.; Boas, D.A. Improved recovery of the hemodynamic response in diffuse optical imaging using short optode separations and state-space modeling. Neuroimage 2011, 56, 1362–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team R: A language and environment for statistical computing. 2017.
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Zimeo Morais, G.A.; Scholkmann, F.; Balardin, J.B.; Furucho, R.A.; de Paula, R.C.V.; Biazoli, C.E.; Sato, J.R. Non-neuronal evoked and spontaneous hemodynamic changes in the anterior temporal region of the human head may lead to misinterpretations of functional near-infrared spectroscopy signals. Neurophotonics 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Buss, A.T.; Fox, N.; Boas, D.A.; Spencer, J.P. Probing the early development of visual working memory capacity with functional near-infrared spectroscopy. Neuroimage 2014, 85, 314–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, Y.; Hiraki, K. Prefrontal cortex and executive function in young children: A review of NIRS studies. Front. Hum. Neurosci. 2013, 7, 867. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, S.; Yamamoto, T. Prefrontal cortical activation associated with working memory in adults and preschool children: An event-related optical topography study. Cereb. Cortex 2004, 14, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Dehaes, M.; Grant, P.E.; Sliva, D.D.; Roche-Labarbe, N.; Pienaar, R.; Boas, D.A.; Franceschini, M.A.; Selb, J. Assessment of the frequency-domain multi-distance method to evaluate the brain optical properties: Monte Carlo simulations from neonate to adult. Biomed. Opt. Express 2011, 2, 552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, E.; Delpy, D.T. Near-infrared light propagation in an adult head model II. Effect of superficial tissue thickness on the sensitivity of the near-infrared spectroscopy signal. Appl. Opt. 2003, 42, 2915. [Google Scholar] [CrossRef] [PubMed]
- Casey, B.J.; Giedd, J.N.; Thomas, K.M. Structural and functional brain development and its relation to cognitive development. Biol. Psychol. 2000, 54, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Casey, B.J.; Tottenham, N.; Liston, C.; Durston, S. Imaging the developing brain: What have we learned about cognitive development? Trends Cogn. Sci. 2005, 9, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.M.; King, S.W.; Franzen, P.L.; Welsh, T.F.; Berkowitz, A.L.; Noll, D.C.; Birmaher, V.; Casey, B.J. A developmental functional MRI study of spatial working memory. Neuroimage 1999, 10, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Aberson, C. Interpreting null results: Improving presentation and conclusions with confidence intervals 1. J. Artic. Support Null Hypothesis 2002, 1, 36–42. [Google Scholar]
- Nambu, I.; Ozawa, T.; Sato, T.; Aihara, T.; Fujiwara, Y.; Otaka, Y.; Osu, R.; Izawa, J.; Wada, Y. Transient increase in systemic interferences in the superficial layer and its influence on event-related motor tasks: A functional near-infrared spectroscopy study. J. Biomed. Opt. 2017, 22, 35008. [Google Scholar] [CrossRef] [PubMed]
- Scholkmann, F.; Kleiser, S.; Metz, A.J.; Zimmermann, R.; Mata Pavia, J.; Wolf, U.; Wolf, M. A review on continuous wave functional near-infrared spectroscopy and imaging instrumentation and methodology. Neuroimage 2014, 85, 6–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, M.D.; Scholkmann, F.; Labruyère, R. Signal processing in functional near-infrared spectroscopy (fNIRS): Methodological differences lead to different statistical results. Front. Hum. Neurosci. 2018, 11, 641. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, F.; Brigadoi, S.; Cutini, S.; Scatturin, P.; Zorzi, M.; Dell’Acqua, R.; Sparacino, G. A reference-channel based methodology to improve estimation of event-related hemodynamic response from fNIRS measurements. Neuroimage 2013, 72, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Scholkmann, F.; Metz, A.J.; Wolf, M. Measuring tissue hemodynamics and oxygenation by continuous-wave functional near-infrared spectroscopy—How robust are the different calculation methods against movement artifacts? Physiol. Meas. 2014, 35, 717–734. [Google Scholar] [CrossRef] [PubMed]
- Bartholomeusz, H.H.; Courchesne, E.; Karns, C.M. Relationship between head circumference and brain volume in healthy normal toddllers, children, and adults. Neuropediatrics 2002, 33, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Okamoto, M.; Dan, H.; Jurcak, V.; Dan, I. Spatial registration of multichannel multi-subject fNIRS data to MNI space without MRI. Neuroimage 2005, 27, 842–851. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hemoglobin Oxygenation | Condition | Total | LA | HA | |||
|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | ||
| O2Hb | control | 0.23 | 0.54 | 0.15 | 0.59 | 0.29 | 0.49 |
| experimental | 0.40 | 0.53 | 0.54 | 0.57 | 0.29 | 0.48 | |
| HHb | control | −0.01 | 0.22 | −0.03 | 0.25 | 0.01 | 0.19 |
| experimental | −0.05 | 0.23 | −0.08 | 0.23 | −0.03 | 0.23 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witmer, J.S.; Aeschlimann, E.A.; Metz, A.J.; Troche, S.J.; Rammsayer, T.H. Functional Near-Infrared Spectroscopy Recordings of Visuospatial Working Memory Processes. Part II: A Replication Study in Children on Sensitivity and Mental-Ability-Induced Differences in Functional Activation. Brain Sci. 2018, 8, 152. https://doi.org/10.3390/brainsci8080152
Witmer JS, Aeschlimann EA, Metz AJ, Troche SJ, Rammsayer TH. Functional Near-Infrared Spectroscopy Recordings of Visuospatial Working Memory Processes. Part II: A Replication Study in Children on Sensitivity and Mental-Ability-Induced Differences in Functional Activation. Brain Sciences. 2018; 8(8):152. https://doi.org/10.3390/brainsci8080152
Chicago/Turabian StyleWitmer, Joëlle S., Eva A. Aeschlimann, Andreas J. Metz, Stefan J. Troche, and Thomas H. Rammsayer. 2018. "Functional Near-Infrared Spectroscopy Recordings of Visuospatial Working Memory Processes. Part II: A Replication Study in Children on Sensitivity and Mental-Ability-Induced Differences in Functional Activation" Brain Sciences 8, no. 8: 152. https://doi.org/10.3390/brainsci8080152
APA StyleWitmer, J. S., Aeschlimann, E. A., Metz, A. J., Troche, S. J., & Rammsayer, T. H. (2018). Functional Near-Infrared Spectroscopy Recordings of Visuospatial Working Memory Processes. Part II: A Replication Study in Children on Sensitivity and Mental-Ability-Induced Differences in Functional Activation. Brain Sciences, 8(8), 152. https://doi.org/10.3390/brainsci8080152