

Dietary Lysophosphatidylcholine Improves the Uptake of Astaxanthin and Modulates Cholesterol Transport in Pacific White Shrimp Litopenaeus vannamei



 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets and Feeding Trial
2.2. Experimental Shrimp and Feeding Management
2.3. Sample Collection
2.4. Analysis of the Proximate Composition and Fatty Acid Composition
2.5. Analysis of Astaxanthin Concentration
2.6. Biochemical Parameters in Serum and Hepatopancreas
2.7. Analysis of Body Color
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.9. Statistical Methods
3. Results
3.1. Growth Performances and Somatic Indices
3.2. Body Proximate Composition
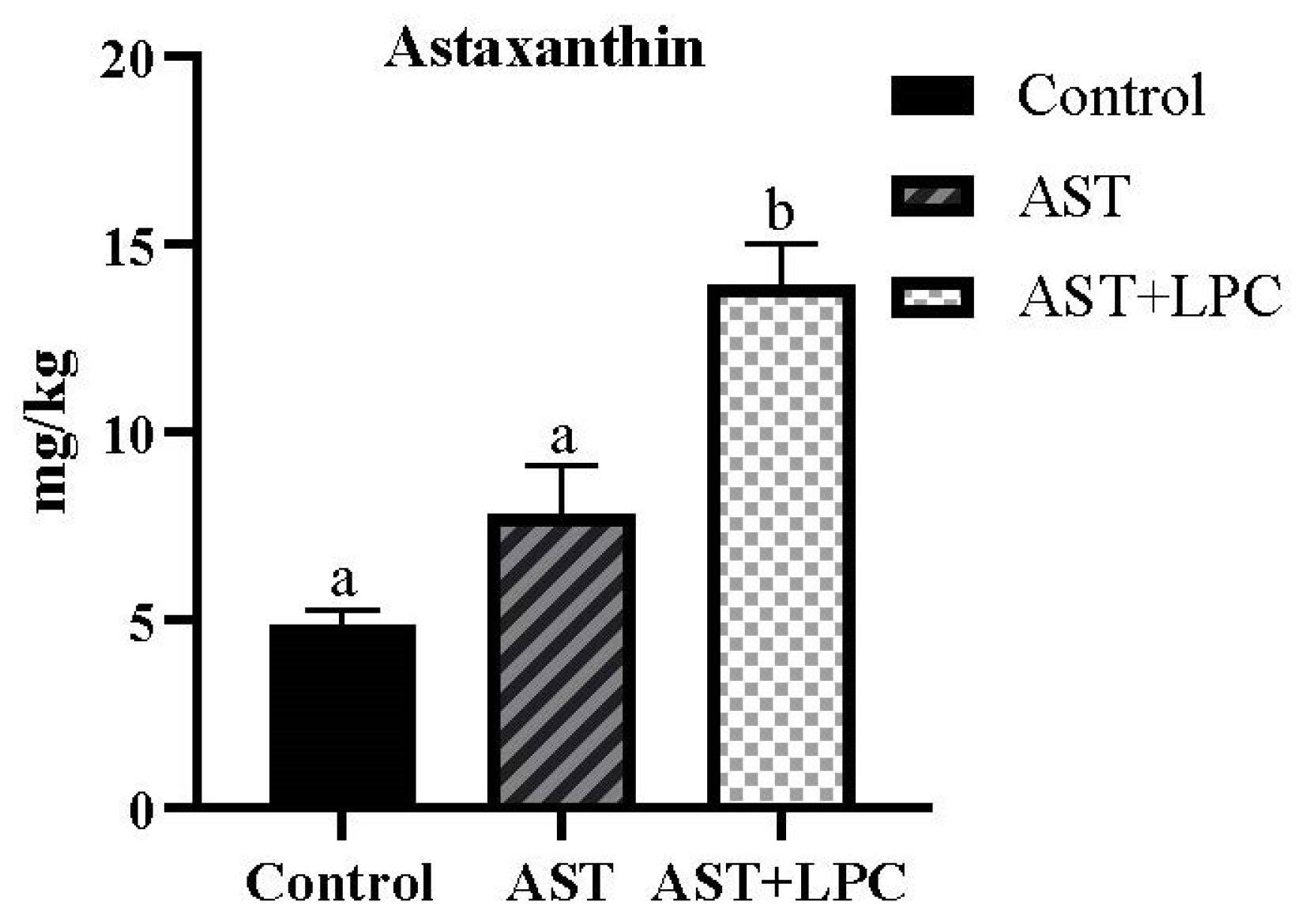
3.3. Muscle Astaxanthin Concentration
3.4. Body Color
3.5. Fatty Acid Profiles of the Hepatopancreas
3.6. Biochemical Parameters in Serum and Hepatopancreas
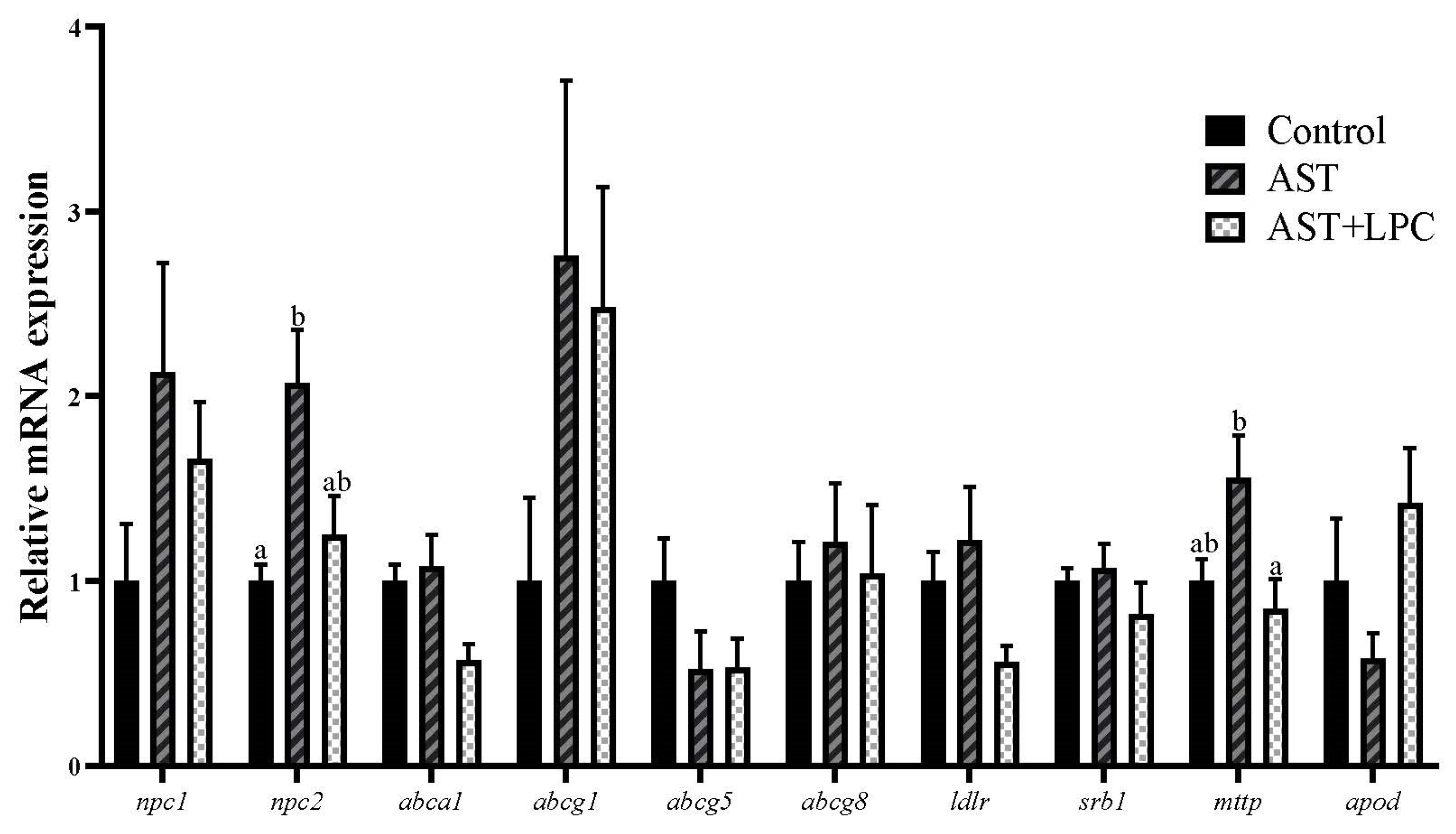
3.7. mRNA Expression of Genes Related to Lipid and Cholesterol Transport
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.K.; Mitra, A.; Rahimnejad, S.; Chi, S.Y.; Kumar, V.; Tan, B.P.; Niu, J.; Xie, S.W. Retrospect of fish meal substitution in Pacific white shrimp (Litopenaeus vannamei) feed: Alternatives, limitations and future prospects. Rev. Aquac. 2023, 16, 382–409. [Google Scholar] [CrossRef]
- Nishida, Y.; Berg, P.C.; Shakersain, B.; Hecht, K.; Takikawa, A.; Tao, R.; Kakuta, Y.; Uragami, C.; Hashimoto, H.; Misawa, N. Astaxanthin: Past, present, and future. Mar. Drugs 2023, 21, 514. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, T. Aquatic animal carotenoids. Fish. Sci. 2001, 67, 5. [Google Scholar] [CrossRef]
- Niu, J.; Tian, L.X.; Liu, Y.J.; Yang, H.J.; Ye, C.X.; Gao, W.; Mai, K.S. Effect of dietary astaxanthin on growth, survival, and stress tolerance of postlarval shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2009, 40, 795–802. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.J.; Tian, L.X.; Yang, H.J.; Liang, G.Y.; Yue, Y.R.; Xu, D.H. Effects of dietary astaxanthin on growth, antioxidant capacity and gene expression in Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2013, 19, 917–927. [Google Scholar] [CrossRef]
- Wang, W.L.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Dawood, M.A.O.; Hossain, M.S.; Zaineldin, A.I. Interactive effects of dietary astaxanthin and cholesterol on the growth, pigmentation, fatty acid analysis, immune response and stress resistance of kuruma shrimp (Marsupenaeus japonicus). Aquac. Nutr. 2019, 25, 946–958. [Google Scholar] [CrossRef]
- Nguyen, K.D. Astaxanthin: A Comparative Case of Synthetic vs. Natural Production; University of Tennessee: Knoxville, TN, USA, 2013. [Google Scholar]
- Xu, H.G.; Luo, X.; Bi, Q.Z.; Wang, Z.D.; Meng, X.X.; Liu, J.S.; Duan, M.; Wei, Y.L.; Liang, M.Q. Effects of dietary lysophosphatidylcholine on growth performance and lipid metabolism of juvenile turbot. Aquac. Nutr. 2022, 2022, 3515101. [Google Scholar] [CrossRef]
- Cai, M.L. Effects of Dietary Lysolecithin on Growth, Lipid Metabolism and Muscle Quality of Red Swamp Crayfish (Procambarus clarkii). Master’s Thesis, Hunan Agricultural University, Changsha, China, 2021. [Google Scholar] [CrossRef]
- Taghavizadeh, M.; Shekarabi, H.P.S.; Mehrgan, S.M.; Islami, H.R. Efficacy of dietary lysophospholipids (Lipidol™) on growth performance, serum immuno-biochemical parameters, and the expression of immune and antioxidant-related genes in rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 525, 735315. [Google Scholar] [CrossRef]
- Liu, G.X.; Ma, S.L.; Chen, F.Y.; Zhang, W.B.; Mai, K.S. Effects of dietary lysolecithin on growth performance, feed utilization, intestinal morphology and metabolic responses of channel catfish (Ictalurus punctatus). Aquac. Nutr. 2020, 26, 456–465. [Google Scholar] [CrossRef]
- Eldessouki, E.A.A.; Diab, A.M.; Selema, T.A.M.A.; Sabry, N.M.; Abotaleb, M.M.; Khalil, R.H.; Abdel, T.M. Dietary astaxanthin modulated the performance, gastrointestinal histology, and antioxidant and immune responses and enhanced the resistance of Litopenaeus vannamei against Vibrio harveyi infection. Aquac. Int. 2022, 30, 1869–1887. [Google Scholar] [CrossRef]
- Samia, F.; Wang, W.L.; Zhou, Y.; Xue, Y.C.; Yi, G.F.; Wu, M.Q.; Huang, X.X. Can dietary β-carotene supplementation provide an alternative to astaxanthin on the performance of growth, pigmentation, biochemical, and immuno-physiological parameters of Litopenaeus vannamei? Aquac. Rep. 2022, 23, 101054. [Google Scholar] [CrossRef]
- Folch, J.; Lee, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.X.; Bi, Q.Z.; Cao, L.; Ma, Q.; Wei, Y.L.; Duan, M.; Liang, M.Q.; Xu, H.G. Evaluation of necessity of cholesterol supplementation in diets of two marine teleosts, turbot (Scophthalmus maximus) and tiger puffer (Takifugu rubripes): Effects on growth and lipid metabolism. Aquac. Nutr. 2022, 2022, 4160991. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real–time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.B.; Sun, Z.Y.; Bi, Q.Z.; Gong, Q.L.; Sun, B.; Wei, Y.L.; Liang, M.Q.; Xu, H.G. Screening of reference genes in tiger puffer (Takifugu rubripes) across tissues and under different nutritional conditions. Fish. Physiol. Biochem. 2021, 47, 1739–1758. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, J.; Cai, X.N.; Ai, Y.; Long, H.; Ren, W.; Huang, A.Y.; Zhang, X.; Xie, Z.Y. Dietary supplementation of astaxanthin is superior to its combination with Lactococcus lactis in improving the growth performance, antioxidant capacity, immunity and disease resistance of white shrimp (Litopenaeus vannamei). Aquac. Rep. 2022, 24, 101124. [Google Scholar] [CrossRef]
- Keng, C.L.; Yusoff, M.F.; Shariff, M.; Kamarudin, M.S.; Nagao, N. Dietary supplementation of astaxanthin enhances hemato-biochemistry and innate immunity of Asian seabass, Lates calcarifer (Bloch, 1790). Aquaculture 2019, 512, 734339. [Google Scholar] [CrossRef]
- Wang, W.L.; Ishikawam, M.; Koshio, S.; Yokoyama, S.; Dawood, M.A.; Zhang, Y.K. Effects of dietary astaxanthin supplementation on survival, growth and stress resistance in larval and post-larval kuruma shrimp, Marsupenaeus japonicus. Aquac. Res. 2018, 49, 2225–2232. [Google Scholar] [CrossRef]
- Song, X.L.; Wang, L.; Li, X.Q.; Chen, Z.Z.; Liang, G.Y.; Leng, X.J. Dietary astaxanthin improved the body pigmentation and antioxidant function, but not the growth of discus fish (Symphysodon spp.). Aquac. Res. 2017, 48, 1359–1367. [Google Scholar] [CrossRef]
- Xie, S.; Yin, P.; Tian, L.; Yu, Y.; Liu, Y.; Niu, J. Dietary supplementation of astaxanthin improved the growth performance, antioxidant ability and immune response of juvenile largemouth bass (Micropterus salmoides) fed high-fat diet. Mar. Drugs 2020, 18, 642. [Google Scholar] [CrossRef]
- Xie, J.J.; Chen, X.; Liu, Y.J.; Tian, L.X.; Xie, S.W.; Niu, J. Effects of dietary astaxanthin on growth performance, hepatic antioxidative activity, hsp70, and HIF-1α gene expression of juvenile golden pompano (Trachinotus ovatus). Isr. J. Aquac.-Bamidgeh 2017, 1430, 12. [Google Scholar] [CrossRef]
- Fang, H.H.; He, X.S.; Zeng, H.L.; Liu, Y.J.; Tian, L.X.; Niu, J. Replacement of astaxanthin with lutein in diets of juvenile Litopenaeus vannamei: Effects on growth performance, antioxidant capacity, and immune response. Front. Mar. Sci. 2021, 8, 803748. [Google Scholar] [CrossRef]
- Tageldein, A.M.; Mohamed, A.; Eman, M.A.; Ahmed, S.A.; Mahmoud, S.K.; Mohamed, A.E.; Zaki, Z.S. Growth performance, immune-related and antioxidant genes expression, and gut bacterial abundance of pacific white leg shrimp, Litopenaeus vannamei, dietary supplemented with natural astaxanthin. Front. Physiol. 2022, 13, 874172. [Google Scholar] [CrossRef] [PubMed]
- Batoul, A.; Keramat, A.A.; Hosein, O.; Mohamad, K.; Soleiman, M. Effects of lysophospholipid on rainbow trout (Oncorhynchus mykiss) growth, biochemical indices, nutrient digestibility and liver histomorphometry when fed fat powder diet. Aquac. Nutr. 2021, 27, 1779–1788. [Google Scholar] [CrossRef]
- Liao, Z.B.; Xu, H.G.; Wei, Y.L.; Zhang, Q.G.; Liang, M.Q. Dietary astaxanthin differentially affected the lipid accumulation in the liver and muscle of the marine teleost, tiger puffer Takifugu rubripes. Aquac. Res. 2018, 10, 3421–3433. [Google Scholar] [CrossRef]
- Smith, B.E.; Hardy, R.W.; Torrissen, O.J. Synthetic astaxanthin deposition in pan-size coho salmon (Oncorhynchus kisutch). Aquaculture 1992, 104, 105–119. [Google Scholar] [CrossRef]
- Supamattaya, K.; Kiriratnikom, S.; Boonyaratpalin, M.; Borowitzka, L. Effect of a dunaliella extract on growth performance, health condition, immune response and disease resistance in black tiger shrimp (Penaeus monodon). Aquaculture 2005, 248, 207–216. [Google Scholar] [CrossRef]
- Zhang, J.J.; Li, X.Q.; Leng, X.J.; Zhang, C.L.; Han, Z.Y.; Zhang, F.G. Effects of dietary astaxanthins on pigmentation of flesh and tissue antioxidation of rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2013, 21, 579–589. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, S.; Dai, J.Y.; Wang, C.C.; Chen, S.M.; Qian, Y.X.; Han, T. Effects of synthetic astaxanthin on the growth performance, pigmentation, antioxidant capacity, and immune response in black tiger prawn (Penaeus monodon). Aquac. Nutr. 2023, 2023, 6632067. [Google Scholar] [CrossRef]
- Huang, S.; Chen, Q.; Zhang, M.M.; Chen, S.M.; Dai, J.Y.; Qian, Y.X.; Gong, Y.Y.; Han, T. Synthetic astaxanthin has better effects than natural astaxanthins on growth performance, body color and n-3 PUFA deposition in black tiger prawn (Penaeus monodon). Aquac. Rep. 2023, 33, 101816. [Google Scholar] [CrossRef]
- Zhao, X.P.; Wang, G.P.; Liu, X.G.; Guo, D.L.; Chen, X.L.; Liu, S.; Li, G.F. Dietary supplementation of astaxanthin increased growth, colouration, the capacity of hypoxia and ammonia tolerance of Pacific white shrimp (Litopenaeus vannamei). Aquac. Rep. 2022, 23, 101093. [Google Scholar] [CrossRef]
- Lin, Y.J.; Chang, J.J.; Huang, H.T.; Lee, C.P.; Hu, Y.F.; Wu, M.L.; Huang, C.Y.; Nan, F.H. Improving red-colorperformance, immune response and resistance to Vibrio parahaemolyticus on white shrimp Penaeus vannamei by an engineered astaxanthin yeast. Sci. Rep. 2023, 13, 2248. [Google Scholar] [CrossRef]
- Fatemeh, J.; Naser, A.; Farzaneh, N.; Enric, G.; Mansour, T.M. Supplementing lysolecithin in corn-oil based diet enhanced growth and improved body biochemical composition in juvenile stellate sturgeon (Acipenser stellatus). Anim. Feed. Sci. Technol. 2024, 310, 115945. [Google Scholar] [CrossRef]
- Juntanapum, W.; Bunchasak, C.; Poeikhampha, T.; Rakangthong, C.; Poungpong, K. Effects of supplementation of lysophosphatidylcholine (LPC) to lying hens on production performance, fat digestibility, blood lipid profile, and gene expression related to nutrients transport in small intestine. J. Anim. Feed Sci. 2020, 29, 258–265. [Google Scholar] [CrossRef]
- Nutautaitė, M.; Racevičiūtė, S.A.; Andalibizadeh, L.; Šašytė, V.; Bliznikas, S.; Pockevičius, A.; Vilienė, V. Improving broiler chickens’ health by using lecithin and lysophosphatidylcholine emulsifiers: A comparative analysis of physiological indicators. Iran. J. Vet. Res. 2021, 22, 33–39. [Google Scholar] [CrossRef]
- Li, S.H.; Luo, X.; Liao, Z.B.; Liang, M.Q.; Xu, H.G.; Mai, K.S.; Zhang, Y.J. Effects of lysophosphatidylcholine on intestinal health of turbot fed high-lipid diets. Nutrients 2022, 14, 4398. [Google Scholar] [CrossRef]
- Imran, H.K.; Syama, J.D.; Kondusamy, A.; Purdhvi, E.M.; Rajabdeen, J.; Vanjiappan, R. Enhancing the dietary value of palm oil in the presence of lysolecithin in tiger shrimp, Penaeus monodon. Aquac. Int. 2018, 26, 509–522. [Google Scholar] [CrossRef]
- Li, H.T.; Tian, L.X.; Wang, Y.D.; Hu, Y.H. Effects of lysolecithin on growth performance, body composition and hematological indices of hybrid tilapia (Oreochromis aureus ♂×Oreochromis niloticus ♀). J. Dalian Fish. Univ. 2010, 25, 143–146. [Google Scholar] [CrossRef]
- Wang, M.; Wu, X.H.; Li, Z. Leptin accelerates lipid metabolism by increasing lipoprotein lipase and hepatic lipase expression in insulin-resistant liver cell model. J. Third Mil. Med. Univ. 2014, 36, 1059–1063. [Google Scholar] [CrossRef]
- Chen, J.Y.; Chen, J.C.; Wu, J.L. Molecular cloning and functional analysis of zebrafish high-density lipoprotein-binding protein Comp. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136B, 117–130. [Google Scholar] [CrossRef]
- Deng, J.; Mai, K.; Ai, Q.; Zhang, W.; Wang, X.; Tan, B.; Xu, W.; Liu, Z.; Ma, H. Interactive effects of dietary cholesterol and protein sources on growth performance and cholesterol metabolism of Japanese flounder (Paralichthys olivaceus). Aquac. Nutr. 2010, 16, 419–429. [Google Scholar] [CrossRef]
- Yun, B.; Mai, K.S.; Zhang, W.B.; Xu, W. Effects of dietary cholesterol on growth performance, feed intake and cholesterol metabolism in juvenile turbot (Scophthalmus maximus L.) fed high plant protein diets. Aquaculture 2011, 319, 105–110. [Google Scholar] [CrossRef]
- Mardones, P.; Quiñones, V.; Amigo, L.; Moreno, M.; Miquel, J.F.; Schwarz, M.; Miettinen, H.E.; Trigatti, B.; Krieger, M.; VanPatten, S.; et al. Hepatic cholesterol and bile acid metabolism and intestinal cholesterol absorption in scavenger receptor class B type I-deficient mice. J. Lipid Res. 2001, 42, 170–180. [Google Scholar] [CrossRef]
- Stanley, S.L.; Stephen, G.W. Implications of reverse cholesterol transport: Recent studies. Clin. Chim. Acta 2015, 439, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Brufau, G.; Groen, A.K.; Kuipers, F. Reverse cholesterol transport revisited: Contribution of biliary versus intestinal cholesterol excretion. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1726–1733. [Google Scholar] [CrossRef]
- Wang, L.M.; Motamed, M.; Infante, E.R.; Abi-Mosleh, L.; Kwon, H.J.; Brown, M.S.; Goldstein, J.L. Identification of Surface Residues on Niemann-Pick C2 Essential for Hydrophobic Handoff of Cholesterol to NPC1 in Lysosomes. Cell Metab. 2010, 12, 166–173. [Google Scholar] [CrossRef]
- Wei, F.; Chen, Q.C.; Cui, K.; Chen, Q.; Li, X.S.; Xu, N.; Mai, K.S.; Ai, Q.H. Lipid overload impairs hepatic VLDL secretion via oxidative stress-mediated PKCδ-HNF4α-MTP pathway in large yellow croaker (Larimichthys crocea). Free Radic. Biol. Med. 2021, 20, 213–225. [Google Scholar] [CrossRef]
- Zhang, D.G.; Zhao, T.; Hogstrand, C.; Ye, H.M.; Xu, X.J.; Luo, Z. Oxidized fish oils increased lipid deposition via oxidative stress-mediated mitochondrial dysfunction and the CREB1-Bcl2-Beclin1 pathway in the liver tissues and hepatocytes of yellow catfish. Food Chem. 2021, 360, 129814. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, F.; Forchino, A.; Antonini, M.; Rimoldi, S.; Terova, G.; Saroglia, M. Effect of dietary astaxanthin sources supplementation on muscle pigmentation and lipid peroxidation in rainbow trout (Oncorhynchus mykiss). Ital. J. Anim. Sci. 2009, 8 (Suppl. S2), 845–847. [Google Scholar] [CrossRef]
- Xu, W.X.; Liu, Y.T.; Huang, W.X.; Yao, C.W.; Yin, Z.Y.; Mai, K.S.; Ai, Q.H. Effects of dietary supplementation of astaxanthin (Ast) on growth performance, activities of digestive enzymes, antioxidant capacity and lipid metabolism of large yellow croaker (Larimichthys crocea) larvae. Aquac. Res. 2022, 53, 4605–4615. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Control | AST | AST + LPC |
|---|---|---|---|
| Fish meal | 20 | 20 | 20 |
| Soybean meal | 30 | 30 | 30 |
| Peanut meal | 14 | 14 | 14 |
| Poultry by-product meal | 4 | 4 | 4 |
| Wheat meal | 21 | 21 | 21 |
| Mineral premix a | 0.5 | 0.5 | 0.5 |
| Vitamin premix a | 1 | 1 | 1 |
| Monocalcium phosphate | 1 | 1 | 1 |
| Vitamin C | 0.2 | 0.2 | 0.2 |
| Choline chloride | 0.2 | 0.2 | 0.2 |
| Ethoxyquin | 0.02 | 0.02 | 0.02 |
| Mold inhibitor | 0.1 | 0.1 | 0.1 |
| Betaine | 0.3 | 0.3 | 0.3 |
| Soya lecithin | 1.5 | 1.5 | 1.5 |
| Soybean oil | 3 | 3 | 3 |
| CarophyllPink b | 0 | 0.2 | 0.2 |
| Lysophosphatidylcholine c | 0 | 0 | 0.1 |
| Y2O3 | 0.1 | 0.1 | 0.1 |
| Bentonite | 1.08 | 0.88 | 0.78 |
| Alginate | 2 | 2 | 2 |
| Proximate composition | |||
| Crude protein | 43.9 | 44.0 | 44.5 |
| Crude lipid | 6.5 | 6.4 | 6.7 |
| Ash | 10.3 | 10.5 | 10.2 |
| Moisture | 8.3 | 8.1 | 8.5 |
| Gross energy (MJ/kg) | 17.8 | 17.8 | 17.7 |
| Primer | Forward Primer (5′−3′) | GenBank Reference | Product Length (bp) |
|---|---|---|---|
| npc1-F | CGAAGGGGAAAAGCCAGAGT | XM_027363410.1 | 87 |
| npc1-R | TTGAGGAGGAAGGGAGCGTA | ||
| npc2-F | CGCAGTATCGGCAGTCAAGA | XM_027358057.1 | 149 |
| npc2-R | GTGGTGTGAAAGGCAACGAC | ||
| abca1-F | ACCTGAAGGCAGGACGAAAG | XM_027375794.1 | 73 |
| abca1-R | GGCATACTCGCGCTATCTGT | ||
| abcg1-F | TCCCGGAAGGCAGGAATAGA | XM_027356334.1 | 121 |
| abcg1-R | TTCATCAGCGTCGACTTCCC | ||
| abcg5-F | TCCGTTTGCCCCGATAACAA | XM_027382028.1 | 74 |
| abcg5-R | TAGCGCTCGAGCAGGTAGTA | ||
| abcg8-F | CCAACGATTCCGAAGGGTCT | XM_027368984.1 | 93 |
| abcg8-R | CGTTGAGGATGAAGTCCCCC | ||
| ldlr-F | CGTCACATGCCCAGCCATAA | XM_027363319.1 | 86 |
| ldlr-R | GATCCGTCATGGCACTCGAA | ||
| srb1-F | GTTCGACATCTACCCGGACC | XM_027352601.1 | 132 |
| srb1-R | AACCAGAAGATGGGCAGGAC | ||
| mttp-F | GCTGCTAAGGAAAGTGCGTG | XM_027380336.1 | 215 |
| mttp-R | AAGGATGCGTCGCTAAGGAG | ||
| apod-F | CAACGCGGTAACAGGGAAAG | XM_027368898.1 | 85 |
| apod-R | GACAACCAGCTTGGCTTCAC | ||
| ef-1α-F | GTATTGGAACAGTGCCCGTG | GU136229.1 | 143 |
| ef-1α-R | ACCAGGGACAGCCTCAGTAAG | ||
| β-actin-F | CGAGGTATCCTCACCCTGAA | AF300705.2 | 176 |
| β-actin-R | GTCATCTTCTCGCGGTTAGC |
| Parameter | Control | AST | AST + LPC | p Value |
|---|---|---|---|---|
| Initial weight g | 2.41 ± 0.09 | 2.34 ± 0.04 | 2.58 ± 0.05 | 0.162 |
| Final weight g | 10.35 ± 0.03 a | 10.96 ± 0.25 ab | 11.45 ± 0.34 b | 0.046 |
| Weight gain g | 7.94 ± 0.07 a | 8.62 ± 0.21 ab | 8.87 ± 0.29 b | 0.041 |
| Weight gain % | 330.33 ± 14.31 | 367.67 ± 3.28 | 344.00 ± 5.00 | 0.229 |
| Feed conversion ratio | 1.35 ± 0.06 | 1.24 ± 0.05 | 1.24 ± 0.04 | 0.573 |
| Feed intake % | 2.93 ± 0.14 | 2.83 ± 0.10 | 2.82 ± 0.01 | 0.792 |
| Survival % | 95.33 ± 2.33 | 98.00 ± 1.00 | 98.50 ± 1.50 | 0.460 |
| Parameter | Control | AST | AST + LPC | p Value |
|---|---|---|---|---|
| Whole shrimp | ||||
| Moisture | 77.20 ± 0.61 | 76.71 ± 0.68 | 76.76 ± 1.03 | 0.894 |
| Crude protein | 17.59 ± 0.47 | 17.72 ± 0.42 | 17.83 ± 0.58 | 0.943 |
| Crude lipid | 0.68 ± 0.07 | 1.32 ± 0.26 | 1.38 ± 0.38 | 0.583 |
| Ash | 3.10 ± 0.12 | 2.90 ± 0.10 | 2.90 ± 0.20 | 0.582 |
| Muscle | ||||
| Moisture | 74.74 ± 0.40 | 74.36 ± 1.11 | 74.33 ± 0.32 | 0.902 |
| Crude protein | 22.65 ± 0.29 | 22.80 ± 82 | 22.58 ± 0.33 | 0.959 |
| Crude lipid | 1.15 ± 0.08 | 1.13 ± 0.07 | 1.17 ± 0.02 | 0.873 |
| Fatty Acid | Control | AST | LPC + AST | p Value |
|---|---|---|---|---|
| 14:0 | 0.33 ± 0.04 | 0.21 ± 0.01 | 0.33 ± 0.04 | 0.093 |
| 16:0 | 19.54 ± 0.29 | 19.49 ± 0.18 | 20.07 ± 1.00 | 0.764 |
| 18:0 | 9.21 ± 0.09 | 9.96 ± 0.24 | 9.32 ± 0.26 | 0.098 |
| 20:0 | 0.19 ± 0.00 | 0.19 ± 0.00 | 0.19 ± 0.01 | 0.527 |
| SFA | 29.26 ± 0.28 | 29.85 ± 0.06 | 29.91 ± 0.86 | 0.648 |
| 14:1n-5 | 0.20 ± 0.02 b | 0.14 ± 0.01 a | 0.19 ± 0.02 ab | 0.035 |
| 16:1n-7 | 1.10 ± 0.08 b | 0.76 ± 0.04 a | 1.07 ± 0.06 b | 0.013 |
| 18:1n-9 | 13.44 ± 0.13 ab | 12.83 ± 0.15 a | 13.90 ± 0.21 b | 0.011 |
| 22:1n-9 | 0.16 ± 0.00 | 0.17 ± 0.01 | 0.18 ± 0.01 | 0.530 |
| MUFA | 14.90 ± 0.11 b | 13.90 ± 0.19 a | 15.35 ± 0.25 b | 0.005 |
| 8:2n-6 | 18.58 ± 0.32 a | 18.88 ± 0.42 ab | 19.99 ± 0.04 b | 0.038 |
| 20:2n-6 | 1.54 ± 0.03 | 1.72 ± 0.08 | 1.59 ± 0.08 | 0.242 |
| n-6 PUFA | 21.82 ± 0.31 a | 22.45 ± 0.41 ab | 23.20 ± 0.17 b | 0.055 |
| 18:3n-3 | 1.05 ± 0.03 | 0.99 ± 0.06 | 1.10 ± 0.04 | 0.292 |
| 20:3n-3 | 1.69 ± 0.03 | 1.84 ± 0.08 | 1.62 ± 0.13 | 0.295 |
| 20:5n-3 | 9.39 ± 0.15 | 9.75 ± 0.22 | 8.61 ± 0.37 | 0.057 |
| 22:5n-3 | 0.84 ± 0.05 | 0.77 ± 0.04 | 0.83 ± 0.03 | 0.456 |
| 22:6n-3 | 10.41 ± 0.19 | 10.14 ± 0.24 | 9.58 ± 0.23 | 0.092 |
| n-3 PUFA | 23.38 ± 0.07 ab | 23.48 ± 0.16 b | 21.73 ± 0.64 a | 0.031 |
| n-3/n-6 | 0.99 ± 0.01 b | 0.97 ± 0.02 b | 0.87 ± 0.02 a | 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Z.; Liu, Y.; Liu, H.; Ye, Z.; Ma, Q.; Wei, Y.; Xiao, L.; Liang, M.; Xu, H. Dietary Lysophosphatidylcholine Improves the Uptake of Astaxanthin and Modulates Cholesterol Transport in Pacific White Shrimp Litopenaeus vannamei. Antioxidants 2024, 13, 505. https://doi.org/10.3390/antiox13050505
Song Z, Liu Y, Liu H, Ye Z, Ma Q, Wei Y, Xiao L, Liang M, Xu H. Dietary Lysophosphatidylcholine Improves the Uptake of Astaxanthin and Modulates Cholesterol Transport in Pacific White Shrimp Litopenaeus vannamei. Antioxidants. 2024; 13(5):505. https://doi.org/10.3390/antiox13050505
Chicago/Turabian StyleSong, Ziling, Yang Liu, Huan Liu, Zhengwei Ye, Qiang Ma, Yuliang Wei, Lindong Xiao, Mengqing Liang, and Houguo Xu. 2024. "Dietary Lysophosphatidylcholine Improves the Uptake of Astaxanthin and Modulates Cholesterol Transport in Pacific White Shrimp Litopenaeus vannamei" Antioxidants 13, no. 5: 505. https://doi.org/10.3390/antiox13050505
APA StyleSong, Z., Liu, Y., Liu, H., Ye, Z., Ma, Q., Wei, Y., Xiao, L., Liang, M., & Xu, H. (2024). Dietary Lysophosphatidylcholine Improves the Uptake of Astaxanthin and Modulates Cholesterol Transport in Pacific White Shrimp Litopenaeus vannamei. Antioxidants, 13(5), 505. https://doi.org/10.3390/antiox13050505