
Immunity from NK Cell Subsets Is Important for Vaccine-Mediated Protection in HPV+ Cancers

1
Department of Thoracic Head and Neck Medical Oncology, Division of Cancer Medicine, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA
2
UTHealth Graduate School of Biomedical Sciences, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA
*
Author to whom correspondence should be addressed.
Vaccines 2024, 12(2), 206; https://doi.org/10.3390/vaccines12020206
Submission received: 26 December 2023
/
Revised: 7 February 2024
/
Accepted: 14 February 2024
/
Published: 17 February 2024
{kind=link}
{kind=link}
{kind=link}
Abstract
:High-risk human papillomaviruses (HPVs) are associated with genital and oral cancers, and the incidence of HPV+ head and neck squamous cell cancers is fast increasing in the USA and worldwide. Survival rates for patients with locally advanced disease are poor after standard-of-care chemoradiation treatment. Identifying the antitumor host immune mediators important for treatment response and designing strategies to promote them are essential. We reported earlier that in a syngeneic immunocompetent preclinical HPV tumor mouse model, intranasal immunization with an HPV peptide therapeutic vaccine containing the combination of aGalCer and CpG-ODN adjuvants (TVAC) promoted clearance of HPV vaginal tumors via induction of a strong cytotoxic T cell response. However, TVAC was insufficient in the clearance of HPV oral tumors. To overcome this deficiency, we tested substituting aGalCer with a clinically relevant adjuvant QS21 (TVQC) and observed sustained, complete regression of over 70% of oral and 80% of vaginal HPV tumors. The TVQC-mediated protection in the oral tumor model correlated with not only strong total and HPV-antigen-specific CD8 T cells, but also natural killer dendritic cells (NKDCs), a novel subset of NK cells expressing the DC marker CD11c. Notably, we observed induction of significantly higher overall innate NK effector responses by TVQC relative to TVAC. Furthermore, in mice treated with TVQC, the frequencies of total and functional CD11c+ NK cell populations were significantly higher than the CD11c− subset, highlighting the importance of the contributions of NKDCs to the vaccine response. These results emphasize the importance of NK-mediated innate immune effector responses in total antitumor immunity to treat HPV+ cancers.
1. Introduction
High-risk human papillomaviruses (HPVs) are the underlying cause for over 5% of all cancers. Despite its decreasing incidence in developed countries, cervical cancer remains the second most common cause of death from cancer in women worldwide [1,2]. In addition, there has been an alarming increase in the incidence of HPV-associated oropharyngeal cancers (OPCs) in recent decades, relative to that of HPV+ cervical cancer, and now considered an epidemic [3,4]. Among fifteen different high-risk HPV genotypes, HPV-16 accounts for over 70% of cervical and 95% of HPV+ OPCs [1,4].
The viral-encoded oncoproteins E6 and E7, critical for the initiation and maintenance of HPV-associated malignancies, are ideal targets for therapeutic HPV vaccines [5,6]. However, vaccines targeting E6 and E7 have thus far shown modest efficacy in clinical trials despite inducing strong antigen-specific T cell responses [7]. The innate immunity forms the frontline defense against infections and diseases, including HPV-associated malignancies [8,9,10,11,12]. Therefore, vaccination strategies that significantly enhance antitumor immune responses with multiple effector functions to overcome the prevailing immunosuppressive tumor microenvironment are necessary to achieve maximum efficacy.
Adjuvants are an integral part of vaccine formulations that enhance the immunogenicity of vaccine antigens and activate innate immune mediators as well as effectors that bridge the innate and adaptive immune responses, specifically the antigen-presenting cells or dendritic cells [8,13]. The glycolipid α-galactosylceramide (aGalCer) is a potent agonist to natural killer T cells (NKTs), which are among the most effective innate immune modulators, for inducing the activation and maturation of dendritic cells (DCs) that, in turn, induce adaptive immune responses [14,15]. Toll-like receptor (TLR) ligand-based adjuvants are among the most commonly tested reagents for improving vaccine potency. Synthetic oligodeoxynucleotides containing one or more unmethylated CpG dinucleotides (CpG-ODN), which are common in bacterial and viral genomic DNA but not in vertebrates, serve as potent stimulators of TLR9 [16]. Unlike the TLR ligand-based adjuvants, QS21 is one of the best characterized natural saponin adjuvants [17,18].
We have previously reported that the HPV-16 E6 and E7 peptides are stimulators of T cell immune memory in HPV+ patients [19]; when formulated as a therapeutic vaccine with aGalCer and CpG-ODN adjuvants (TVAC) and delivered by the intranasal route, it induces strong mucosal and systemic antigen-specific T cell responses and protection against vaginal HPV tumors in C57BL/6 mice [20,21]. Here, we investigated the effectiveness of TVAC against an oral HPV tumor model and present evidence demonstrating the efficacy of a modified vaccine formulation where the aGalCer adjuvant in TVAC is replaced by another adjuvant QS21. The resulting vaccine, termed TVQC (therapeutic vaccine containing QS21 and CpG-ODN adjuvants), afforded protection in both the oral and vaginal preclinical HPV tumor models. We found that the curative efficacy of TVQC, in terms of >70% of mice exhibiting sustained long-term tumor regression, was associated with robust induction of multiple polyfunctional antitumor effector populations, in addition to HPV-antigen-specific CD8 T cells, including a subset of innate NK cells that express the dendritic cell marker CD11c, referred to as NKDCs.
2. Materials and Methods
2.1. Animals
C57BL/6J male and female mice (6–8 weeks) were procured from The Jackson Laboratory (Bar Harbor, ME, USA) and maintained in a pathogen-free environment. All animal studies were pre-approved and carried out in accordance with the University of Texas MD Anderson Cancer Center Institutional Animal Care and Use Committee (IACUC) guidelines. Mice were monitored daily for overall health and euthanized according to IACUC criteria.
2.2. Tumor Cell Lines
The mEER tumor cells are mouse tonsil epithelial cells expressing HPV-16 E6 and E7 along with H-ras, and they were a kind gift from Dr. John Lee (Avera Cancer Institute, Sioux Falls, SD, USA). These cells were maintained in complete Dulbecco’s Modified Eagle Medium (DMEM)/F12 media containing various supplements as previously described [22]; cells were sub-cultured at 80% confluence the day before tumor induction in mice. The TC-1-luc tumor cell line is of lung epithelial origin from C57BL/6 mice, expressing the E6 and E7 oncogenes of HPV-16 as well as firefly luciferase and H-ras. This cell line was a kind gift from Drs. T.-C. Wu and C. Hung (Johns Hopkins School of Medicine, Baltimore, MD, USA). The TC-1luc cells were maintained in complete Roswell Park Memorial Institute (RPMI) 1640 media supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Atlanta Biologicals, Flowery Branch, GA, USA), 50 units/mL of penicillin–streptomycin and 50 μg/mL gentamycin.
2.3. In Vivo Tumor Challenge
Syngeneic male C57BL/6 mice were implanted with 4 × 104 mEER cells in 50 μL phosphate-buffered saline (PBS) into the base of the tongue as described previously [23,24]. Mice were monitored twice weekly for body weight changes. Mice with oral tumors were euthanized when the mice lost 20% or more of their initial body weight. For the experiments involving the analysis of tumor-infiltrating leukocytes (TILs), 1 × 105 mEER tumor cells in PBS were mixed in a 2:1 ratio with Matrigel (BD Biosciences, San Jose, CA, USA) for implantation. For intravaginal tumor challenge, 2 × 104 TC-1 cells were implanted in the vaginal tract of diestrus-synchronized 6- to 8-week-old female C57BL/6 mice according to a previously described protocol [25,26]. Intravaginal TC-1 tumor growth was monitored using an In Vivo Imaging System (IVIS), a small animal imaging system (Perkin Elmer, Waltham, MA, USA), and expressed as the average luminescent signal in a selected region of interest (ROI) (p/s/cm2/sr) as a measure of tumor size. For the TIL experiments, 3 × 104 TC-1 cells mixed 2:1 with Matrigel were used for intravaginal implantation.
2.4. Intranasal Vaccination
The therapeutic vaccine consisted of the following four peptides corresponding to the HPV-16 E6 and E7 oncoproteins: the E643–57 peptide, Q15L (QLLRREVYDFAFRDL); the E649–58 peptide, V10C (VYDFAFRDLC); the E744–62 peptide, Q19D (QAEPDRAHVYNIVTFCCKCD); and the E749–57 peptide, R9F (RAHVYNIVTF) [19,20]. Vaccine-grade (>90% pure) peptides were purchased from Elim Biopharma (Hayward, CA, USA) and used at 100 μg per dose of each, along with single or combinations of adjuvants (10 μg CpG-ODN, 5 μg QS-21, or 2 μg aGalCer). The R9F and V10C peptides represent murine H2b-restricted epitopes. Vaccine-grade ODN 1826 was purchased from InvivoGen (San Diego, CA, USA), and GMP-grade QS-21 was from Desert King (San Diego, CA, USA). ODN 1826 is a type B CpG-ODN specific for mouse TLR9 that is known to induce a Th1 response in addition to B cell activation, and it is a weak stimulator of IFNα and plasmacytoid DC [27,28]. The adjuvant aGalCer was purchased from DiagnoCline (Hackensack, NJ, USA). All adjuvants were reconstituted according to the manufacturers’ recommendations. Tumor-bearing mice were randomized, and on days 5 and 11 following tumor implantation, vaccines were administered via the intranasal route as described previously [23,26]. Untreated mice that received intranasal endotoxin-free PBS or those treated with a mixture of peptides and individual adjuvants served as control groups.
2.5. Magnetic Resonance Imaging (MRI)
For the mEER oral tumor model, tumor volume was measured using MRI. Mice were imaged under 2% isoflurane vapor anesthesia in a 7T small animal MR scanner (Biospec, Bruker Biospin Inc., Billerica, MA, USA) using transmit/receive volume coils with an inner diameter of 35 mm. The following parameters and settings were used for acquiring image sequences: field-of-view 4 × 3 cm2, matrix 256 × 192 and spatial resolution 156 microns; a T2-weighted coronal Rapid Acquisition with Relaxation Enhancement (RARE) with T2-Sagital; echo time 38 ms; repetition 1800 ms; slice thickness 0.50 m; and slice gap 0.50 mm. Tumor volume was calculated based on the image sequences analysis in three dimensions using Image J software version 1.53 (NIH, Bethesda, MD, USA) after defining the region of interest (tumor) on all possible sections.
2.6. Immune Cell Isolation
On day 15 or 16 post-tumor challenge, mice were euthanized and tumors, spleens, and tumor-draining lymph nodes (LNs) were collected to characterize the cell-mediated antitumor immune responses. Briefly, tumors were digested in complete RPMI media containing Collagenase H (Sigma-Aldrich, St. Louis, MO, USA) and DNase (Roche, Indianapolis, IN, USA), and they were incubated at 37 °C for 45 min before passing through a 70 μm cell strainer. Tumor-infiltrating leukocytes (TILs) were isolated using 67%:44% Percoll (Cytiva, Marlborough, MA, USA) gradient centrifugation. Single-cell suspensions from the spleens were prepared using mechanical disruption and passing through cell strainers followed by red blood cell lysis. In mice with mEER oral tumors, the cervical lymph nodes were collected, and in mice with TC-1 vaginal tumors, the inguinal lymph nodes were collected. In both cases, these tumor-draining lymph nodes were processed using mechanical disruption through a 70 μm cell strainer followed by centrifugation and re-suspension in complete RPMI medium.
2.7. Flow Cytometry
Following isolation, cells were incubated at 37 °C in 5% CO2 for 4–6 h with GolgiPlug/Brefeldin A (ThermoFisher, Waltham, MA, USA). Cells were blocked using mouse Fc-block (anti-CD16/32), stained for surface markers, and fixed and permeabilized using the intracellular/Foxp3 Fix-Perm reagent kit from eBioscience (ThermoFisher, Waltham, MA, USA); this was followed by staining for intracellular/functional markers (Table S1). FACS data acquisition was performed on a five-laser Fortessa X-20 flow cytometer (BD Bioscience) and analyzed using FlowJo version 10 (FlowJo LLC, Ashland, OR, USA). Forward and side scatter parameters were used to set the singlet and leukocyte gates. The fixable viability stain 510 included in the surface antibody cocktail was used to gate-out dead cells and only analyze viable cells. HPV-16 E7 antigen-specific CD8 T cells were detected using a specific tetramer reagent that was part of the multiparametric panel. The APC-labeled H-2Db epitope E749–57 (RAHYNIVTF)-peptide-loaded tetramer was obtained from the NIH MHC tetramer core facility at Emory University (Atlanta, GA, USA). For the detection of NKDCs, the gating strategy involving CD3-NK1.1+ CD11b+(int) followed by expression of CD11c+ (Figure S1) was followed, as previously described by Terme et al. [29].
2.8. Statistical Analysis
All graphing and statistical analyses were performed using GraphPad Prism version 8 (San Diego, CA, USA). An ordinary one-way ANOVA was used for comparing multiple groups, and a two-sample Student’s t-test was applied for comparing two groups. Statistical significance for matched pairs was determined using the Wilcoxon matched pairs test. Statistical significance between the survival curves was calculated using a log-rank test (Mantel–Cox). Graphs show the mean ± standard deviation (SD) for each group unless otherwise indicated. Experiments were repeated at least twice to confirm the reproducibility of results.
3. Results
3.1. Association of Innate Immunity from NKDCs with Vaccine-Mediated Protection in the Oral HPV Tumor Mouse Model
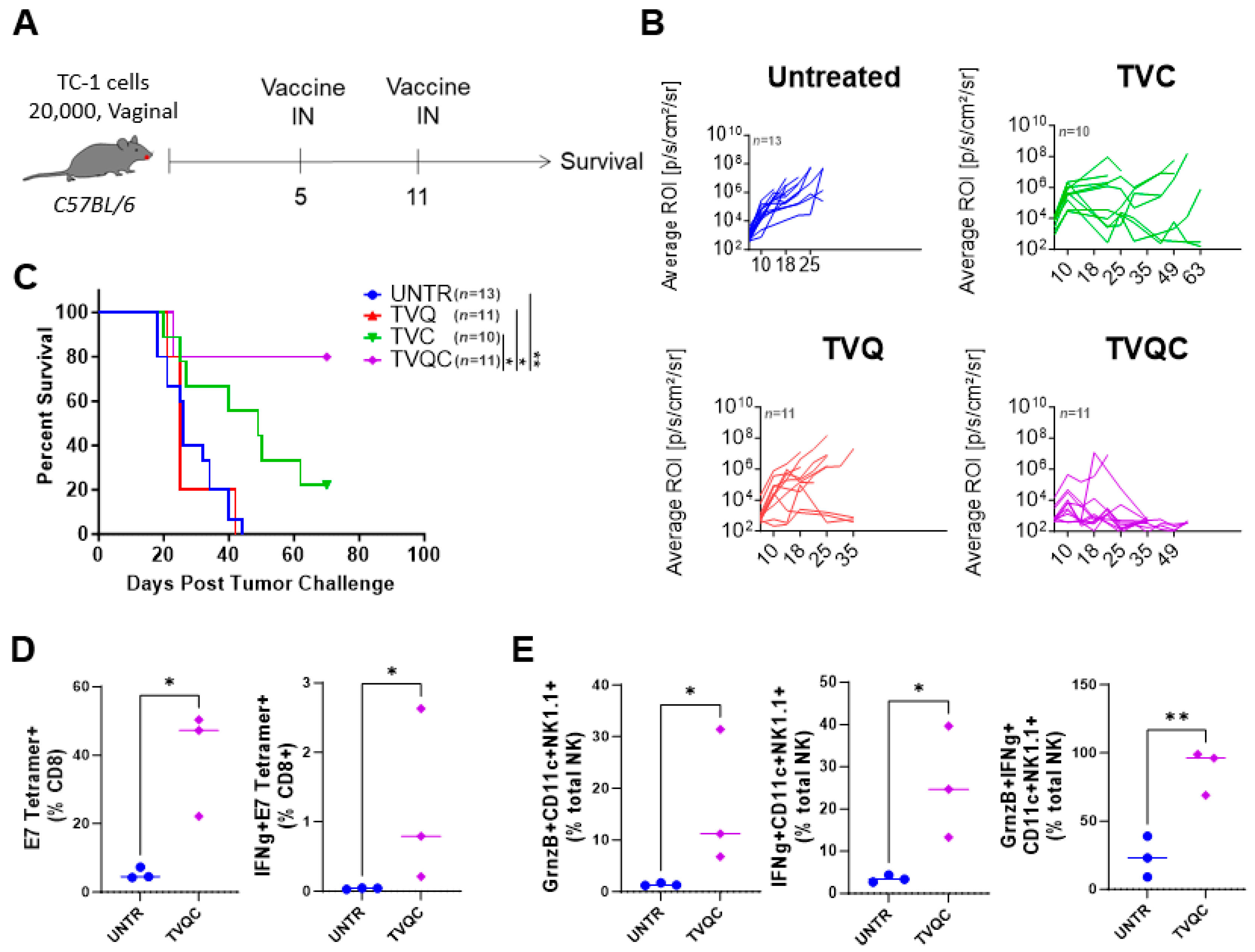
Based on the previously reported efficacy of the HPV-16 E6/E7 peptide therapeutic vaccine adjuvanted with aGalCer and CpG (TVAC) in the TC-1-Luc vaginal HPV tumor model [21], we tested intranasal administration of TVAC to mEER oral HPV tumor-bearing mice as described before [23,24]. Mice with visible tongue tumors by day 5 were randomized and either untreated or treated with TVAC or the therapeutic vaccine containing single adjuvants aGalCer (TVA) or CpG-ODN (TVC) by the intranasal route on days 5 and 11 after tumor implantation (Figure 1A). We observed that mice treated with TVAC and TVC as well as untreated groups experienced a high tumor burden and were sacrificed on day 51 (Figure 1B). Thus, in contrast to the significant TVAC-mediated tumor-free survival we reported in mice with vaginal HPV tumors [21], the present data indicate poor efficacy of TVAC against oral HPV tumors.
Because TVAC provided ineffective protection in the oral tumor model, we tested a modified therapeutic vaccine to incorporate the combination of QS21 and CpG-ODN (TVQC), where QS21 was used in place of aGalCer for intranasal administration in the mEER oral model. We observed that 72% of mice treated with TVQC showed sustained tumor regression and extended survival for 10 weeks following tumor challenge; this was significantly higher when compared to the control groups, including the untreated mice (Figure 1C). In addition, the tumor volume based on MRI measurements of the oral tumors on day 18 showed a significant reduction in tumor size in the TVQC-treated mice compared to the untreated mice (Figure 1D). Analysis of the tumor-infiltrating leukocytes (TILs) indicated that both TVAC and TVQC were effective in inducing strong HPV E7 antigen-specific CD8 T cell responses, even though the levels were higher in the mice treated with TVQC (Figure 1E). Thus, these data, while clearly demonstrating the enhanced effectiveness of TVQC relative to TVAC to protect against oral HPV tumors, the immunological responses beyond the adoptive HPV-specific CD8 T cells are unclear.
We therefore carried out multiparametric flow cytometric analyses of TILs from mice treated with the two vaccines in comparison to those in untreated mice, focusing on the levels of the innate immune NK cell response (Figure 2). We observed significantly elevated frequencies of total and functional (granzyme B- and/or IFNg-expressing) NK cells in mice treated with TVQC (Figure 2A). Importantly, a subset of total and functional NK cells that express the DC marker CD11c, referred to in the literature as NKDCs, were significantly higher in mice treated with TVQC compared to TVAC-treated mice (Figure 2B). To evaluate the contribution of NKDCs to the efficacy of TVQC, we compared the CD11c+ NK cells (NKDCs) to the CD11c− subset within the tumor microenvironment (TME) of TVQC-vaccinated mice. We observed significantly elevated frequencies of total as well as granzyme B- and/or IFNg-expressing NKDCs compared to the CD11c− subset (Figure 2C). Together, these data highlight the importance of innate effector immunity, specifically NKDCs, in the therapeutic efficacy of TVQC. The frequency of CD4 and CD8 T cells was also analyzed (Figure S2).
3.2. Both HPV E7-Specific and NKDC Responses Are Associated with TVQC-Mediated Protection against HPV Vaginal Tumors
We further evaluated whether TVQC, effective against oral HPV tumors in the mEER model, would also be useful to treat vaginal HPV tumors using the TC-1 model as an immunocompetent preclinical surrogate for HPV+ genital cancers [26,30]. Syngeneic C57BL/6 female mice were implanted with intravaginal TC-1luc tumor cells, as described in the Methods and Materials, and treated with TVQC delivered by the intranasal route on days 5 and 11 post-tumor implantation (Figure 3A). We observed tumor regression (Figure 3B) along with significant survival advantages in 80% of the vaccinated mice (Figure 3C). The therapeutic efficacy of TVQC correlated with significant induction of total and IFNg-producing tumor-antigen-specific CD8 T cells (Figure 3D) and NKDCs expressing granzyme B, IFNg or both (Figure 3E) in the tumor when compared to the untreated controls.
4. Discussion
Data presented here demonstrate that the adaptive immunity from total and HPV-antigen-specific CD8 T cells as well as the innate immunity from NK cell subsets both contribute towards the therapeutic vaccine-mediated protection against HPV oral tumors in the established preclinical model for HPV+ oropharyngeal cancers. Importantly, the vaccine containing the combination of QS21 and CpG-ODN as adjuvants (TVQC) was effective in inducing NKDCs, a novel subset of NK cells that express the dendritic cell marker CD11c, while the vaccine formulation with aGalCer in place of QS21 (TVAC) was less efficient in inducing NKDCs despite priming strong adaptive immunity via significant frequencies of antigen-specific CD8 T cells by both vaccines. Since robust induction of HPV-antigen-specific CTLs by TVAC was sufficient to treat TC-1 vaginal tumors [21], it appears that protection in the orthotopic mEER oral tumors requires induction of both an HPV-antigen-specific CTL response and a functional innate NK cell response in the form of functional NKDCs.
The NKDCs have been reported to possess both NK and antigen-presenting cell (APC) functions, potentially capable of cross-presenting tumor antigens [29], thus acting as a key link between the innate and adaptive immune responses. Even though a formal analyses for the APC function of NKDCs was not conducted in the present investigation, we believe that NKDCs contributed to the enhanced adaptive antigen-specific CTL response, as indicated by the relatively high frequency of HPV E7-specific CD8 T cells in the TME of mice treated with TVQC vs. those of TVAC (Figure 1E).
Innate, antigen-independent immune responses are key components of the tumor microenvironment, and their importance in promoting better cancer therapeutic outcomes is well known [9,10,31]. Moreover, in virus-associated cancers, such as the HPV+ cervical and oropharyngeal tumors, NK cells are known to be crucial in immunosurveillance and antitumor immunity [12]. In fact, the presence of activated NK cells in head and neck squamous cell carcinoma (HNSCC) has been shown to correlate with significantly better clinical outcomes [11,32]. However, we believe our study is the first report to show the role of NK responses in determining the efficacy of therapeutic vaccines against HPV-associated cancers, as well as the adequate induction of viral antigen-specific CD8 cytotoxic T cells (CTLs). The important attribute of TVQC efficacy in the current investigation is the robust induction of multiple effector immune responses, including polyfunctional NKDCs and tumor antigen-specific CTLs.
It is important to note that while total NK cells provide important contributions to the innate effector response of TVQC, in our studies, we observed that the CD11c+ NK subset (NKDC) exhibited a more potent functional phenotype than the CD11c− subset (Figure 2C), highlighting the significance of NKDCs in antitumor immunity. Similar to our findings, others have reported increased IFNg production by NKDCs compared to total NK cells [33,34]. Here, in addition to elevated IFNg, we show enhanced cytotoxicity shown by granzyme B and higher polyfunctionality (GrnzB+, IFNg+) from TVQC, all of which are known to be key components of the antitumor response [34,35].
The NK subset, NKDCs, has been reported as a potent innate immune responder with lytic capabilities and serves as an important source of IFNg production when activated [33,36]. However, a limitation of NKDCs in the immune response is their low frequency in circulation [33]. The scarcity of NKDCs highlights the importance of identifying a means to expand and activate them. It is known that adjuvants, as essential components of vaccine formulations, play an important role in mediating the production of cytokines, which in turn promotes various immune effector populations. Published reports show that aGalCer induces IL-4 and IFNg; CpG-ODN induces type I IFN; and QS21 induces IL-1, IL-18, and IL-12 [37,38,39,40,41,42]. The precise mechanisms underlying the effectiveness of the QS21 and CpG-ODN adjuvant combination to promote significant innate and adaptive cellular immune responses observed in the present investigation are unclear. Semmling et al. [43,44] described the phenomenon termed the dual licensing of DCs. They used a combination of adjuvants that use different signaling mechanisms to induce the secretion of distinct sets of chemokines to recruit multiple sets of effector immune cells. It is possible that the balanced Th1 and Th2 responses from QS21 [18], when combined with TLR9-mediated signaling from the CpG-ODN in the TVQC vaccines, promoted the effective induction of multiple effector immune responses. It is known that IFNg is important in antitumor immunity by promoting NK and CD8 CTL functions, while IL-15 modulates antitumor immunity by promoting NK cells and CD8 T cell memory [45,46,47,48]. Moreover, IL-15 by itself has been used as a mucosal adjuvant along with the BCG vaccine and autologous whole-cell lung tumor vaccines in preclinical models [49,50,51]. Chaudry et al. reported that expansion with IL-15 significantly enhanced IFNg production and proliferation of NKDCs [33]. Furthermore, combinations of IL-2, IL-15, IL-18, and IL-21 have been used to successfully expand the total and subsets of NK cells [52,53], and we obtained preliminary data showing higher plasma levels of these cytokines in mice treated with TVQC . Overall, the elevated functionality and expansion potential of NKDCs, by cytokines or adjuvants, makes them an attractive subset for cellular immunotherapy to promote effective antitumor immunity.
5. Conclusions
Overall, evidence from the present investigation into preclinical oral and vaginal HPV tumor models supports and extends our earlier observations concerning the effectiveness of therapeutic HPV peptide vaccination by incorporating combinations of diverse adjuvants. While adaptive immunity, in terms of antigen-specific CD8 T cells, was essential for protection in both the preclinical models, the present investigation highlights the importance of the innate immunity afforded by NKDCs, a subset of NK cells, in vaccine-mediated protection against oral HPV tumors. Importantly, incorporation of QS21, a saponin adjuvant, along with CpG-ODN into the vaccine formulation (TVQC) was effective in inducing the NKDC responses. In TVQC-vaccinated mice, NKDCs in the TME exhibited high polyfunctionality, indicative of high levels of cytokine secretion and cytotoxicity. Importantly, we observed significant functional advantages in the CD11c+ (NKDC) vs. the CD11c− subset, highlighting the importance of this NK subset in the antitumor immunity mediated by the vaccine. Future studies testing the depletion and/or adoptive transfer of NKDCs should further substantiate their protective role. Additionally, since NKDCs express the DC marker CD11c, it would be interesting to investigate their potential contributions as antigen-presenting cells.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/vaccines12020206/s1. Figure S1: Flow cytometry gating strategy for NKDC in mice Figure S2: Total CD4 and CD8 T cells. Table S1: Antibodies used for flow cytometric analyses.
Author Contributions
Conceptualization, M.P.O. and K.J.S.; formal analysis, M.P.O. and K.J.S.; investigation, M.P.O., A.V.Y. and K.J.S.; writing—original draft preparation, M.P.O.; writing—review and editing, K.J.S.; visualization, M.P.O. and K.J.S.; supervision, K.J.S.; and funding acquisition, K.J.S. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the Cancer Prevention and Research Institute of Texas (CPRIT RP180472) and the HPV Moonshot grant awards from the MD Anderson Cancer Center.
Institutional Review Board Statement
All animal procedures were conducted in compliance with the University of Texas MD Anderson Cancer Center Institutional Animal Care and Use Committee (IACUC) guidelines (Project ID: 00000858-RN04, approval date 4 November 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The MD Anderson Cancer Center Advanced Cytometry and Sorting Core Facility and the Small Animal Imaging facility are both supported under the NIH/NCI award number P30CA016672. Technical assistance provided by Gloria Sierra and Sita Nookala for the TC-1 experiments is acknowledged.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Berman, T.A.; Schiller, J.T. Human papillomavirus in cervical cancer and oropharyngeal cancer: One cause, two diseases. Cancer 2017, 123, 2219–2229. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.A.; Jhingran, A.; Oaknin, A.; Denny, L. Cervical cancer. Lancet 2019, 393, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Gillison, M.L. Human papillomavirus in head and neck cancer: Its role in pathogenesis and clinical implications. Clin. Cancer Res. 2009, 15, 6758–6762. [Google Scholar] [CrossRef] [PubMed]
- Gillison, M.L.; Chaturvedi, A.K.; Anderson, W.F.; Fakhry, C. Epidemiology of Human Papillomavirus–Positive Head and Neck Squamous Cell Carcinoma. J. Clin. Oncol. 2015, 33, 3235–3242. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Jeang, J.; Cheng, K.; Cheng, T.; Yang, B.; Wu, T.-C.; Hung, C.-F. Current state in the development of candidate therapeutic HPV vaccines. Expert Rev. Vaccines 2016, 15, 989–1007. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Maraj, B.; Tran, N.P.; Knoff, J.; Chen, A.; Alvarez, R.D.; Hung, C.-F.; Wu, T.-C. Emerging human papillomavirus vaccines. Expert Opin. Emerg. Drugs 2012, 17, 469–492. [Google Scholar] [CrossRef] [PubMed]
- van Poelgeest, M.I.; Welters, M.J.; van Esch, E.M.; Stynenbosch, L.F.; Kerpershoek, G.; van Persijn van Meerten, E.L.; van den Hende, M.; Löwik, M.J.G.; Berends-van der Meer, D.M.A.; Fathers, L.M. HPV16 synthetic long peptide (HPV16-SLP) vaccination therapy of patients with advanced or recurrent HPV16-induced gynecological carcinoma, a phase II trial. J. Transl. Med. 2013, 11, 88. [Google Scholar] [CrossRef]
- Akira, S. Innate Immunity and Adjuvants. Philos. Trans. R Soc. Lond. B Biol. Sci. 2011, 366, 2748–2755. [Google Scholar] [CrossRef]
- Dar, T.B.; Henson, R.M.; Shiao, S.L. Targeting Innate Immunity to Enhance the Efficacy of Radiation Therapy. Front. Immunol. 2018, 9, 3077. [Google Scholar] [CrossRef]
- Morvan, M.G.; Lanier, L.L. NK cells and cancer: You can teach innate cells new tricks. Nat. Rev. Cancer 2016, 16, 7–19. [Google Scholar] [CrossRef]
- Villegas, F.R.; Coca, S.; Villarrubia, V.G.; Jiménez, R.; Chillón, M.J.; Jareño, J.; Zuil, M.; Callol, L. Prognostic significance of tumor infiltrating natural killer cells subset CD57 in patients with squamous cell lung cancer. Lung Cancer 2002, 35, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, S.N.; Reighard, S.D.; Gyurova, I.E.; A Cranert, S.A.; E Mahl, S.E.; Karmele, E.P.; McNally, J.P.; Moran, M.T.; Brooks, T.R.; Yaqoob, F.; et al. Roles of natural killer cells in antiviral immunity. Curr. Opin. Virol. 2016, 16, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Pashine, A.; Valiante, N.M.; Ulmer, J.B. Targeting the innate immune response with improved vaccine adjuvants. Nat. Med. 2005, 11 (Suppl. 4), S63–S68. [Google Scholar] [CrossRef] [PubMed]
- Hermans, I.F.; Silk, J.D.; Gileadi, U.; Salio, M.; Mathew, B.; Ritter, G.; Schmidt, R.; Harris, A.L.; Old, L.; Cerundolo, V. NKT Cells Enhance CD4+ and CD8+ T Cell Responses to Soluble Antigen In Vivo through Direct Interaction with Dendritic Cells. J. Immunol. 2003, 171, 5140–5147. [Google Scholar] [CrossRef] [PubMed]
- Silk, J.D.; Hermans, I.F.; Gileadi, U.; Chong, T.W.; Shepherd, D.; Salio, M.; Mathew, B.; Schmidt, R.R.; Lunt, S.J.; Williams, K.J.; et al. Utilizing the adjuvant properties of CD1d-dependent NK T cells in T cell–mediated immunotherapy. J. Clin. Investig. 2004, 114, 1800–1811. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.H.; Lee, J.; Kline, L.; Mathison, J.C.; Ulevitch, R.J. Toll-like receptor 9 mediates CpG-DNA signaling. J. Leukoc. Biol. 2002, 71, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Soltysik, S.; Wu, J.-Y.; Recchia, J.; Wheeler, D.A.; Newman, M.J.; Coughlin, R.T.; Kensil, C.R. Structure/function studies of QS-21 adjuvant: Assessment of triterpene aldehyde and glucuronic acid roles in adjuvant function. Vaccine 1995, 13, 1403–1410. [Google Scholar] [CrossRef]
- Marciani, D.J. Elucidating the Mechanisms of Action of Saponin-Derived Adjuvants. Trends Pharmacol. Sci. 2018, 39, 573–585. [Google Scholar] [CrossRef]
- Sarkar, A.K.; Tortolero-Luna, G.; Follen, M.; Sastry, K.J. Inverse correlation of cellular immune responses specific to synthetic peptides from the E6 and E7 oncoproteins of HPV-16 with recurrence of cervical intraepithelial neoplasia in a cross-sectional study. Gynecol. Oncol. 2005, 99 (Suppl. 1), S251–S261. [Google Scholar] [CrossRef]
- Manuri, P.R.; Nehete, B.; Nehete, P.N.; Reisenauer, R.; Wardell, S.; Courtney, A.N.; Gambhira, R.; Lomada, D.; Chopra, A.K.; Sastry, K.J. Intranasal immunization with synthetic peptides corresponding to the E6 and E7 oncoproteins of human papillomavirus type 16 induces systemic and mucosal cellular immune responses and tumor protection. Vaccine 2007, 25, 3302–3310. [Google Scholar] [CrossRef]
- Sierra, G.; Dorta-Estremera, S.; Hegde, V.L.; Nookala, S.M.; Yanamandra, A.V.; Sastry, K.J. Intranasal Therapeutic Peptide Vaccine Promotes Efficient Induction and Trafficking of Cytotoxic T Cell Response for the Clearance of HPV Vaginal Tumors. Vaccines 2020, 8, 259. [Google Scholar] [CrossRef]
- Williams, R.; Lee, D.W.; Elzey, B.D.; Anderson, M.E.; Hostager, B.S.; Lee, J.H. Preclinical models of HPV+ and HPV− HNSCC in mice: An immune clearance of HPV+ HNSCC. Head Neck 2009, 31, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Dorta-Estremera, S.; Chin, R.L.; Sierra, G.; Nicholas, C.; Yanamandra, A.V.; Nookala, S.M.; Yang, G.; Singh, S.; Curran, M.A.; Sastry, K.J. Mucosal HPV E6/E7 Peptide Vaccination in Combination with Immune Checkpoint Modulation Induces Regression of HPV(+) Oral Cancers. Cancer Res. 2018, 78, 5327–5339. [Google Scholar] [CrossRef] [PubMed]
- Dorta-Estremera, S.; Hegde, V.L.; Slay, R.B.; Sun, R.; Yanamandra, A.V.; Nicholas, C.; Nookala, S.; Sierra, G.; Curran, M.A. Targeting interferon signaling and CTLA-4 enhance the therapeutic efficacy of anti-PD-1 immunotherapy in preclinical model of HPV(+) oral cancer. J. Immunother. Cancer 2019, 7, 252. [Google Scholar] [CrossRef]
- Decrausaz, L.; Gonçalves, A.; Domingos-Pereira, S.; Pythoud, C.; Stehle, J.; Schiller, J.; Jichlinski, P.; Nardelli-Haefliger, D. A novel mucosal orthotopic murine model of human papillomavirus-associated genital cancers. Int. J. Cancer 2011, 128, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Bartkowiak, T.; Singh, S.; Yang, G.; Galvan, G.; Haria, D.; Ai, M.; Allison, J.P.; Sastry, K.J.; Curran, M.A. Unique potential of 4-1BB agonist antibody to promote durable regression of HPV+ tumors when combined with an E6/E7 peptide vaccine. Proc. Natl. Acad. Sci. USA 2015, 112, E5290–E5299. [Google Scholar] [CrossRef] [PubMed]
- Steinhagen, F.; Kinjo, T.; Bode, C.; Klinman, D.M. TLR-based immune adjuvants. Vaccine 2011, 29, 3341–3355. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Kirschning, C.J.; Häcker, H.; Redecke, V.; Hausmann, S.; Akira, S.; Wagner, H.; Lipford, G.B. Human TLR9 confers responsiveness to bacterial DNA via species-specific CpG motif recognition. Proc. Natl. Acad. Sci. USA 2001, 98, 9237–9242. [Google Scholar] [CrossRef] [PubMed]
- Terme, M.; Mignot, G.; Ullrich, E.; Bonmort, M.; Minard-Colin, V.; Jacquet, A.; Schultze, J.L.; Kroemer, G.; Leclerc, C.; Chaput, N.; et al. The Dendritic Cell–like Functions of IFN-Producing Killer Dendritic Cells Reside in the CD11b+ Subset and Are Licensed by Tumor Cells. Cancer Res. 2009, 69, 6590–6597. [Google Scholar] [CrossRef]
- Lin, K.Y.; Guarnieri, F.G.; Staveley-O’Carroll, K.F.; Levitsky, H.I.; August, J.T.; Pardoll, D.M.; Wu, T.C. Treatment of established tumors with a novel vaccine that enhances major histocompatibility class II presentation of tumor antigen. Cancer Res. 1996, 56, 21–26. [Google Scholar]
- Moynihan, K.D.; Irvine, D.J. Roles for Innate Immunity in Combination Immunotherapies. Cancer Res. 2017, 77, 5215–5221. [Google Scholar] [CrossRef]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The prognostic landscape of genes and infil-trating immune cells across human cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef]
- Chaudhry, U.I.; Plitas, G.; Burt, B.M.; Kingham, T.P.; Raab, J.R.; DeMatteo, R.P. NK Dendritic Cells Expanded in IL-15 Exhibit Antitumor Responses In Vivo. J. Immunol. 2007, 179, 4654–4660. [Google Scholar] [CrossRef]
- Taieb, J.; Chaput, N.; Ménard, C.; Apetoh, L.; Ullrich, E.; Bonmort, M.; Péquignot, M.; Casares, N.; Terme, M.; Flament, C. A novel dendritic cell subset involved in tumor immunosurveillance. Nat. Med. 2006, 12, 214–219. [Google Scholar] [CrossRef]
- Cullen, S.P.; Brunet, M.; Martin, S.J. Granzymes in cancer and immunity. Cell Death Differ. 2010, 17, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Plitas, G.; Chaudhry, U.I.; Kingham, T.P.; Raab, J.R.; DeMatteo, R.P. NK Dendritic Cells Are Innate Immune Responders to Listeria monocytogenes Infection. J. Immunol. 2007, 178, 4411–4416. [Google Scholar] [CrossRef] [PubMed]
- Hancock, G.E.; Heers, K.M.; Smith, J.D. QS-21 Synergizes with Recombinant Interleukin-12 to Create a Potent Adjuvant Formulation for the Fusion Protein of Respiratory Syncytial Virus. Viral Immunol. 2000, 13, 503–509. [Google Scholar] [CrossRef]
- Mikloska, Z.; Ruckholdt, M.; Ghadiminejad, I.; Dunckley, H.; Denis, M.; Cunningham, A.L. Monophosphoryl lipid A and QS21 increase CD8 T lymphocyte cytotoxicity to herpes simplex virus-2 infected cell proteins 4 and 27 through IFN-gamma and IL-12 production. J. Immunol. 2000, 164, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Silla, S.; Fallarino, F.; Boon, T.; Uyttenhove, C. Enhancement by IL-12 of the cytolytic T lymphocyte (CTL) response of mice immunized with tumor-specific peptides in an adjuvant containing QS21 and MPL. Eur. Cytokine Netw. 1999, 10, 181–190. [Google Scholar]
- Lacaille-Dubois, M.-A. Updated insights into the mechanism of action and clinical profile of the immunoadjuvant QS-21: A review. Phytomedicine 2019, 60, 152905. [Google Scholar] [CrossRef]
- Iizuka, A.; Ikarashi, Y.; Yoshida, M.; Heike, Y.; Takeda, K.; Quinn, G.; Wakasugi, H.; Kitagawa, M.; Takaue, Y. Interleukin (IL)-4 promotes T helper type 2-biased natural killer T (NKT) cell expansion, which is regulated by NKT cell-derived interferon-gamma and IL-4. Immunology 2008, 123, 100–107. [Google Scholar] [CrossRef]
- Yin, S.; Wang, H.; Bertola, A.; Feng, D.; Xu, M.-J.; Wang, Y.; Gao, B. Activation of invariant natural killer T cells impedes liver regeneration by way of both IFN-gamma- and IL-4-dependent mechanisms. Hepatology 2014, 60, 1356–1366. [Google Scholar] [CrossRef]
- Semmling, V.; Lukacs-Kornek, V.; A Thaiss, C.; Quast, T.; Hochheiser, K.; Panzer, U.; Rossjohn, J.; Perlmutter, P.; Cao, J.; I Godfrey, D.; et al. Alternative cross-priming through CCL17-CCR4-mediated attraction of CTLs toward NKT cell–licensed DCs. Nat. Immunol. 2010, 11, 313–320. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Semmling, V.; Franken, L.; Wagner, H.; Kurts, C. Chemokines: A New Dendritic Cell Signal for T Cell Activation. Front. Immunol. 2011, 2, 31. [Google Scholar] [CrossRef]
- Cheng, W.F.; Hung, C.F.; Lin, K.Y.; Ling, M.; Juang, J.; He, L.; Lin, C.T. CD8+ T cells, NK cells and IFN-gamma are important for control of tumor with downregulated MHC class I expression by DNA vaccination. Gene Ther. 2003, 10, 1311–1320. [Google Scholar] [CrossRef]
- Young, H.A.; Hardy, K.J. Role of interferon-gamma in immune cell regulation. J. Leukoc. Biol. 1995, 58, 373–381. [Google Scholar] [CrossRef]
- Street, D.; Kaufmann, A.M.; Vaughan, A.; Fisher, S.G.; Hunter, M.; Schreckenberger, C.; Potkul, R.K.; Gissmann, L.; Qiao, L. Interferon-gamma enhances susceptibility of cervical cancer cells to lysis by tumor-specific cytotoxic T cells. Gynecol. Oncol. 1997, 65, 265–272. [Google Scholar] [CrossRef]
- Martini, M.; Innamorati, G.; Mazzocco, M.; Ugel, S.; Bronte, V.; Krampera, M.; Mosna, F.; Cestari, T.; Colombatti, M.; Sartoris, S. IFN-gamma-mediated upmodulation of MHC class I expression activates tumor-specific immune response in a mouse model of prostate cancer. Vaccine 2010, 28, 3548–3557. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; NI, J.; Meng, H.; Li, D.; Wei, Y.; Luo, Y.; Wu, Y. Interleukin-15: A potent adjuvant enhancing the efficacy of an autologous whole-cell tumor vaccine against Lewis lung carcinoma. Mol. Med. Rep. 2014, 10, 1828–1834. [Google Scholar] [CrossRef] [PubMed]
- Umemura, M.; Nishimura, H.; Saito, K.; Yajima, T.; Matsuzaki, G.; Mizuno, S.; Sugawara, I.; Yoshikai, Y. Interleukin-15 as an immune adjuvant to increase the efficacy of Mycobacterium bovis bacillus Calmette-Guerin vaccination. Infect. Immun. 2003, 71, 6045–6048. [Google Scholar] [CrossRef] [PubMed]
- Melchionda, F.; Fry, T.J.; Milliron, M.J.; McKirdy, M.A.; Tagaya, Y.; Mackall, C.L. Adjuvant IL-7 or IL-15 overcomes immunodominance and improves survival of the CD8+ memory cell pool. J. Clin. Investig. 2005, 115, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Xie, S.; Chen, M.; Li, Y.; Yue, J.; Ma, J.; Shu, X.; He, Y.; Xiao, W.; Tian, Z. Advances in NK cell production. Cell. Mol. Immunol. 2022, 19, 460–481. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Nakazawa, T.; Nakamura, M.; Nishimura, F.; Matsuda, R.; Omoto, K.; Shida, Y.; Murakami, T.; Nakagawa, I.; Motoyama, Y.; et al. Ex vivo-expanded highly purified natural killer cells in combination with temozolomide induce antitumor effects in human glioblastoma cells in vitro. PLoS ONE 2019, 14, e0212455. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Differential efficacy of TVAC vs. TVQC in the oral HPV tumor model. Syngeneic C57Bl/6 mice were injected with mEER tumor cells as indicated and treated with intranasal (IN) delivery of the indicated E6/E7 HPV peptide therapeutic vaccine formulations on days 5 and 11 post-tumor implantation (A). Untreated mice (UNTR) and mice vaccinated with single adjuvants (TVA, TVC, TVQ) served as controls. Mice in all the groups were monitored over time, and Kaplan–Meier survival curves are shown for mice treated with TVAC (B) and TVQC (C). The Mantel–Cox log-rank test, ** p < 0.005, **** p < 0.0001 (n = 5–25 mice per group), shows a significant regression of mEER oral tumors with the TVQC vaccine. The oral tumor volume for mice in the untreated and TVQC-treated groups, as determined using magnetic resonance imaging (MRI) analyses on day 18, showed a significant reduction in tumor size in vaccinated mice (D). Elevated frequencies of antigen-specific CD8 T cells were observed in the tumors with both vaccine formulations (E). Significance was determined using an ordinary one-way ANOVA * p < 0.05, *** p < 0.0005, **** p < 0.0001 (n = 5–6 mice per group).
Figure 1.
Differential efficacy of TVAC vs. TVQC in the oral HPV tumor model. Syngeneic C57Bl/6 mice were injected with mEER tumor cells as indicated and treated with intranasal (IN) delivery of the indicated E6/E7 HPV peptide therapeutic vaccine formulations on days 5 and 11 post-tumor implantation (A). Untreated mice (UNTR) and mice vaccinated with single adjuvants (TVA, TVC, TVQ) served as controls. Mice in all the groups were monitored over time, and Kaplan–Meier survival curves are shown for mice treated with TVAC (B) and TVQC (C). The Mantel–Cox log-rank test, ** p < 0.005, **** p < 0.0001 (n = 5–25 mice per group), shows a significant regression of mEER oral tumors with the TVQC vaccine. The oral tumor volume for mice in the untreated and TVQC-treated groups, as determined using magnetic resonance imaging (MRI) analyses on day 18, showed a significant reduction in tumor size in vaccinated mice (D). Elevated frequencies of antigen-specific CD8 T cells were observed in the tumors with both vaccine formulations (E). Significance was determined using an ordinary one-way ANOVA * p < 0.05, *** p < 0.0005, **** p < 0.0001 (n = 5–6 mice per group).

Figure 2.
TVQC induces a more robust innate effector response than TVAC in mice with oral HPV tumors. Syngeneic C57Bl/6 mice were injected with mEER tumor cells as indicated and treated with intranasal (IN) delivery of TVAC, TVQC or untreated (UNTR). TVQC induced significantly elevated frequencies of total and functional (granzyme B- and/or IFNg-expressing) NK cells (A) and NKDCs (B) in the TME. Significance was determined using an ordinary one-way ANOVA * p < 0.05, ** p < 0.005 (n = 5–6 mice per group). Within the TVQC-vaccinated mice, the total and functional CD11c−/+ NK cells were compared with the CD11c+ subset (NKDCs) exhibiting a higher frequency and functionality in the TME (C). Significance was determined using a Wilcoxon matched pairs test **** p < 0.0001, *** p < 0.0001 (n = 19 mice).
Figure 2.
TVQC induces a more robust innate effector response than TVAC in mice with oral HPV tumors. Syngeneic C57Bl/6 mice were injected with mEER tumor cells as indicated and treated with intranasal (IN) delivery of TVAC, TVQC or untreated (UNTR). TVQC induced significantly elevated frequencies of total and functional (granzyme B- and/or IFNg-expressing) NK cells (A) and NKDCs (B) in the TME. Significance was determined using an ordinary one-way ANOVA * p < 0.05, ** p < 0.005 (n = 5–6 mice per group). Within the TVQC-vaccinated mice, the total and functional CD11c−/+ NK cells were compared with the CD11c+ subset (NKDCs) exhibiting a higher frequency and functionality in the TME (C). Significance was determined using a Wilcoxon matched pairs test **** p < 0.0001, *** p < 0.0001 (n = 19 mice).
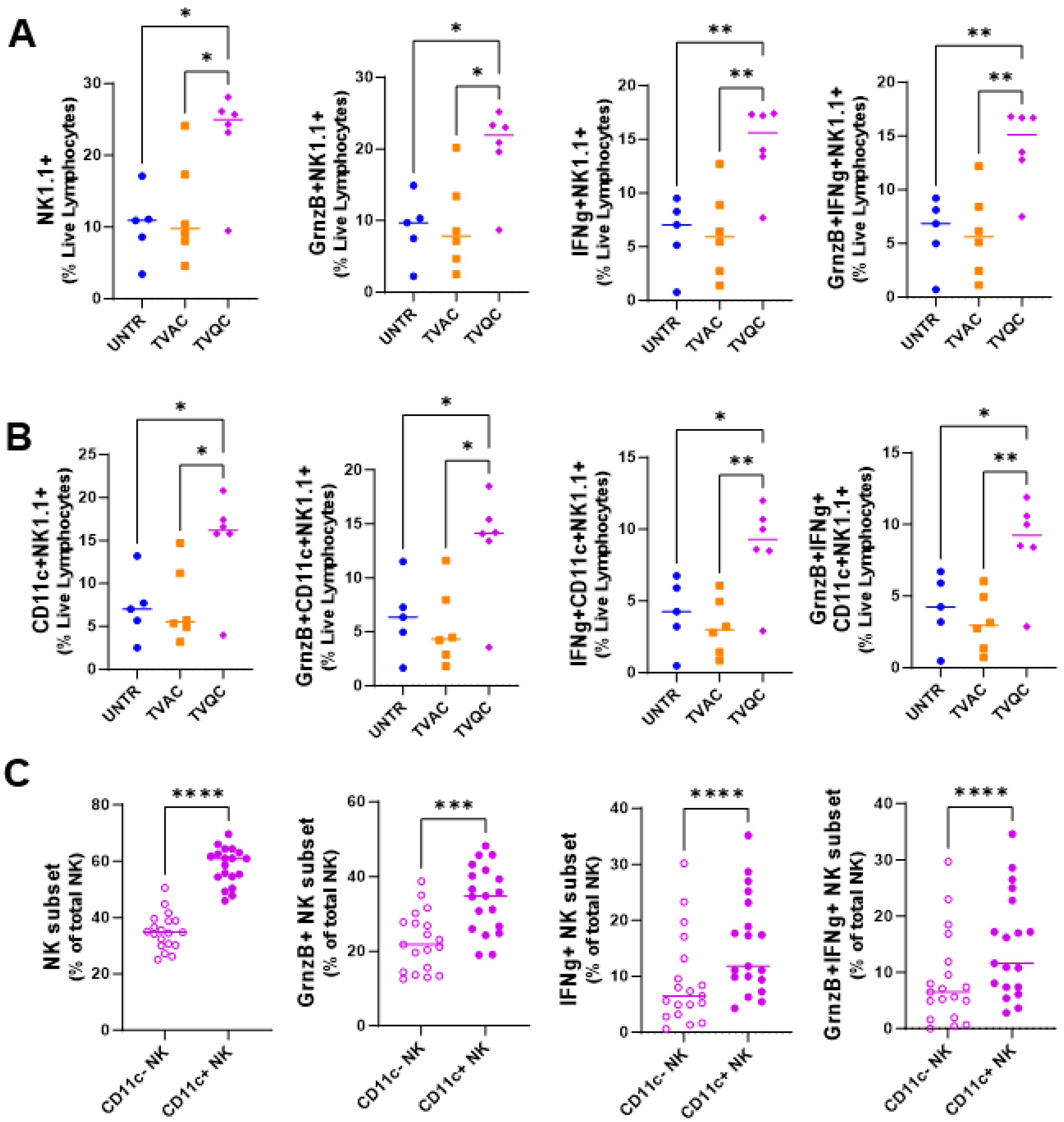
Figure 3.
Syngeneic C57BL/6 female mice were implanted with intravaginal TC-1luc tumors, as described in the Materials and Methods, and treated with intranasal HPV peptide therapeutic vaccine formulations as indicated on days 5 and 11 post-tumor implantation (A). Intravaginal TC-1 tumor size was monitored by luciferase imaging using the IVIS bioluminescence imaging system, and tumor growth curves for the indicated groups are shown (B). Survival curves are shown for intravaginal TC-1 tumor-bearing mice as indicated (C). A Mantel–Cox log-rank test was used to determine the significance between the groups, * p < 0.05, ** p < 0.005 (n = 10–13 mice per group). Immune responses associated with vaccine efficacy were assessed by flow cytometric analysis of TILs on day 16 following tumor implantation. The frequencies of the total and IFNg+ E7 tetramer+ CD8 T cells (D) and NKDCs expressing granzyme B, IFNg or both are shown (E). Significance was determined using an unpaired t-test, * p < 0.05, ** p < 0.01 (representative data from one experiment with n = 3 mice per group; the experiment was repeated one more time).
Figure 3.
Syngeneic C57BL/6 female mice were implanted with intravaginal TC-1luc tumors, as described in the Materials and Methods, and treated with intranasal HPV peptide therapeutic vaccine formulations as indicated on days 5 and 11 post-tumor implantation (A). Intravaginal TC-1 tumor size was monitored by luciferase imaging using the IVIS bioluminescence imaging system, and tumor growth curves for the indicated groups are shown (B). Survival curves are shown for intravaginal TC-1 tumor-bearing mice as indicated (C). A Mantel–Cox log-rank test was used to determine the significance between the groups, * p < 0.05, ** p < 0.005 (n = 10–13 mice per group). Immune responses associated with vaccine efficacy were assessed by flow cytometric analysis of TILs on day 16 following tumor implantation. The frequencies of the total and IFNg+ E7 tetramer+ CD8 T cells (D) and NKDCs expressing granzyme B, IFNg or both are shown (E). Significance was determined using an unpaired t-test, * p < 0.05, ** p < 0.01 (representative data from one experiment with n = 3 mice per group; the experiment was repeated one more time).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
O’Hara, M.P.; Yanamandra, A.V.; Sastry, K.J. Immunity from NK Cell Subsets Is Important for Vaccine-Mediated Protection in HPV+ Cancers. Vaccines 2024, 12, 206. https://doi.org/10.3390/vaccines12020206
AMA Style
O’Hara MP, Yanamandra AV, Sastry KJ. Immunity from NK Cell Subsets Is Important for Vaccine-Mediated Protection in HPV+ Cancers. Vaccines. 2024; 12(2):206. https://doi.org/10.3390/vaccines12020206
Chicago/Turabian StyleO’Hara, Madison P., Ananta V. Yanamandra, and K. Jagannadha Sastry. 2024. "Immunity from NK Cell Subsets Is Important for Vaccine-Mediated Protection in HPV+ Cancers" Vaccines 12, no. 2: 206. https://doi.org/10.3390/vaccines12020206
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.