
Protective and Regenerative Effects of Reconstituted HDL on Human Rotator Cuff Fibroblasts under Hypoxia: An In Vitro Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Sample Collection
2.2. Reagents
2.3. Study Design
2.4. Measurement of Intracellular ROS Production
2.5. Western Blot Analyses
2.6. Cell Viability Analyses
2.7. Analyses for Apoptosis Rates
2.8. Analyses of Cellular Proliferation
2.9. Statistical Analysis
3. Results
3.1. Rates of Intracellular ROS Production
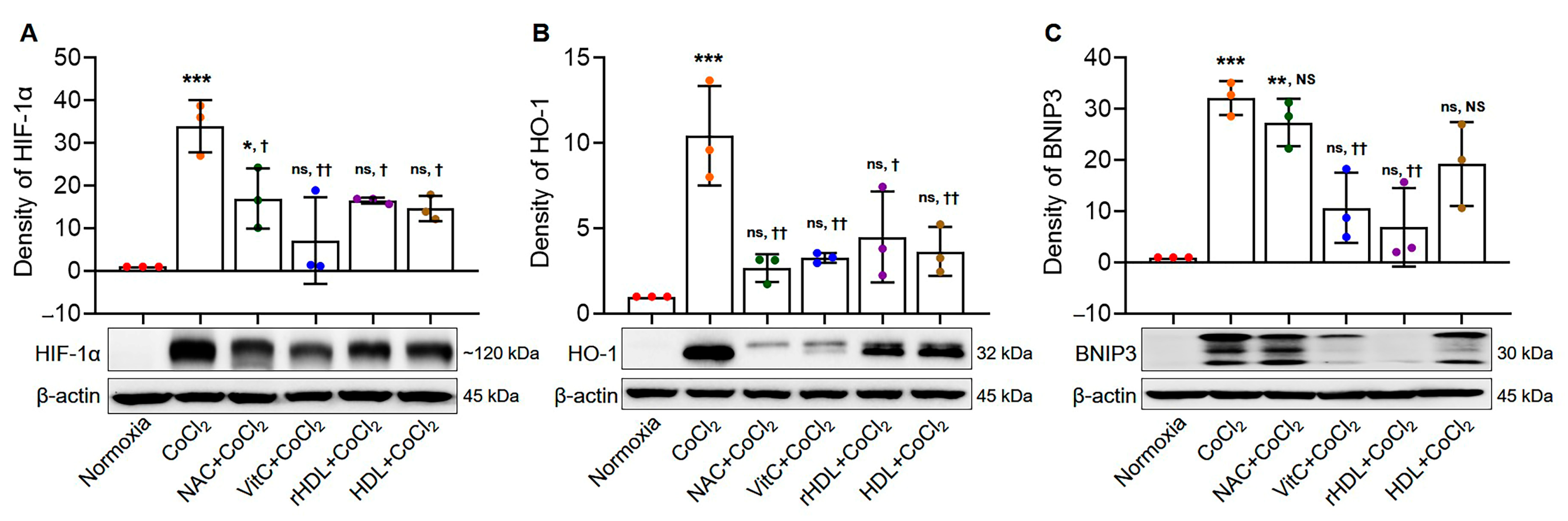
3.2. Analyses of the Expressions of HIF-1α, HO-1, and BNIP3
3.3. Cell viability Analyses
3.4. Analyses for Apoptosis
3.5. Analysis of VEGF and MMP-2 Expressions
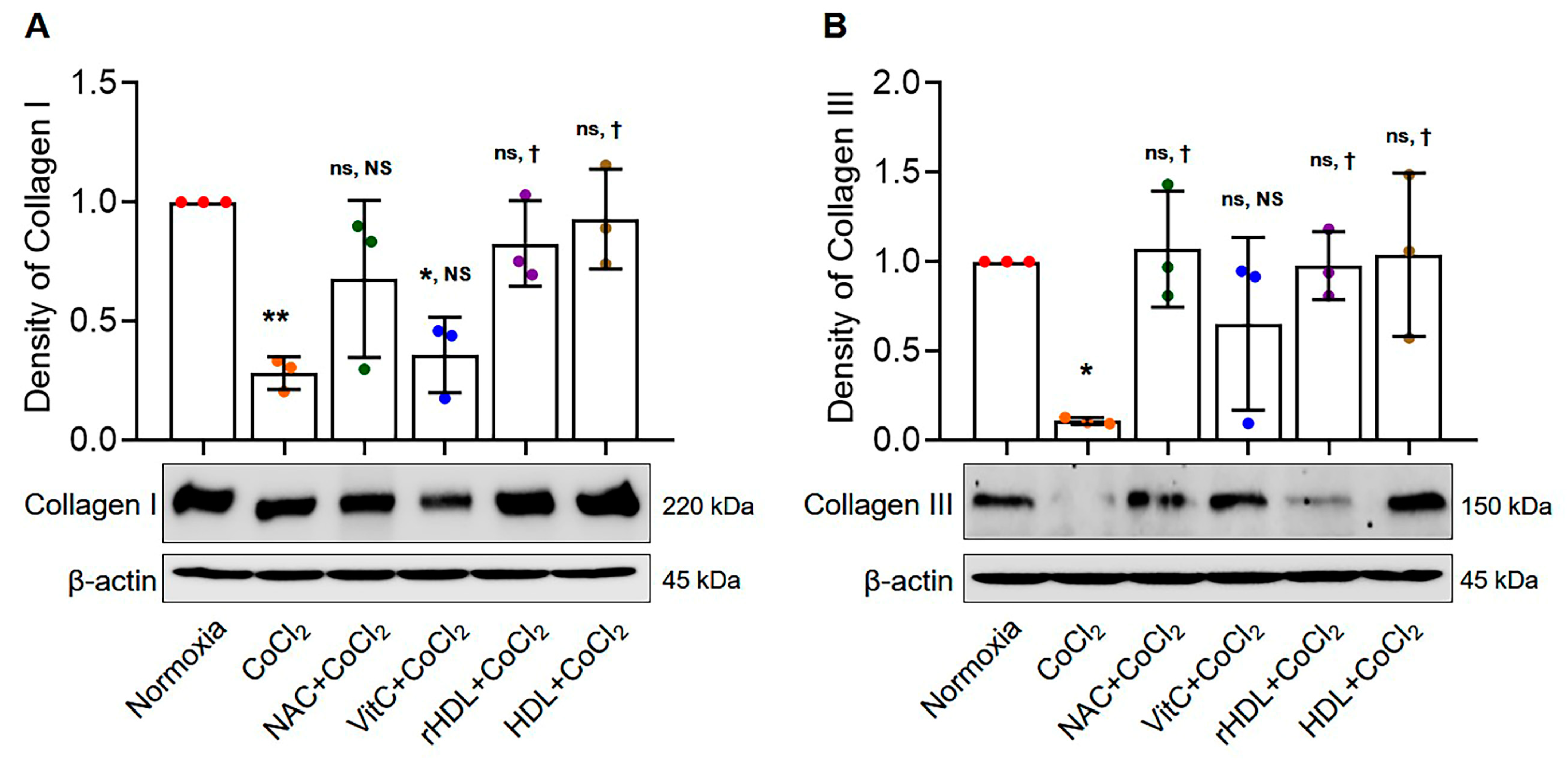
3.6. Analyses of Collagen I and III Production
3.7. Analyses of Cell Proliferation Abilities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BNIP3 | Bcl-2/E1B-19kDa interacting protein 3 |
| DAPI | 4′, 6-diamidino-2-phenylindole |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| HDL | high-density lipoprotein |
| HIF-1α | hypoxia-inducible factor-1α |
| HO-1 | heme oxygenase-1 |
| Hypo-HDLemia | hypo-high-density lipoproteinemia |
| MMP-2 | matrix metalloproteinase-2 |
| NAC | N-acetylcysteine |
| PARP-1 | poly ADP-ribose polymerase-1 |
| RCFs | rotator cuff fibroblasts |
| rHDL | reconstituted HDL |
| ROS | intracellular reactive oxygen species |
| VEGF | vascular endothelial growth factors |
| Vit C | vitamin C |
References
- Park, H.B.; Gwark, J.Y.; Im, J.H.; Jung, J.; Na, J.B.; Yoon, C.H. Factors Associated with Atraumatic Posterosuperior Rotator Cuff Tears. J. Bone Jt. Surg. Am. 2018, 100, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.E.; Bjornsson Hallgren, H.C.; Lunsjo, K.; Frobell, R. No differences in histopathological degenerative changes found in acute, trauma-related rotator cuff tears compared with chronic, nontraumatic tears. Knee Surg. Sports Traumatol. Arthrosc. 2022, 30, 2521–2527. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Griffin, D.R.; Parsons, N.; Lawrence, T.M.; Modi, C.S.; Drew, S.J.; Smith, C.D. Microvascular blood flow in normal and pathologic rotator cuffs. J. Shoulder Elbow Surg. 2015, 24, 1954–1960. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, E.J.; Cook, C.; Brennan, M.; Wyland, D.; Garrison, J.C.; Driesner, D. Vascularity and tendon pathology in the rotator cuff: A review of literature and implications for rehabilitation and surgery. Br. J. Sports Med. 2010, 44, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.F.; Uhthoff, H.K. The microvascular pattern of the supraspinatus tendon. Clin. Orthop. Relat. Res. 1990, 254, 35–38. [Google Scholar] [CrossRef]
- Funakoshi, T.; Iwasaki, N.; Kamishima, T.; Nishida, M.; Ito, Y.; Kondo, M.; Minami, A. In vivo visualization of vascular patterns of rotator cuff tears using contrast-enhanced ultrasound. Am. J. Sports Med. 2010, 38, 2464–2471. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Murrell, G.A.; Wei, A.Q.; Wang, M.X. Apoptosis in rotator cuff tendonopathy. J. Orthop. Res. 2002, 20, 1372–1379. [Google Scholar] [CrossRef] [PubMed]
- Lakemeier, S.; Reichelt, J.J.; Patzer, T.; Fuchs-Winkelmann, S.; Paletta, J.R.; Schofer, M.D. The association between retraction of the torn rotator cuff and increasing expression of hypoxia inducible factor 1alpha and vascular endothelial growth factor expression: An immunohistological study. BMC Musculoskelet. Disord. 2010, 11, 230. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, Y.S.; Ok, J.H.; Song, H.J. Apoptosis occurs throughout the diseased rotator cuff. Am. J. Sports Med. 2013, 41, 2249–2255. [Google Scholar] [CrossRef]
- Pufe, T.; Petersen, W.J.; Mentlein, R.; Tillmann, B.N. The role of vasculature and angiogenesis for the pathogenesis of degenerative tendons disease. Scand. J. Med. Sci. Sports 2005, 15, 211–222. [Google Scholar] [CrossRef]
- Fuhrmann, D.C.; Brune, B. Mitochondrial composition and function under the control of hypoxia. Redox Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef] [PubMed]
- McBeath, R.; Edwards, R.W.; O’Hara, B.J.; Maltenfort, M.G.; Parks, S.M.; Steplewski, A.; Osterman, A.L.; Shapiro, I.M. Tendinosis develops from age- and oxygen tension-dependent modulation of Rac1 activity. Aging Cell 2019, 18, e12934. [Google Scholar] [CrossRef] [PubMed]
- Petersen, W.; Varoga, D.; Zantop, T.; Hassenpflug, J.; Mentlein, R.; Pufe, T. Cyclic strain influences the expression of the vascular endothelial growth factor (VEGF) and the hypoxia inducible factor 1 alpha (HIF-1alpha) in tendon fibroblasts. J. Orthop. Res. 2004, 22, 847–853. [Google Scholar] [CrossRef]
- Benson, R.T.; McDonnell, S.M.; Knowles, H.J.; Rees, J.L.; Carr, A.J.; Hulley, P.A. Tendinopathy and tears of the rotator cuff are associated with hypoxia and apoptosis. J. Bone Jt. Surg. Br. 2010, 92, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Gerber, H.P.; Condorelli, F.; Park, J.; Ferrara, N. Differential transcriptional regulation of the two vascular endothelial growth factor receptor genes. Flt-1, but not Flk-1/KDR, is up-regulated by hypoxia. J. Biol. Chem. 1997, 272, 23659–23667. [Google Scholar] [CrossRef]
- Petersen, W.; Pufe, T.; Kurz, B.; Mentlein, R.; Tillmann, B. Angiogenesis in fetal tendon development: Spatial and temporal expression of the angiogenic peptide vascular endothelial cell growth factor. Anat. Embryol. 2002, 205, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.S.; Raza, S.A.; Pilcher, J.; Heron, C.; Poloniecki, J.D. The prevalence of neovascularity in patients clinically diagnosed with rotator cuff tendinopathy. BMC Musculoskelet. Disord. 2009, 10, 163. [Google Scholar] [CrossRef]
- Abboud, J.A.; Kim, J.S. The effect of hypercholesterolemia on rotator cuff disease. Clin. Orthop. Relat. Res. 2010, 468, 1493–1497. [Google Scholar] [CrossRef]
- Park, H.B.; Gwark, J.Y.; Kwack, B.H.; Jung, J. Hypo-High-Density Lipoproteinemia Is Associated with Preoperative Tear Size and With Postoperative Retear in Large to Massive Rotator Cuff Tears. Arthroscopy 2020, 36, 2071–2079. [Google Scholar] [CrossRef]
- Gordon, D.J.; Rifkind, B.M. High-density lipoprotein--the clinical implications of recent studies. N. Engl. J. Med. 1989, 321, 1311–1316. [Google Scholar] [CrossRef]
- Tan, J.T.; Ng, M.K.; Bursill, C.A. The role of high-density lipoproteins in the regulation of angiogenesis. Cardiovasc. Res. 2015, 106, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Sumi, M.; Sata, M.; Miura, S.; Rye, K.A.; Toya, N.; Kanaoka, Y.; Yanaga, K.; Ohki, T.; Saku, K.; Nagai, R. Reconstituted high-density lipoprotein stimulates differentiation of endothelial progenitor cells and enhances ischemia-induced angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; van Eck, M.; Van Craeyveld, E.; Jacobs, F.; Carlier, V.; Van Linthout, S.; Erdel, M.; Tjwa, M.; De Geest, B. Critical role of scavenger receptor-BI-expressing bone marrow-derived endothelial progenitor cells in the attenuation of allograft vasculopathy after human apo A-I transfer. Blood 2009, 113, 755–764. [Google Scholar] [CrossRef]
- Seetharam, D.; Mineo, C.; Gormley, A.K.; Gibson, L.L.; Vongpatanasin, W.; Chambliss, K.L.; Hahner, L.D.; Cummings, M.L.; Kitchens, R.L.; Marcel, Y.L.; et al. High-density lipoprotein promotes endothelial cell migration and reendothelialization via scavenger receptor-B type I. Circ. Res. 2006, 98, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.T.; Prosser, H.C.; Vanags, L.Z.; Monger, S.A.; Ng, M.K.; Bursill, C.A. High-density lipoproteins augment hypoxia-induced angiogenesis via regulation of post-translational modulation of hypoxia-inducible factor 1alpha. FASEB J. 2014, 28, 206–217. [Google Scholar] [CrossRef]
- Prosser, H.C.; Tan, J.T.; Dunn, L.L.; Patel, S.; Vanags, L.Z.; Bao, S.; Ng, M.K.; Bursill, C.A. Multifunctional regulation of angiogenesis by high-density lipoproteins. Cardiovasc. Res. 2014, 101, 145–154. [Google Scholar] [CrossRef]
- Kim, R.J.; An, S.H.; Gwark, J.Y.; Park, H.B. Antioxidant effects on hypoxia-induced oxidative stress and apoptosis in rat rotator cuff fibroblasts. Eur. Cell Mater. 2021, 41, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Cornell, H.R.; Zargar Baboldashti, N.; Thompson, M.S.; Carr, A.J.; Hulley, P.A. Regulation of hypoxia-induced cell death in human tenocytes. Adv. Orthop. 2012, 2012, 984950. [Google Scholar] [CrossRef]
- Vucic, E.; Rosenson, R.S. Recombinant high-density lipoprotein formulations. Curr. Atheroscler. Rep. 2011, 13, 81–87. [Google Scholar] [CrossRef]
- Cao, Y.N.; Xu, L.; Han, Y.C.; Wang, Y.N.; Liu, G.; Qi, R. Recombinant high-density lipoproteins and their use in cardiovascular diseases. Drug Discov. Today 2017, 22, 180–185. [Google Scholar] [CrossRef]
- Goldberg, M.A.; Dunning, S.P.; Bunn, H.F. Regulation of the erythropoietin gene: Evidence that the oxygen sensor is a heme protein. Science 1988, 242, 1412–1415. [Google Scholar] [CrossRef]
- Abdel-Rahman Mohamed, A.; Metwally, M.M.M.; Khalil, S.R.; Salem, G.A.; Ali, H.A. Moringa oleifera extract attenuates the CoCl(2) induced hypoxia of rat’s brain: Expression pattern of HIF-1alpha, NF-kB, MAO and EPO. Biomed. Pharmacother. 2019, 109, 1688–1697. [Google Scholar] [CrossRef]
- Borcar, A.; Menze, M.A.; Toner, M.; Hand, S.C. Metabolic preconditioning of mammalian cells: Mimetic agents for hypoxia lack fidelity in promoting phosphorylation of pyruvate dehydrogenase. Cell Tissue Res. 2013, 351, 99–106. [Google Scholar] [CrossRef]
- Huang, Y.J.; Nan, G.X. Oxidative stress-induced angiogenesis. J. Clin. Neurosci. 2019, 63, 13–16. [Google Scholar] [CrossRef]
- Lendahl, U.; Lee, K.L.; Yang, H.; Poellinger, L. Generating specificity and diversity in the transcriptional response to hypoxia. Nat. Rev. Genet. 2009, 10, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Rajagopal, V.; Gonsalves, C.; Johnson, C.; Kalra, V.K. A novel role of hypoxia-inducible factor in cobalt chloride- and hypoxia-mediated expression of IL-8 chemokine in human endothelial cells. J. Immunol. 2006, 177, 7211–7224. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, A.; Liakos, P.; Tsakalof, A.; Georgatsou, E.; Simos, G.; Bonanou, S. Cobalt induces hypoxia-inducible factor-1alpha (HIF-1alpha) in HeLa cells by an iron-independent, but ROS-, PI-3K- and MAPK-dependent mechanism. Free Radic. Res. 2006, 40, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Amersi, F.; Buelow, R.; Kato, H.; Ke, B.; Coito, A.J.; Shen, X.D.; Zhao, D.; Zaky, J.; Melinek, J.; Lassman, C.R.; et al. Upregulation of heme oxygenase-1 protects genetically fat Zucker rat livers from ischemia/reperfusion injury. J. Clin. Investig. 1999, 104, 1631–1639. [Google Scholar] [CrossRef]
- Taketani, S.; Kohno, H.; Yoshinaga, T.; Tokunaga, R. Induction of heme oxygenase in rat hepatoma cells by exposure to heavy metals and hyperthermia. Biochem. Int. 1988, 17, 665–672. [Google Scholar]
- Wang, G.L.; Jiang, B.H.; Rue, E.A.; Semenza, G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 1995, 92, 5510–5514. [Google Scholar] [CrossRef]
- Prabhakaran, K.; Li, L.; Zhang, L.; Borowitz, J.L.; Isom, G.E. Upregulation of BNIP3 and translocation to mitochondria mediates cyanide-induced apoptosis in cortical cells. Neuroscience 2007, 150, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Kang, Y. Hypoxia and hypoxia-inducible factors: Master regulators of metastasis. Clin. Cancer Res. 2010, 16, 5928–5935. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.V.; Sapochnik, D.; Garcia Sola, M.; Coso, O. Regulation of the Expression of Heme Oxygenase-1: Signal Transduction, Gene Promoter Activation, and Beyond. Antioxid. Redox Signal 2020, 32, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Azenshtein, E.; Meshel, T.; Shina, S.; Barak, N.; Keydar, I.; Ben-Baruch, A. The angiogenic factors CXCL8 and VEGF in breast cancer: Regulation by an array of pro-malignancy factors. Cancer Lett. 2005, 217, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Zhang, H.; Dinavahi, R.; Li, F.; Xiang, Y.; Raman, V.; Bhujwalla, Z.M.; Felsher, D.W.; Cheng, L.; Pevsner, J.; et al. HIF-dependent antitumorigenic effect of antioxidants in vivo. Cancer Cell 2007, 12, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Sceneay, J.; Liu, M.C.; Chen, A.; Wong, C.S.; Bowtell, D.D.; Moller, A. The antioxidant N-acetylcysteine prevents HIF-1 stabilization under hypoxia in vitro but does not affect tumorigenesis in multiple breast cancer models in vivo. PLoS ONE 2013, 8, e66388. [Google Scholar] [CrossRef] [PubMed]
- Motterlini, R.; Foresti, R.; Bassi, R.; Calabrese, V.; Clark, J.E.; Green, C.J. Endothelial heme oxygenase-1 induction by hypoxia. Modulation by inducible nitric-oxide synthase and S-nitrosothiols. J. Biol. Chem. 2000, 275, 13613–13620. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tan, P.; Zhou, W.; Zhu, X.; Cui, Y.; Zhu, L.; Feng, X.; Qi, H.; Zheng, J.; Gu, P.; et al. N-acetylcysteine protects against hypoxia mimetic-induced autophagy by targeting the HIF-1alpha pathway in retinal ganglion cells. Cell Mol. Neurobiol. 2012, 32, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Aitio, M.L. N-acetylcysteine -- passe-partout or much ado about nothing? Br. J. Clin. Pharmacol. 2006, 61, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Guzy, R.D.; Schumacker, P.T. Oxygen sensing by mitochondria at complex III: The paradox of increased reactive oxygen species during hypoxia. Exp. Physiol. 2006, 91, 807–819. [Google Scholar] [CrossRef]
- Kotake-Nara, E.; Saida, K. Endothelin-2/vasoactive intestinal contractor: Regulation of expression via reactive oxygen species induced by CoCl2, and Biological activities including neurite outgrowth in PC12 cells. ScientificWorldJournal 2006, 6, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Yan, M.; Xu, W.; Huo, H.; Sun, L.; Zheng, Z.; Liu, X. Cobalt chloride induces PC12 cells apoptosis through reactive oxygen species and accompanied by AP-1 activation. J. Neurosci. Res. 2001, 64, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.C.; Zhou, W.; Shu, R.; Ni, J. Hypoxia induces apoptosis and autophagic cell death in human periodontal ligament cells through HIF-1alpha pathway. Cell Prolif. 2012, 45, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Dalgard, C.L.; Mohyeldin, A.; McFate, T.; Tait, A.S.; Verma, A. Reversible inactivation of HIF-1 prolyl hydroxylases allows cell metabolism to control basal HIF-1. J. Biol. Chem. 2005, 280, 41928–41939. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed]
- Tewari, M.; Quan, L.T.; O’Rourke, K.; Desnoyers, S.; Zeng, Z.; Beidler, D.R.; Poirier, G.G.; Salvesen, G.S.; Dixit, V.M. Yama/CPP32 beta, a mammalian homolog of CED-3, is a CrmA-inhibitable protease that cleaves the death substrate poly(ADP-ribose) polymerase. Cell 1995, 81, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Li, J.; Li, L.; Sun, F.; Xie, J. Effects of hypoxia ischemia on caspase-3 expression and neuronal apoptosis in the brain of neonatal mice. Exp. Ther. Med. 2019, 17, 4517–4521. [Google Scholar] [CrossRef] [PubMed]
- Khurana, P.; Ashraf, Q.M.; Mishra, O.P.; Delivoria-Papadopoulos, M. Effect of hypoxia on caspase-3, -8, and -9 activity and expression in the cerebral cortex of newborn piglets. Neurochem. Res. 2002, 27, 931–938. [Google Scholar] [CrossRef]
- Marti, J.M.; Garcia-Diaz, A.; Delgado-Bellido, D.; O’Valle, F.; Gonzalez-Flores, A.; Carlevaris, O.; Rodriguez-Vargas, J.M.; Ame, J.C.; Dantzer, F.; King, G.L.; et al. Selective modulation by PARP-1 of HIF-1alpha-recruitment to chromatin during hypoxia is required for tumor adaptation to hypoxic conditions. Redox Biol. 2021, 41, 101885. [Google Scholar] [CrossRef]
- Mucci, S.; Isaja, L.; Rodriguez-Varela, M.S.; Ferriol-Laffouillere, S.L.; Marazita, M.; Videla-Richardson, G.A.; Sevlever, G.E.; Scassa, M.E.; Romorini, L. Acute severe hypoxia induces apoptosis of human pluripotent stem cells by a HIF-1alpha and P53 independent mechanism. Sci. Rep. 2022, 12, 18803. [Google Scholar] [CrossRef]
- Kim, Y.W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Dulak, J.; Deshane, J.; Jozkowicz, A.; Agarwal, A. Heme oxygenase-1 and carbon monoxide in vascular pathobiology: Focus on angiogenesis. Circulation 2008, 117, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Dulak, J.; Loboda, A.; Jozkowicz, A. Effect of heme oxygenase-1 on vascular function and disease. Curr. Opin. Lipidol. 2008, 19, 505–512. [Google Scholar] [CrossRef]
- Sadaghianloo, N.; Yamamoto, K.; Bai, H.; Tsuneki, M.; Protack, C.D.; Hall, M.R.; Declemy, S.; Hassen-Khodja, R.; Madri, J.; Dardik, A. Increased Oxidative Stress and Hypoxia Inducible Factor-1 Expression during Arteriovenous Fistula Maturation. Ann. Vasc. Surg. 2017, 41, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.S.; Park, H.J.; Hwang, H.R.; Kwon, A.; Lim, W.H.; Yi, W.J.; Han, D.H.; Kim, Y.H.; Baek, J.H. The effect of antioxidants on the production of pro-inflammatory cytokines and orthodontic tooth movement. Mol. Cells 2011, 32, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Antonicelli, F.; Bellon, G.; Debelle, L.; Hornebeck, W. Elastin-elastases and inflamm-aging. Curr. Top. Dev. Biol. 2007, 79, 99–155. [Google Scholar] [CrossRef] [PubMed]
- Fields, G.B. Interstitial collagen catabolism. J. Biol. Chem. 2013, 288, 8785–8793. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.R.; Kondo, S.; Hirose, K.; Ishiguro, N.; Hasegawa, Y.; Iwata, H. Expression and enzymatic activity of MMP-2 during healing process of the acute supraspinatus tendon tear in rabbits. J. Orthop. Res. 2002, 20, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Aimes, R.T.; Quigley, J.P. Matrix metalloproteinase-2 is an interstitial collagenase. Inhibitor-free enzyme catalyzes the cleavage of collagen fibrils and soluble native type I collagen generating the specific 3/4- and 1/4-length fragments. J. Biol. Chem. 1995, 270, 5872–5876. [Google Scholar] [CrossRef]
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef]
- Konttinen, Y.T.; Ceponis, A.; Takagi, M.; Ainola, M.; Sorsa, T.; Sutinen, M.; Salo, T.; Ma, J.; Santavirta, S.; Seiki, M. New collagenolytic enzymes/cascade identified at the pannus-hard tissue junction in rheumatoid arthritis: Destruction from above. Matrix Biol. 1998, 17, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Patterson, M.L.; Atkinson, S.J.; Knauper, V.; Murphy, G. Specific collagenolysis by gelatinase A, MMP-2, is determined by the hemopexin domain and not the fibronectin-like domain. FEBS Lett. 2001, 503, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Riley, G. The pathogenesis of tendinopathy. A molecular perspective. Rheumatology 2004, 43, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, J.; Uhthoff, H.K.; Sarkar, K.; Murnaghan, J.P. Collagen type III in rotator cuff tears: An immunohistochemical study. J. Shoulder Elbow Surg. 1992, 1, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Hirose, K.; Kondo, S.; Choi, H.R.; Mishima, S.; Iwata, H.; Ishiguro, N. Spontaneous healing process of a supraspinatus tendon tear in rabbits. Arch. Orthop. Trauma. Surg. 2004, 124, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Rai, V.; Agrawal, D.K. Regulation of Collagen I and Collagen III in Tissue Injury and Regeneration. Cardiol. Cardiovasc. Med. 2023, 7, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, J.; Wang, J.; Zhang, T.; Xu, D.; Hu, W.; Feng, Z. The Interplay Between Tumor Suppressor p53 and Hypoxia Signaling Pathways in Cancer. Front. Cell Dev. Biol. 2021, 9, 648808. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, R.J.; Park, H.B. Protective and Regenerative Effects of Reconstituted HDL on Human Rotator Cuff Fibroblasts under Hypoxia: An In Vitro Study. Antioxidants 2024, 13, 497. https://doi.org/10.3390/antiox13040497
Kim RJ, Park HB. Protective and Regenerative Effects of Reconstituted HDL on Human Rotator Cuff Fibroblasts under Hypoxia: An In Vitro Study. Antioxidants. 2024; 13(4):497. https://doi.org/10.3390/antiox13040497
Chicago/Turabian StyleKim, Ra Jeong, and Hyung Bin Park. 2024. "Protective and Regenerative Effects of Reconstituted HDL on Human Rotator Cuff Fibroblasts under Hypoxia: An In Vitro Study" Antioxidants 13, no. 4: 497. https://doi.org/10.3390/antiox13040497
APA StyleKim, R. J., & Park, H. B. (2024). Protective and Regenerative Effects of Reconstituted HDL on Human Rotator Cuff Fibroblasts under Hypoxia: An In Vitro Study. Antioxidants, 13(4), 497. https://doi.org/10.3390/antiox13040497