
Application of Desert DSEs to Nonhost Plants: Potential to Promote Growth and Alleviate Drought Stress of Wheat Seedlings
1
School of Life Sciences, Hebei University, Baoding 071002, China
2
Key Laboratory of Microbial Diversity Research and Application of Hebei Province, Baoding 071002, China
*
Author to whom correspondence should be addressed.
Agriculture 2022, 12(10), 1539; https://doi.org/10.3390/agriculture12101539
Submission received: 12 August 2022
/
Revised: 12 September 2022
/
Accepted: 21 September 2022
/
Published: 24 September 2022
(This article belongs to the Special Issue High Yield Cultivation, Growth and Development Mechanism of Wheat)
Abstract
:To evaluate the applicability of desert dark septate endophytes (DSEs) in crop cultivation, Alternaria alternata (Fr.) Keissl. (Aa), Paraphoma pye Moslemi & P.W.J. Taylor (Pp), and Paraphoma radicina (McAlpine) Morgan-Jones & J.F. White (Pr) were inoculated into nonhost wheat growing under three water conditions. The plants’ biomass, vegetative growth, and physiological parameters were investigated. At harvest, all DSE strains were effective colonizers under all treatments. These DSEs generally positively affected wheat growth but varied among different DSE species, and this promoting effect was more obvious under drought conditions. Under mild drought (MD) treatments, Aa and Pr increased the leaf number and plant height of hosts. Pr showed a significant beneficial influence on the wheat’s total biomass under all treatments, while Aa and Pp showed benefits only under the well-watered or MD treatment. The response mechanisms of DSE-inoculated wheat under stress may be due to the enhanced photosynthetic efficiency and antioxidant system. Generally, Pr had a stronger beneficial effect. The improved growth and fitness of the inoculated hosts under drought stress may reduce their water supply requirements during seedling growth. We speculate that inoculating wheat with this strain could be a promising approach for water-saving cultivation in arid environments.
1. Introduction
An increased intensity and frequency for water deficit stress is expected under global climate change, and adverse effects on a larger spatial scale have been observed [1,2]. It is estimated that global agricultural suffering from drought stress will have a twofold increase in the 21st century [3]. In China, the world’s largest producer of cereal crops, wheat and other important grain crops are severely reduced under drought conditions [4,5]. Triticum aestivum (wheat) belongs to Poaceae, contains starch, protein, and sugar, and is one of the major grains worldwide [6,7,8]. Drought stress can affect wheat growth at all stages, and the seedling stage is usually the most vulnerable period throughout the whole life of the plant, so it is more sensitive to external drought stress [7,9,10]. For example, drought stress applied to wheat seedlings can lead to a half-root-length reduction [11]. A more severe level of water deficit stress will become a common phenomenon in the future [3,7,12]. Figuring out how to improve wheat growth and stress resistance by sustainable methods is extremely urgent. In addition to traditional breeding and genetic engineering methods, the development of microbial technologies for green agriculture has become an important strategy to improve plants’ stress resistance [8,13,14,15,16,17]. Besides, chemically synthesized fertilizer was added around the world to overcome the food shortages. These have left a negative impact on the soil environment and influenced food security and sustainable development [18]. Microbes, especially fungi that harbored plant tissues endophytically, were considered as an environmentally friendly approach [19,20].
Fungi and plants usually form common coexistence relationships in nature [21,22,23,24]. Research on improving plants’ drought resistance through fungi has been widely reported. For example, shoot endophytes and root mycorrhizal fungi can promote host growth under drought stress and can alleviate the adverse effects caused by droughts [25,26,27]. In addition, dark septate endophytes (DSEs) have also been observed in various plants in recent decades [28,29]. DSEs are fungi with characteristic melanized septum hyphae and microsclerotium structures that colonize the roots of healthy plants (both intercellularly and intracellularly) [30,31]. With weak host specificity, DSEs are frequent colonizers of several plants that are widely distributed, especially in drought, cold, high altitude, wetland, heavy metal polluted and other adverse environments [19,32,33,34,35,36,37,38,39]. According to conservative estimates, DSEs have been found in at least 600 species of plants (including nonmycorrhizal plants) around the world [30], and it has been reported that DSEs are more widely distributed than mycorrhizal fungi in stressful environments [40]. Studies have shown that DSEs can not only be purely cultured in vitro but also improve the absorption of soil water, nitrogen, and phosphorus by plants, promoting hosts’ survival adaptability under adverse conditions [41,42,43,44,45,46,47,48,49,50].
Research on DSEs has shown that they are common colonizers in dry areas and are beneficial to hosts to resist water deficits [35,41,51,52]. Wu et al. [41] found DSE colonization in all 20 plant species in a desert area in China, and the DSE colonization rate was higher in dominant plant species in sandy land areas. The wide distribution of DSEs in arid areas indicates their potential role in the adaptation of arid vegetation to dry soils. Perez-naranjo et al. [53] artificially inoculated five DSEs into desert grasses under water stress and found that four DSEs promoted plant growth. Zuo et al. [54] showed that desert DSEs promoted the growth of Hedysarum scoparium and Ammopiptanthus mongolicus under drought conditions. The extended DSE hyphae around host roots have also been proposed to benefit plant survival in drought soils by improving water absorption [51,55]. In studies using crops as materials, DSEs can reduce the damage of antioxidant enzyme activity induced by droughts to rice plants and promote their growth [55].
The extensive distribution of DSEs in various natural and artificial ecosystems and the low host specificity indicate that DSEs have the potential to be applied in nonhost plants. There are many reports related to DSEs isolated from non-crops that have been proven to be able to successfully colonize rice, tomato, and maize and be beneficial to host plants [39,42,56]. For example, three DSEs isolated from the desert plant H. scoparium promoted maize growth and development under drought stress by altering root development [57]; two DSE strains from Ageratina adenophora and Buddleja officinalis roots successfully colonized tomato seedlings and improved host tolerance to Zn and Cd stress [39]. DSEs isolated from Vochysia divergens improved nitrogen and phosphorus nutrition and the photosynthesis of cowpea growing in salt conditions and promoted host tolerance to stress [58]. Considered the growing global food demand and decreasing amount of cultivated land, DSEs were selected to evaluate their potential application in crop cultivation. However, knowledge of the effects of desert plant DSEs on nonhost species and their application potential is still needed.
Characterization of new fungi, especially from extreme environments, would not only help in promoting the fitness of desert plants but also expand their application scope in agriculture. Psammochloa villosa (Poaceae) is a desert grass distributed in northern China and is also an important dominant forage grass in this area [59,60]. During long-term evolution, it has developed morphological and physiological characteristics suitable for arid habitats and has a strong sand-fixation ability, which is an important resource for desert restoration [37,60]. Hou et al. [37] observed DSEs in P. villosa roots naturally grown in the deserts of northern China. The preliminary investigation found that DSE fungi widely existed in the roots of P. villosa, indicating unique characteristics of adaptation to drought environments, which need to be further studied. As mentioned above, plant endophytic fungi living in arid habitats can not only help the original host to resist the stressed soil environment, but also give the non-host plants (such as crops) stress tolerance to specific habitats, which provides a new idea for improving the drought resistance of crops. Combined with their characteristics of a unique distribution habitat, low host specificity, and pure culture in vitro, we speculated that DSEs are not only an important choice for desert restoration but also benefit plant fitness under dry soils, which are vital to the modern agricultural development.
To explore the DSE–plant interactions and using DSE inoculants for crop cultivation, three DSE strains, Alternaria alternata (Aa), Paraphoma pye (Pp), and Paraphoma radicina (Pr), previously obtained from P. villosa in Northwest China, were artificially inoculated into wheat seedlings. The inoculation experiment was conducted under three water treatments in a greenhouse. The plant growth and fitness indexes under different water treatments were studied to evaluate the potential of wheat and DSE interactions to improve wheat growth at the seedlings stage and to provide a new solution to improve the productivity of wheat under drought conditions.
2. Materials and Methods
2.1. Fungal Strains and Plant Cultivation
Three DSEs previously obtained from Psammochloa villosa roots were selected as fungal inoculants. These DSE strains were phylogenetically identified as Alternaria alternata (Aa), Paraphoma pye (Pp), and Paraphoma radicina (Pr) based on ITS4-5.8S-ITS5 genes (GenBank accession MK246179, MT138911, KX197926) [37,61]. These strains were inoculated in flasks containing modified Melin-Norkrans (MMN) fluid medium (1000 mL distilled water contains 15.0 g of glucose, 0.5 g of KH2PO4, 0.25 g of (NH4)2HPO4, 0.2 g of citric acid, 0.15 g of MgSO4·7H2O, 0.05 g of CaCl2, 0.025 g of NaCl, 100 µg of VB1, and 1.2 mL of FeCl3 (1%); pH 5.5) as previously described by Zuo et al. and were cultured at 27 °C and 120 rpm for 14 d [62]. After culturing, the mycelia were washed several times using sterile water to ensure that there was no residual culture fluid on the mycelia. Approximately 5 g of each strain was weighed and thoroughly mixed in sterile water.
The inoculation experiment used seeds of wheat, cultivar “Jimai 22”, obtained from Hebei Agricultural University. Plump-eared and uniformly sized wheat seeds were selected for surface disinfection with 3% NaClO and washed with sterilized water. The sterilized seeds were soaked in sterile water in petri dishes for pregermination to ensure a consistent initial growth. Two days later, five seedlings of the same growth status were transferred into one pot containing 3 kg of substrate. The growth substrate was surface farmland soil (5–20 cm), which was sieved (2 mm) to remove impurities and then stored. The experimental soil contained 80.68 mg/kg available nitrogen, 20.35 mg/g organic matter, and 59.70 µg/g available phosphorus.
2.2. Experimental Design
A completely random two-factor experiment was designed. DSE inoculation was the first factor, which included the noninoculated control group (CK) and inoculation with Aa, Pp, and Pr. The second factor contained three water treatments: well-watered (WW), mild drought stress (MD) and severe drought stress (ED) treatments; that is, the soil water content was kept at 70%, 50%, and 30%, respectively, of the field water capacity. Each treatment had 5 replicates. For the DSE inoculation treatment, 50 mL inoculum was added to each pot, and the same volume of sterile inoculum was used in the control treatment. Then, the wheat seedlings were cultured in a greenhouse (day/night 16 h/8 h, 28 °C/22 °C, humidity 55%), and the plants were grown under a soil water content at 70% field capacity. After two weeks of growth, different water treatments were applied. Specifically, two-thirds of the seedlings retained 50% and 30% of the soil water content as the drought treatment, and the other group given a good supply of water served as the control. Then, the water treatment was retained by measuring the soil moisture every day. The seedlings were irrigated by the weighing method to achieve the corresponding level of drought treatment for 30 d. Then, the seedlings were harvested immediately. The position of the culture pot was randomly adjusted periodically to ensure that the growth difference of the wheat was independent of the position.
2.3. Photosynthetic Indexes and Chlorophyll Content
Before harvest, the leaves’ chlorophyll content was measured using the first completely mature leaf from the apex (Spad-502, Konica Minolta Sensing, Inc., Osaka, Japan) [63]. This analyzer recorded the maximal absorbance values of Chl a and b at 650 nm. Similarly, we used a gas analyzer (LI-6400XT, Licor, Lincoln, NE, USA) to determine the leaf photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), and intercellular CO2 concentration (Ci). Readings were taken between 9 and 11 h under a natural temperature, CO2 concentration, and saturated light (1500 μmol m−2 s−1) [64].
2.4. Growth Parameters
The value of leaf numbers and plant height of every seedling was recorded. Then, the third and fourth leaves in one pot were picked and immediately frozen at −80 °C for physiological index analysis. Then, the shoot and root samples were harvested separately. The roots in each pot were washed and weighed, and partial fresh roots were used for the DSE colonization status. The rest of the parts were placed in an oven for 2 days (70 °C) for biomass measurement.
2.5. DSE Colonization in Wheat Roots
To confirm the success of the inoculation experiment, several root segments were randomly selected and cut into 1 cm pieces and washed using tap water. Subsequently, these roots were placed in 10% (w/v) KOH solution at 90 °C. The root segments were then soaked in acid fuchsine for staining as previously described [65,66]. Approximately 30 root fragments for each sample were used to record the DSE structure using a light microscope (OLYMPUS-BX51, Tokyo, Japan) at × 400 magnification.
2.6. Physiological Indexes
The wheat leaves in each pot were ground in a chilled homogenization buffer to determine the physiological indexes. As described previously, the NBT photochemical reduction method was selected to test superoxide dismutase (SOD) activity [67]. The glutathione (GSH) content was measured according to the DTNB method [68]. The thiobarbituric acid method was used to determine the content of malondialdehyde (MDA) [69]. The leaf proline was detected using the acid ninhydrin colorimetric method [70]. In addition, the soluble protein and sugar contents were determined by the G205 Coomassie brilliant blue and anthrone colorimetry methods. Root auxin was determined by a plant auxin (IAA) ELISA kit (Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China) [71,72].
2.7. Statistical Analysis
The influences of the DSE and water treatments on the vegetative growth and physiological indexes were evaluated according to the two-way ANOVA with Tukey’s test at p < 0.05. Before the ANOVAs were conducted, the data were verified for the normality and homogeneity via the Shapiro–Wilk and Levene’s tests. The above analyses were conducted using SPSS 22.0 (SPSS Inc., Chicago, IL, USA). Principal component analysis (PCA) of the wheat growth indexes was performed using Canoco software (version 5, Microcomputer Power, Ithaca, NY, USA).
3. Results
3.1. DSE in the Wheat Roots
At the end of the experiment, we did not observe DSE hyphae or microsclerotia in the noninoculated roots, while DSE structures were found in the root tissues of all DSE-infected seedlings (Figure 1), and the total colonization rate was 24–55%.
3.2. Influences of DSE Inoculation on the Growth of Wheat Seedlings
DSE colonization, water treatment, and their interaction had significant influences on the wheat’s vegetative parameters (Table 1). As drought treatment intensified, the leaf number and plant height of the wheat seedlings decreased, but DSE inoculation alleviated that damage. Under WW conditions, DSE did not exhibit obvious influences on the hosts, while the leaf number and plant height of wheat inoculated with Aa and Pr under the MD and ED treatments were greater than those of the noninoculated wheat (Figure 2a,b).
The shoot biomass of wheat seedlings inoculated with Pr increased significantly under the WW and MD treatments, while Pp showed obvious benefits under the MD treatment (Figure 2c). For root biomass, the influences of DSE on the host were more beneficial under the dry treatment. The root biomass of seedlings inoculated with Pr increased significantly under the WW treatment, while Aa enhanced this value for the hosts under the MD treatment. Under ED treatment, all inoculated plants had significantly higher root biomass values than those of the control (Figure 2c). For total biomass, Pr inoculation had significant beneficial effects on this value for the hosts under all treatments, while Pp and Aa only showed benefits under ND and/or MD conditions (Figure 2d).
Under WW conditions, the auxin content of Aa-inoculated and Pp-inoculated seedlings was significantly higher. Under drought stress, the auxin content of plants inoculated with DSE was significantly increased, among which the value of Pr-inoculated plants was significantly increased by 64.1% under MD stress, and the auxin content of Aa-, Pp-, and Pr-inoculated plants was significantly increased by 42.0–49.8% compared with plants that did not harbor DSE under ED stress (Figure 2e).
3.3. Influences of DSE Inoculation on the Antioxidant Enzyme System of Wheat Seedlings
DSE inoculation and water treatment significantly influenced the activity of SOD and the MDA and GSH contents in the wheat leaves (Table 2). Under WW treatment, most DSEs did not affect the antioxidant enzyme system in leaves, except that the GSH content in the leaves was significantly increased by Pp inoculation. Under ED treatment, the GSH content in Aa-wheat was significantly increased compared to that of the uninfected control (Figure 3a). Under MD and ED treatments, DSE inoculation significantly promoted the activity of SOD, and this value in wheat leaves inoculated with Aa and Pr was significantly increased by 33.2–53.9% (MD) and 32.3–53.9% (ED), respectively, compared with that of the control treatment (Figure 3b). The Pp strain significantly enhanced SOD activity in wheat leaves under MD conditions. The MDA content of wheat was significantly decreased compared with that of the control under drought treatments. The MDA content in leaves inoculated with Pr under MD and ED stress was significantly reduced by 36.5% and 53.2%, respectively, compared with that of the uninoculated plants, and the MDA values of Pp-inoculated seedlings was also significantly decreased under ED conditions (Figure 3c).
3.4. Influences of DSE Inoculation on the Soluble Protein, Soluble Sugar, and Proline Contents of Wheat Leaves
Under the WW and MD treatments, the soluble protein content of DSE-inoculated plants increased. Aa and Pp inoculation significantly increased the soluble protein content of plants under the WW treatment compared with the noninoculated control, while the soluble protein values of Pp-inoculated wheat was significantly higher under MD and ED treatments (Table 2, Figure 4a). When drought stress intensified, the content of leaves’ soluble sugars increased gradually, and DSE inoculation further increased this value (Figure 4b). Under all treatments, Aa and Pr significantly enhanced the soluble sugar values of host plants, and Pp exhibited significant benefits only under the MD and ED treatments. The content of proline in the leaves of plants inoculated with DSE increased under the three water treatments. The proline content of Pp-inoculated and Pr-inoculated wheats under the WW treatment and of wheat inoculated with Aa, Pp, and Pr under the MD and ED treatments increased significantly compared with the control. Under the ED treatment, the proline values of DSE-inoculated wheat increased by 49.7–111.7% compared with the control (Figure 4c).
3.5. Influences of DSE Inoculation on Wheat Seedling Performance Based on Principal Component Analysis (PCA)
Under different water conditions, the separation between DSE inoculation and CK treatments was clear along the PCA axis, indicating that different DSE inoculations had a significant effect on the wheat seedlings (Figure 5). The plant height, SOD activity, soluble protein content, Pn, and Gs of wheat seedlings under WW conditions were the main indexes that differed among the different treatments (Figure 5a). Under MD conditions, the SOD activity, MDA content, Pn, Gs, IAA, soluble sugar, and proline contents in wheat were the main indexes that differed among the different treatments (Figure 5b). Under ED conditions, the GSH, soluble sugar, IAA, and proline contents and Pn, Gs, and Ci were the main indexes that differed between the inoculation treatment and the control (Figure 5c).
3.6. Influences of DSE on Leaf Photosynthetic Indexes and Chlorophyll Content
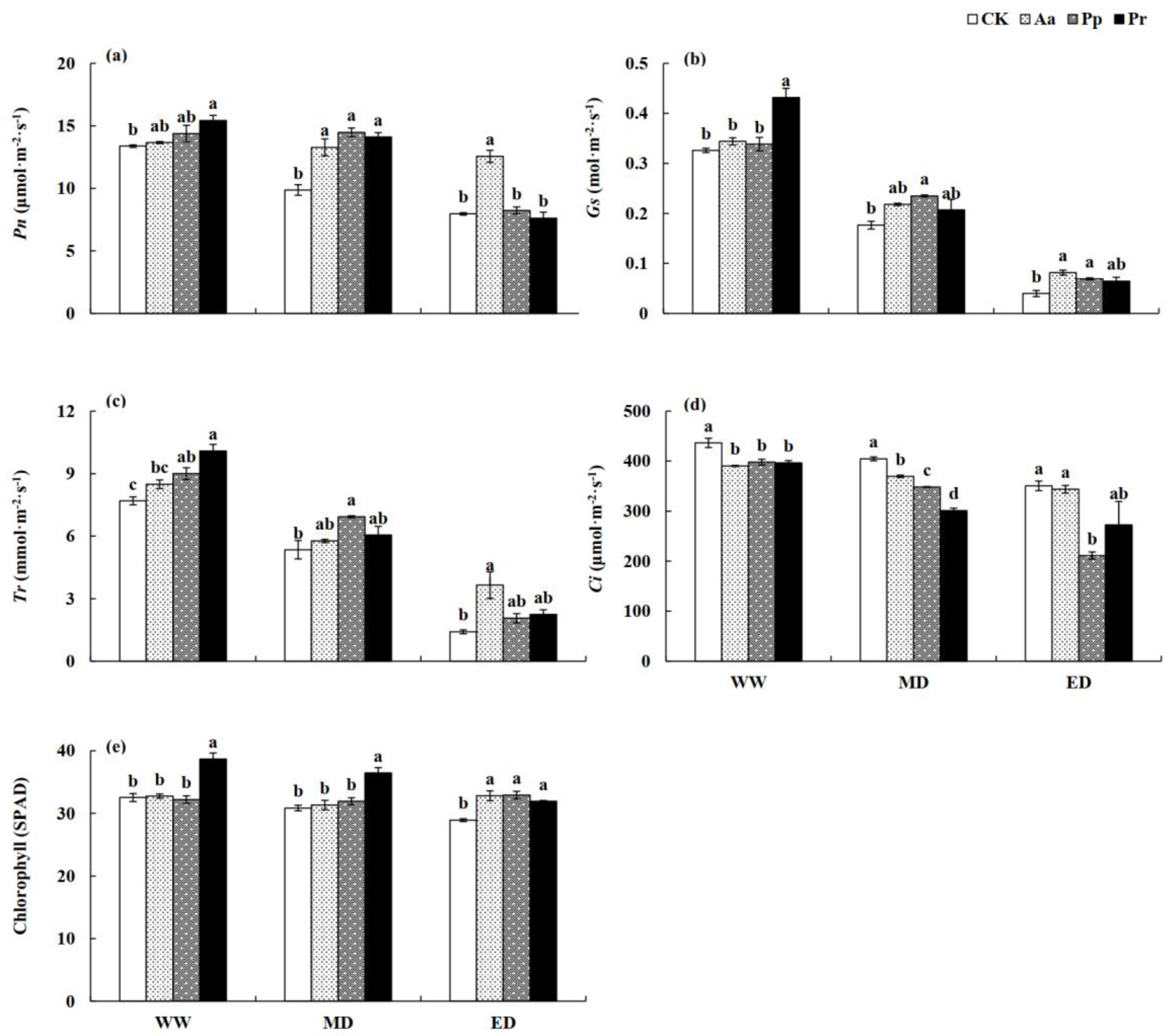
DSE and water treatment significantly affected the photosynthesis indexes and chlorophyll content in the wheat leaves (Table 3). Under WW treatment, the Pn of wheat plants inoculated with Pr increased significantly, while the photosynthetic rate in response to inoculation with the other strains did not show an obvious increase (Figure 6a). The effect of DSE on the photosynthetic rate of wheat was more significant under drought stress, especially under the MD treatment. The Pn of wheat inoculated with DSE significantly increased, by 34.4–46.7%, compared with the control. Under ED treatment, the photosynthetic rate of wheat inoculated with Aa increased significantly (Figure 6a). With an intensified water deficit (Figure 6b,c), the values of leaves’ Gs and Tr decreased gradually, but under all treatments, these values of wheat leaves increased due to DSE inoculation. Under well-watered conditions, Pr promoted the value of leaves’ Gs, and the Tr of DSE-inoculated seedlings was higher than that of the control (except for Aa). The transpiration rate and stomatal conductance of Pp-inoculated plants under the MD treatment and Aa- and Pp-inoculated plants under the ED treatment increased significantly (Figure 6b,c). The Ci of wheat leaves decreased due to DSE inoculation, and significant effects were observed under the WW and MD treatments. Under the ED treatment, the effect of DSE inoculation was reduced, and the value of Ci did not show a significant difference compared with that of the control, except for the Pp inoculation condition (Figure 6d).
Under the WW and MD treatments, the chlorophyll content of wheat leaves inoculated with P significantly increased compared with that of the control, while the other DSE strains had no significant effects. Under the ED treatment, seedlings harboring DSE showed greater chlorophyll values than that of the control (Figure 6e).
4. Discussion
4.1. Application Potential of DSE Inoculants for Nonhost Plants
DSEs are characterized by their wide distribution and low host specificity, making it possible to apply them to other host plants. This approach has been widely reported for other microorganisms [73,74]. However, most reports on the effects of DSEs on nonhost plants focus on the host’s resistance to heavy metals and disease resistance, while there are few studies on the plant’s response to drought stress [51,75]. In previous inoculation studies, we confirmed that desert DSE fungi could colonize new desert plants that grow in the same habitats. In the current study, we further indicate that DSEs from the root system of desert plants successfully colonized wheat roots under all water treatments, indicating that they may have a certain application potential. At the same time, the effect of DSEs on nonhost wheat was similar to that presented in a previous report; that is, the effect was influenced by the DSE species and water supply [30,51]. After harvest, most DSE fungi benefit wheat growth and stress resistance, but this promoting effect was more obvious under drought conditions. Combined with previous reports, this seems to be a relatively common phenomenon. The reason may be that symbiotic fungi originating from arid habitats prefer to benefit host plants in environments similar to their original habitats, which may explain why some DSE fungi are considered adverse fungi [47]. In summary, it is important to consider the original background of the DSEs in future studies.
4.2. Desert DSEs Affected the Growth of Nonhost Plants
In the symbiosis between fungi and plants, fungi obtain places and substances to grow such as carbohydrates from the host plants, while fungi provide a variety of beneficial effects for the host [34,76,77]. In this experiment, Aa and Pr significantly promoted the leaf growth and increased the plant height of wheat seedlings. Leaves are the main organs of photosynthesis, and chloroplasts are the main organelles of photosynthesis. Photosynthetic pigments are vital for plants with regard to light absorption and utilization [64]. Many external environments affect plant photosynthesis and growth by affecting chlorophyll biosynthesis [78,79]. In this study, the chlorophyll content in the CK-treated wheat leaves reduced markedly under the MD condition; however, the leaf chlorophyll content of wheat harboring DSEs was not significantly affected. These results are similar to that of a previous report about plants harboring AM fungi under drought conditions [80]. In addition, drought stress usually reduces the values of Pn, Gs, and Tr in wheat leaves [81]. However, DSE enhanced the photosynthetic rate of wheat leaves under MD conditions in the present study. Moreover, the DSEs significantly reduced the intercellular CO2 concentration, which indicated that the DSEs could alleviate the damage induced by drought on the photosynthetic processes [64]. Similar results have been observed in studies on rice inoculated with DSEs under water deficit conditions. These findings indicated that DSEs influence the maintenance of the photosynthetic system in the host.
Plant phytohormones are usually closely related to growth, physiological regulation, and resistance to environmental stresses and participate in all stages of plant growth [82,83]. For example, plant photosynthesis, nutrient absorption and other physiological metabolic processes are regulated by plant hormones [84]. In this study, the auxin content in wheat inoculated with most DSEs was higher than that of the control under well-watered conditions, and the increase in auxin under drought stress caused by inoculation with DSEs was enhanced, indicating that DSEs may promote wheat growth and improve wheat tolerance to drought stress by increasing the auxin content in wheat roots. Auxin is an important regulatory factor in plant growth, development, and resistance to drought [85]. Studies have shown that endophytic fungi have the ability to synthesize auxin and enhance the tolerance of host plants to abiotic stresses [86]. The accumulation of phytohormones such as IAA content helps to reduce transpiration loss in plants growing in dry soil [64]. In other crops, the endophytic fungi Alternaria alternata and Fusarium tricinctum have been found to secrete auxin and promote the growth and development of rice roots and shoots [87]. However, the molecular basis of DSE-mediated phytohormone accumulation under drought conditions needs to be tested further.
4.3. The Influence of Desert DSEs on the Drought Resistance of Nonhost Wheat
A lack of water usually negatively affects plant growth, including reactive oxygen species (ROS) accumulation, inducing membrane lipid peroxidation and oxidative damage. However, plants can produce antioxidants to reduce damage [88]. Among them, SOD is an important antioxidant enzyme in plants [89]. Endophytes help hosts tolerate dry conditions by inducing antioxidant responses in host plants [90]. In this study, the SOD activity in wheat leaves increased under MD and ED treatments after DSE inoculation. This affect has also been observed in other fungi. Tyagi et al. [91] found that AM fungi significantly increased the SOD activity of wheat plants under drought stress. Radhakrishnan et al. [92] also showed that the inoculation of endophytic fungi increased the SOD activity of soybean under salt stress. The MDA content reflects the degree of lipid oxidation of the plant membrane [93]. The decreased MDA content of plants harboring DSEs in this study suggested that these fungi reduce the oxidative damage to wheat under water deficit stress. He et al. [94] also reported that DSE reduced the content of MDA in leaves of Glycyrrhiza growing in drought soil. In addition, GSH plays an important role in eliminating ROS in plants grown in drought condition [95]. Sadeghi et al. [96] found that the inoculation of endophytic fungi under drought stress increased the GSH content of citrus plants. In this study, the strain Pp significantly promoted GSH production in wheat plants under moderate drought stress, indicating that DSE fungi could promote the production of GSH in wheat to remove reactive oxygen species and thus improve the drought resistance of wheat. Generally, DSEs can help hosts overcome water deficits by relieving oxidation harm.
When adapting to water deficits, plants produce more organic substances such as sugar and proline [97], and soluble sugar is usually an osmotic regulatory substance [98]. In this study, the soluble sugar content of wheat leaves increased with intensified drought conditions, and inoculation with DSE fungi further increased the soluble sugar content of wheat plants under all water treatments, especially inoculation with DSE fungi, which had the strongest effect on improving the soluble sugar content of wheat under severe drought stress. Thus, DSEs might improve the drought adaptability of wheat by promoting the accumulation of soluble sugars in wheat leaves. Pang et al. [99] found that endophytic fungi could increase the soluble sugar content of rice seedlings under drought stress. In addition, proline is one of the most effective affinity osmotic regulatory substances produced by plants during adaptations to drought stress [100,101]. In this study, inoculation with DSE fungi increased the proline content in wheat plants under normal water conditions and drought stress, and DSE fungi had the strongest effect on the proline content under severe drought stress. Moghaddam’s results also showed that Neocamarosporium sp. had a weak influence on crops’ proline content in the low drought treatment, but the opposite results were observed as the degree of drought increased [102]. These results suggest that the DSE fungi in this study may promote wheat growth and drought resistance by upregulating proline biosynthesis, maintaining the cell water status, and protecting the cell membrane and proteins from stress.
Generally, DSEs in desert areas have been widely investigated and reported. These fungi are usually inoculated into host and nonhost desert plants to test their application potential for vegetation restoration in dry soils. Considering the growing global food demand, three DSE strains in the current study, previously obtained from a desert plant, were selected to evaluate their potential application in crop cultivation. Most DSEs promoted nonhost wheat growth and resistance against water deficits under a simulation natural experiment, indicating that DSE colonization of wheat roots may promote seedling development in natural farm environments. The increased dry weight and photosynthetic ability of hosts suggests more organic matter and nutrients to support plant survival under drought conditions. In addition, the response mechanisms of DSE-inoculated wheat were consistent with those on desert plants, such as an enhanced production of antioxidant enzymes and an osmotic regulation mechanism. We therefore speculate that desert DSEs have the potential to promote wheat growth and therefore can play vital roles in changing the traditional method of crop cultivation in arid environments.
5. Conclusions
In conclusion, DSE strains isolated from desert plants colonized wheat roots under both well-watered and drought conditions, indicating that these three DSEs could be used as growth promoters in wheat planting. In this study, inoculation with DSE increased growth and biomass accumulation and alleviated drought damage in wheat seedlings. The improved drought resistance of the host may be related to the increased photosynthetic efficiency, leaf chlorophyll content, and enhanced antioxidant activity. Among them, Pr had a more beneficial influence on wheat growth under drought stress conditions. Considering the reduction in agricultural landscapes and the increasing human population, the demand for high wheat productivity will become more urgent. We speculate that inoculating wheat seedlings with DSE strains could be a promising approach for crop plants in arid environments. However, the application of inoculation technology in nature and the wheat–DSE interaction at other growth stages remain to be elucidated. Furthermore, the application of DSEs as a biofertilizer in wheat and the other crops will be further investigated.
Author Contributions
Conceptualization, X.L. and X.H.; methodology, X.L. and Y.L.; software, X.L. and Y.L.; validation, Y.L., Q.Y. and M.X.; formal analysis, X.H.; data curation, X.L.; writing—original draft preparation, Y.L.; writing—review and editing, X.L. and X.H.; funding acquisition, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation of Hebei Province, China (C2020201043); and the National Natural Science Foundation of China (31800345).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used in this research are all available upon request, and the accession numbers of DSEs in this study are presented in the “Materials and Methods”. Their sequences can be found in NCBI GenBank.
Acknowledgments
We appreciate the students Junmeng Long and Lifeng Hou for sampling and laboratory work. Thanks to Gansu Anxi Extreme-arid Desert National Nature Reserve for the support of the field investigation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Elkouk, A.; Pokhrel, Y.; Satoh, Y.; Bouchaou, L. Implications of changes in climate and human development on 21st-century global drought risk. J. Environ. Manag. 2022, 317, 115378. [Google Scholar] [CrossRef]
- Zachary, H.; Hoylman, R.; Bocinsky, K.; Kelsey, G.J. Drought assessment has been outpaced by climate change: Empirical arguments for a paradigm shift. Nat. Commun. 2022, 13, 2715. [Google Scholar]
- Zhao, T.; Dai, A. Uncertainties in historical changes and future projections of drought. Part II: Model-simulated historical and future drought changes. Clim. Chang. 2017, 144, 535–548. [Google Scholar] [CrossRef]
- Ghose, B. Food security and food self-sufficiency in China, from past to 2050. Food Energy Secur. 2014, 3, 86–95. [Google Scholar] [CrossRef]
- Jiang, J.; Huo, Z.; Feng, S.; Kang, S.; Wang, F.; Zhang, C. Effects of deficit irrigation with saline water on spring wheat growth and yield in arid Northwest China. J. Arid Land 2013, 5, 143–154. [Google Scholar] [CrossRef]
- Yu, X.R.; Li, B.; Wang, L.L.; Chen, X.Y.; Wang, W.J.; Gu, Y.J.; Wang, Z.; Xiong, F. Effect of drought stress on the development of endosperm starch granules and the composition and physicochemical properties of starches from soft and hard wheat. J. Sci. Food Agric. 2016, 96, 2746–2754. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, S.; Cheng, M.; Jiang, H.; Zhang, X.; Peng, C.; Lu, X.; Zhang, M.; Jin, J. Effect of drought on agronomic traits of rice and wheat: A meta-analysis. Int. J. Environ. Res. 2018, 15, 839. [Google Scholar] [CrossRef]
- Bapela, T.; Shimelis, H.; Tsilo, T.J.; Mathew, I. Genetic improvement of wheat for drought tolerance: Progress, challenges and opportunities. Plants 2022, 11, 1331. [Google Scholar] [CrossRef]
- Khadka, K.; Raizada, M.N.; Navabi, A. Recent progress in germplasm evaluation and gene mapping to enable breeding of drought-tolerant wheat. Front. Plant Sci. 2020, 11, 1149. [Google Scholar] [CrossRef]
- Baloch, M.J.; Dunwell, J.; Dennet, M.; Rajpar, I.; Jatoi, W.A.; Veesar, N.F. Evaluating spring wheat cultivars for drought tolerance through yield and physiological parameters at booting and anthesis. Afr. J. Biotechnol. 2012, 11, 11559–11565. [Google Scholar] [CrossRef]
- Ayalew, H.; Liu, H.; Börner, A.; Kobiljski, B.; Liu, C.; Yan, G. Genome-wide association mapping of major root length QTLs under PEG induced water stress in wheat. Front. Plant Sci. 2018, 9, 1759. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Change 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Llorens, E.; Sharon, O.; Camañes, G.; García-Agustín, P.; Sharon, A. Endophytes from wild cereals protect wheat plants from drought by alteration of physiological responses of the plants to water stress. Environ. Microbiol. 2019, 21, 3299–3312. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, M.; Germida, J.J.; Vujanovic, V. Fungal endophytes enhance wheat heat and drought tolerance in terms of grain yield and second-generation seed viability. J. Appl. Microbiol. 2014, 116, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Rezakhani, L.; Motesharezadeh, B.; Tehrani, M.M.; Etesami, H.; Mirseyed, H. Effect of silicon and phosphate-solubilizing bacteria on improved phosphorus (P) uptake is not specific to insoluble P-fertilized sorghum (Sorghum bicolor L.) plants. J. Plant Growth Regul. 2019, 39, 239–253. [Google Scholar] [CrossRef]
- Santos, M.; Ignacio, C.; Fernando, D.; Brenda, S.; Alejandro, M. Advances in the role of dark septate endophytes in the plant resistance to abiotic and biotic stresses. J. Fungi 2021, 7, 939. [Google Scholar] [CrossRef]
- Radzikowska, D.; Sulewska, H.; Bandurska, H.; Ratajczak, K.; Szymańska, G.; Kowalczewski, P.Ł.; Głowicka-Wołoszyn, R. Analysis of physiological status in response to water deficit of spelt (Triticum aestivum ssp. spelta) Cultivars in Reference to Common Wheat (Triticum aestivum ssp. vulgare). Agronomy 2022, 12, 1822. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, Y.; Zhang, S.; Wang, Y. What could promote farmers to replace chemical fertilizers with organic fertilizers? J. Clean. Prod. 2018, 199, 882–890. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A. Endophytic fungal communities and their biotechnological implications for agro-environmental sustainability. Folia Microbiol. 2022, 67, 203–232. [Google Scholar] [CrossRef]
- Rauf, M.; Awais, M.; Din, A.U.; Ali, K.; Gul, H.; Rahman, M.M.; Hamayun, M.; Arif, M. Molecular mechanisms of the 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase producing Trichoderma asperellum MAP1 in enhancing wheat tolerance to waterlogging stress. Front. Plant Sci. 2020, 11, 2213. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Zou, Y.N.; Kuca, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Elucidating the mechanisms underlying enhanced drought tolerance in plants mediated by arbuscular mycorrhizal fungi. Front. Microbiol. 2021, 12, 809473. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wu, Q.; Xu, L.; Druzhinina, I.S.; Stukenbrock, E.H.; Nieuwenhuis, B.P.S.; Zhong, Z.; Liu, Z.-J.; Wang, X.; Cai, F.; et al. Genomic landscape of a relict fir-associated fungus reveals rapid convergent adaptation towards endophytism. ISME J. 2022, 16, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Oberhofer, M.; Güsewell, S.; Leuchtmann, A. Effects of natural hybrid and non-hybrid Epichloë endophytes on the response of Hordelymus europaeus to drought stress. New Phytol. 2014, 201, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, Q.; Li, J.; Wang, Y.; Liu, X.Y.; Hu, Y.L.; Chen, J.Z. Contributions of an arbuscular mycorrhizal fungus to growth and physiology of loquat (Eriobotrya japonica) plants subjected to drought stress. Mycol. Prog. 2015, 14, 84. [Google Scholar] [CrossRef]
- Pedranzani, H.; Rodríguez-Rivera, M.; Gutiérrez, M.; Porcel, R.; Hause, B.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis regulates physiology and performance of Digitaria eriantha plants subjected to abiotic stresses by modulating antioxidant and jasmonate levels. Mycorrhiza 2016, 26, 141–152. [Google Scholar] [CrossRef]
- Jumpponen, A.; Trappe, J.M. Dark septate endophytes: A review of facultative biotrophic root-colonizing fungi. New Phytol. 1998, 140, 295–310. [Google Scholar] [CrossRef]
- Wagg, C.; Pautler, M.; Massicotte, H.B.; Peterson, R.L. The cooccurrence of ectomycorrhizal, arbuscular mycorrhizal, and dark septate fungi in seedlings of four members of the Pinaceae. Mycorrhiza 2008, 18, 103–110. [Google Scholar] [CrossRef]
- Mandyam, K.G.; Jumpponen, A. Mutualism-parasitism paradigm synthesized from results of root-endophyte models. Front. Microbiol. 2014, 5, 776. [Google Scholar]
- Ruotsalainen, A.L.; Kauppinen, M.; Wli, P.R.; Saikkonen, K.; Helander, M.; Tuomi, J. Dark septate endophytes, mutualism from by-products? Trends Plant Sci. 2022, 27, 247–254. [Google Scholar] [CrossRef]
- Newsham, K.K.; Upson, R.; Read, D.J. Mycorrhizas and dark septate root endophytes in polar regions. Fungal Ecol. 2009, 2, 10–20. [Google Scholar] [CrossRef]
- Zubek, S.; Nobis, M.; Błaszkowski, J.; Mleczko, P.; Nowak, A. Fungal root endophyte associations of plants endemic to the Pamir Alay Mountains of Central Asia. Symbiosis 2011, 54, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, D.G.; Németh, J.B.; Barry, K.; Hainaut, M.; Henrissat, B.; Johnson, J.; Kuo, A.; Lim, J.H.P.; Lipzen, A.; Nolan, M. Comparative genomics provides insights into the lifestyle and reveals functional heterogeneity of dark septate endophytic fungi. Sci. Rep. 2018, 8, 6321. [Google Scholar] [CrossRef] [PubMed]
- Knapp, D.G.; Imrefi, I.; Boldpurev, E.; Csíkos, S.; Akhmetova, G.; Berek-Nagy, P.J.; Otgonsuren, B.; Kovács, G.M. Root-colonizing endophytic fungi of the dominant grass Stipa krylovii from a Mongolian steppe grassland. Front. Microbiol. 2019, 10, 2565. [Google Scholar] [CrossRef] [PubMed]
- Lugo, M.A.; Menoyo, E.; Allione, L.R.; Negritto, M.A.; Henning, J.A.; Anton, A.M. Arbuscular mycorrhizas and dark septate endophytes associated with grasses from the Argentine Puna. Mycologia 2018, 110, 654–665. [Google Scholar] [CrossRef]
- Hou, L.; He, X.; Li, X.; Wang, S.; Zhao, L. Species composition and colonization of dark septate endophytes are affected by host plant species and soil depth in the Mu Us sandland, northwest China. Fungal Ecol. 2019, 39, 276–284. [Google Scholar] [CrossRef]
- Mateu, M.; Baldwin, A.; Maul, J.; Yarwood, S. Dark septate endophyte improves salt tolerance of native and invasive lineages of Phragmites australis. ISME J. 2020, 14, 1943–1954. [Google Scholar] [CrossRef]
- Zhu, L.; Li, T.; Wang, C.; Zhang, X.; Xu, L.; Xu, R.; Zhao, Z. The effects of dark septate endophyte (DSE) inoculation on tomato seedlings under Zn and Cd stress. Environ. Sci. Pollut. Res. 2018, 25, 35232–35241. [Google Scholar] [CrossRef]
- Li, M.; Hou, L.F.; Liu, J.Q.; Yang, J.Y.; Zou, Y.L.; Zhao, L.L.; He, X.L. Growth-promoting effects of dark septate endophytes on the non-mycorrhizal plant Isatis indigotica under different water conditions. Symbiosis 2021, 85, 291–303. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, T.; He, X. Mycorrhizal and dark septate endophytic fungi under the canopies of desert plants in Mu Us Sandy Land of China. Front. Agric. China 2009, 3, 164–170. [Google Scholar] [CrossRef]
- Vergara, C.; Araujo, K.E.C.; Urquiaga, S.; Santa-Catarina, C.; Schultz, N.; Araujo, E.D.S.; Balieiro, F.D.C.; Xavier, G.R.; Zilli, J.E. Dark septate endophytic fungi increase green manure-N-15 recovery efficiency, N contents, and micronutrients in rice grains. Front. Plant Sci. 2018, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, W.; Hou, J. Plant performance of enhancing licorice with dual inoculating dark septate endophytes and Trichoderma viride mediated via effects on root development. BMC Plant Biol. 2020, 20, 325. [Google Scholar] [CrossRef]
- Gaber, D.A.; Berthelot, C.; Camehl, I.; Kovács, G.M.; Blaudez, D.; Franken, P. Salt stress tolerance of dark septate endophytes is independent of melanin accumulation. Front. Microbiol. 2020, 11, 562931. [Google Scholar] [CrossRef] [PubMed]
- Gehring, C.; Sevanto, S.; Patterson, A.; Ulrich, D.E.M.; Kuske, C.R. Ectomycorrhizal and dark septate fungal associations of pinyon pine are differentially affected by experimental drought and warming. Front. Plant Sci. 2020, 11, 582574. [Google Scholar] [CrossRef] [PubMed]
- Spagnoletti, F.N.; Chiocchio, V.M. Tolerance of dark septate endophytic fungi (DSE) to agrochemicals in vitro. Rev. Argent Microbiol. 2020, 52, 43–49. [Google Scholar] [CrossRef]
- Akhtar, N.; Wani, A.K.; Dhanjal, D.S.; Mukherjee, S. Insights into the beneficial roles of dark septate endophytes in plants under challenging environment: Resilience to biotic and abiotic stresses. World J. Microb. Biot. 2022, 38, 79. [Google Scholar] [CrossRef]
- Iqbal, Z.; Iqbal, M.S.; Hashem, A.; Abd_Allah, E.F.; Ansari, M.I. Plant defense responses to biotic stress and its interplay with fluctuating dark/light. Front. Plant Sci. 2021, 12, 631810. [Google Scholar] [CrossRef]
- Su, Z.Z.; Dai, M.D.; Zhu, J.N.; Liu, X.H.; Li, L.; Zhu, X.M.; Wang, J.-Y.; Yuan, Z.-L.; Lin, F.-C. Dark septate endophyte Falciphora oryzae-assisted alleviation of cadmium in rice. J. Hazard Mater. 2021, 419, 126435. [Google Scholar] [CrossRef]
- Potisek, M.; Likar, M.; Vogel-Mikuš, K.; Arcon, I.; Grdadolnik, J.; Regvar, M. 1,8-dihydroxy naphthalene (DHN)-melanin confers tolerance to cadmium in isolates of melanised dark septate endophytes. Ecotoxicol Environ. Saf. 2021, 222, 112493. [Google Scholar] [CrossRef]
- Li, X.; He, X.; Hou, L.; Ren, Y.; Wang, S.; Su, F. Dark septate endophytes isolated from a xerophyte plant promote the growth of Ammopiptanthus mongolicus under drought condition. Sci. Rep. 2018, 8, 7896. [Google Scholar] [CrossRef]
- Xie, L.; He, X.; Wang, K.; Hou, L.; Sun, Q. Spatial dynamics of dark septate endophytes in the roots and rhizospheres of Hedysarum scoparium in northwest China and the influence of edaphic variables. Fungal Ecol. 2017, 26, 135–143. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Herrera, J.; Sinsabaugh, R.L.; Odenbach, K.J.; Lowrey, T.; Natvig, D.O. Novel root fungal consortium associated with a dominant desert grass. Appl. Environ. Microb. 2008, 74, 2805–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Su, F.; He, X.; Li, M. Colonization by dark septate endophytes improves the growth of Hedysarum scoparium under multiple inoculum levels. Symbiosis 2020, 82, 201–214. [Google Scholar] [CrossRef]
- Santos, S.G.; Silva, P.R.; Garcia, A.C.; Zilli, J.É.; Berbara, R.L. Dark septate endophyte decreases stress on rice plants. Braz. J. Microbiol. 2017, 48, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, M.J.; Zhang, X.T.; Zhang, H.B.; Sha, T.; Zhao, Z.W. Improved tolerance of maize (Zea mays L.) to heavy metals by colonization of a dark septate endophyte (DSE) Exophiala pisciphila. Sci. Total Environ. 2011, 409, 1069–1074. [Google Scholar] [CrossRef]
- Li, X.; He, X.L.; Zhou, Y.; Hou, Y.T.; Zuo, Y.L. Effects of dark septate endophytes on the performance of Hedysarum scoparium under water deficit stress. Front. Plant Sci. 2019, 10, 903. [Google Scholar] [CrossRef]
- Farias, G.C.; Nunes, K.G.; Soares, M.A.; de Siqueira, K.A.; Lima, W.C.; Neves, A.L.R.; de Lacerda, C.F.; Filho, E.G. Dark septate endophytic fungi mitigate the effects of salt stress on cowpea plants. Braz. J. Microbiol. 2020, 51, 243–253. [Google Scholar] [CrossRef]
- Li, Y.P.; He, X.L.; Zhao, L.L. Tempo-spatial dynamics of arbuscular mycorrhizal fungi under clonal plant Psammochloa villosa Trin. Bor in Mu Us sandland. Eur. J. Soil Biol. 2010, 46, 295–301. [Google Scholar] [CrossRef]
- Liu, Y.; Lv, T.; Su, X.; Ren, Z. Complete chloroplast genome of Psammochloa villosa (Poaceae), a pioneer grass endemic to sand dunes in Northwest China. Cytol. Genet. 2020, 54, 582–587. [Google Scholar] [CrossRef]
- Hou, L. Species Diversity and Salt Tolerance of Dark Septate Endophytes in Three Desert Plants. Ph.D. Dissertation, Hebei University, Baoding, China, 2020. [Google Scholar]
- Zuo, Y.; Hu, Q.; Liu, J.; He, X.L. Relationship of root dark septate endophytes and soil factors to plant species and seasonal variation in extremely arid desert in Northwest China. Appl. Soil Ecol. 2022, 175, 104454. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Xu, M.; Ye, Q.; Gao, H.; He, X. Improved tolerance of Artemisia ordosica to drought stress via dark septate endophyte (DSE) symbiosis. J. Fungi 2022, 8, 730. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, X. Dark septate endophyte improves the drought-stress resistance of Ormosia hosiei seedlings by altering leaf morphology and photosynthetic characteristics. Plant Ecol. 2021, 222, 761–771. [Google Scholar] [CrossRef]
- Li, X.; He, C.; He, X.; Su, F.; Hou, L.; Ren, Y.; Hou, Y. Dark septate endophytes improve the growth of host and non-host plants under drought stress through altered root development. Plant Soil 2019, 439, 259–272. [Google Scholar] [CrossRef]
- Zuo, Y.; Hu, Q.; Zhang, K.; He, X. Host and tissue affiliations of culturable endophytic fungi associated with xerophytic plants in the desert region of Northwest China. Agronomy 2022, 12, 727. [Google Scholar] [CrossRef]
- Elavarthi, S.; Martin, B. Spectrophotometric assays for antioxidant enzymes in plants. Methods Mol. Biol. 2010, 639, 273–280. [Google Scholar]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Method Enzymol. 1985, 113, 548–555. [Google Scholar]
- Peever, T.L.; Higgins, V.J. Electrolyte leakage, lipoxygenase, and lipid peroxidation induced in tomato leaf tissue by specific and nonspecific elicitors from Cladosporium fulvum. Plant Physiol. 1989, 90, 867–875. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Christos, D.G.; Konstantinos, G.; George, Z. Mechanism of Coomassie brilliant blue G-250 binding to proteins: A hydrophobic assay for nanogram quantities of proteins. Anal. Bioanal. Chem. 2008, 3912, 391–403. [Google Scholar]
- Hou, L.; Li, X.; He, X.; Zuo, Y.; Zhang, D.; Zhao, L. Effect of dark septate endophytes on plant performance of Artemisia ordosica and associated soil microbial functional group abundance under salt stress. Appl. Soil Ecol. 2021, 165, 103998. [Google Scholar] [CrossRef]
- Khaksar, G.; Treesubsuntorn, C.; Thiravetyan, P. Effect of endophytic Bacillus cereus ERBP inoculation into non-native host: Potentials and challenges for airborne formaldehyde removal. Plant Physiol. Biochem. 2016, 107, 326–336. [Google Scholar] [CrossRef]
- Irizarry, I.; White, J.F. Application of bacteria from non-cultivated plants to promote growth, alter root architecture and alleviate salt stress of cotton. J. Appl. Microbiol. 2017, 122, 1110–1120. [Google Scholar] [CrossRef]
- Ban, Y.; Xu, Z.; Yang, Y.; Zhang, H.; Chen, H.; Tang, M. Effect of dark septate endophytic fungus Gaeumannomyces cylindrosporus on plant growth, photosynthesis and pb tolerance of maize (Zea mays L.). Pedosphere 2017, 27, 283–292. [Google Scholar] [CrossRef]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowakhuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Song, X.; Halifu, S.; Yu, W.; Song, R. Effects of dark septate endophytes strain A024 on damping-off biocontrol, plant growth and the rhizosphere soil environment of Pinus sylvestris var. mongolica annual seedlings. Plants 2020, 9, 913. [Google Scholar]
- Arunyanark, A.; Jogloy, S.; Akkasaeng, C.; Vorasoot, N.; Patanothai, A. Chlorophyll stability is an indicator of drought tolerance in peanut. J. Agron. Crop Sci. 2010, 194, 113–125. [Google Scholar] [CrossRef]
- Ghobadi, M.; Taherabadi, S.; Ghobadi, M.E.; Mohammadi, G.R.; Jalali-Honarmand, S. ntioxidant capacity, photosynthetic characteristics and water relations of sunflower (Helianthus annuus L.) cultivars in response to drought stress. Ind. Crops Prod. 2013, 50, 29–38. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Zhang, K.; Tian, C.; Guo, J. Arbuscular mycorrhizal fungi improve plant growth of Ricinus communis by altering photosynthetic properties and increasing pigments under drought and salt stress. Ind Crops Prod. 2018, 117, 13–19. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Lubna, A.S.; Hamayun, M.; Khan, A.L.; Waqas, M.; Khan, M.A.; Jan, R.; In-Jung, L.; Hussain, A. Salt tolerance of Glycine max. L induced by endophytic fungus Aspergillus flavus CSH1, via regulating its endogenous hormones and antioxidative system. Plant Physiol. Biochem. 2018, 128, 13–23. [Google Scholar] [CrossRef]
- Santner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between plant hormones and heavy metals responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, L.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef]
- Xu, L.; Wu, C.; Oelmueller, R.; Zhang, W. Role of phytohormones in Piriformospora indica-induced growth promotion and stress tolerance in plants: More questions than answers. Front. Microbiol. 2018, 9, 1646. [Google Scholar] [CrossRef]
- Khan, A.R.; Ullah, I.; Waqas, M.; Shahzad, R.; Hong, S.; Park, G.; Jung, B.K.; Lee, I.; Shin, J. Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World J. Microbiol. Biotechnol. 2015, 31, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Impa, S.M.; Nadaradjan, S.; Jagadish, S.V.K. Drought stress induced reactive oxygen species and anti-oxidants in plants. In Abiotic Stress Responses in Plants; Springer: New York, NY, USA, 2012; pp. 131–147. [Google Scholar]
- Dastogeer, K.M.G. Influence of fungal endophytes on plant physiology is more pronounced under stress than well-watered conditions: A meta-analysis. Planta 2018, 248, 1403–1416. [Google Scholar] [CrossRef] [PubMed]
- Guler, N.S.; Pehlivan, N.; Karaoglu, S.A.; Guzel, S.; Bozdeveci, A. Trichoderma atroviride ID20G inoculation ameliorates drought stress-induced damages by improving antioxidant defense in maize seedlings. Acta Physiol. Plant 2016, 38, 132. [Google Scholar] [CrossRef]
- Tyagi, J.; Varma, A.; Pudake, R.N. Evaluation of comparative effects of arbuscular mycorrhiza (Rhizophagus intraradices) and endophyte (Piriformospora indica) association with finger millet (Eleusine coracana) under drought stress. Eur. J. Soil Biol. 2017, 81, 1–10. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Khan, A.L.; Kang, S.M.; Lee, I. A comparative study of phosphate solubilization and the host plant growth promotion ability of Fusarium verticillioides RK01 and Humicola sp. KNU01 under salt stress. Ann. Microbiol. 2015, 65, 585–593. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Tian, Y.; Xue, Y.; Xu, X.; Sui, Y.; Yu, H. Oxidative stress and potential biomarkers in tomato seedlings subjected to soil lead contamination. Ecotox. Environ. Safe 2008, 71, 685–691. [Google Scholar] [CrossRef]
- He, C.; Wang, W.; Hou, J. Plant growth and soil microbial impacts of enhancing licorice with inoculating dark septate endophytes under drought stress. Front. Microbiol. 2019, 10, 2277. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wang, Y.; Xia, D.; Gao, C.; Chao, C. Overexpression of a GST gene (ThGSTZ1) from Tamarix hispida improves drought and salinity tolerance by enhancing the ability to scavenge reactive oxygen species. Plant Cell Tissue Org. 2014, 117, 99–112. [Google Scholar] [CrossRef]
- Sadeghi, F.; Samsampour, D.; Seyahooei, M.A.; Bagheri, A.; Soltani, J. Fungal endophytes alleviate drought-induced oxidative stress in mandarin (Citrus reticulata L.): Toward regulating the ascorbate-glutathione cycle. Sci. Hortic. 2020, 261, 108991. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Chen, L.; Liu, L.; Lu, B.; Ma, T.; Li, C. Exogenous melatonin promotes seed germination and osmotic regulation under salt stress in cotton (Gossypium hirsutum L.). PLoS ONE 2020, 15, e0228241. [Google Scholar]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial diversity of upland rice roots and their influence on rice growth and drought tolerance. Microorganisms 2020, 8, 1329. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Bio/Technol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Alibolandi, M.; Mirzahoseini, H. Chemical assistance in refolding of bacterial inclusion bodies. Biochem. Res. Int. 2011, 2011, 631607. [Google Scholar]
- Moghaddam, M.S.H.; Safaie, N.; Soltani, J.; Hagh-Doust, N. Desert-adapted fungal endophytes induce salinity and drought stress resistance in model crops. Plant Physiol. Biochem. 2021, 160, 225–238. [Google Scholar] [CrossRef]
Figure 1.
Typical DSE hyphae and microsclerotia in the inoculated seedlings. H, hyphae; M, microsclerotia. (a–e) DSE hyphae in the inoculated wheat roots. (f–j) DSE microsclerotia in the inoculated wheat roots.
Figure 1.
Typical DSE hyphae and microsclerotia in the inoculated seedlings. H, hyphae; M, microsclerotia. (a–e) DSE hyphae in the inoculated wheat roots. (f–j) DSE microsclerotia in the inoculated wheat roots.
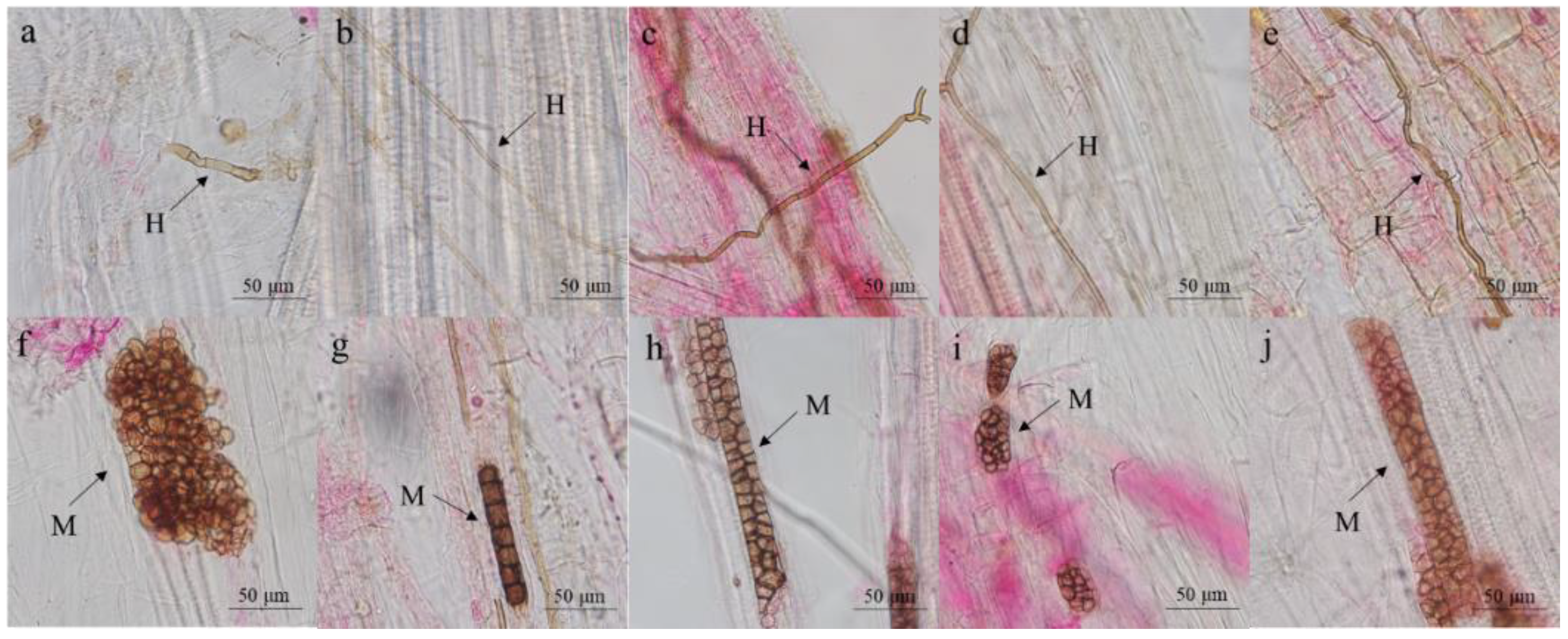
Figure 2.
Influences of DSE on the growth parameters of wheat seedlings grown in different water conditions. (a) leaf number; (b) plant height; (c) shoot and root biomass; (d) total biomass; (e) auxin content. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups (p < 0.05). WW, well-watered. MD, mild drought stress. ED, severe drought stress.
Figure 2.
Influences of DSE on the growth parameters of wheat seedlings grown in different water conditions. (a) leaf number; (b) plant height; (c) shoot and root biomass; (d) total biomass; (e) auxin content. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups (p < 0.05). WW, well-watered. MD, mild drought stress. ED, severe drought stress.
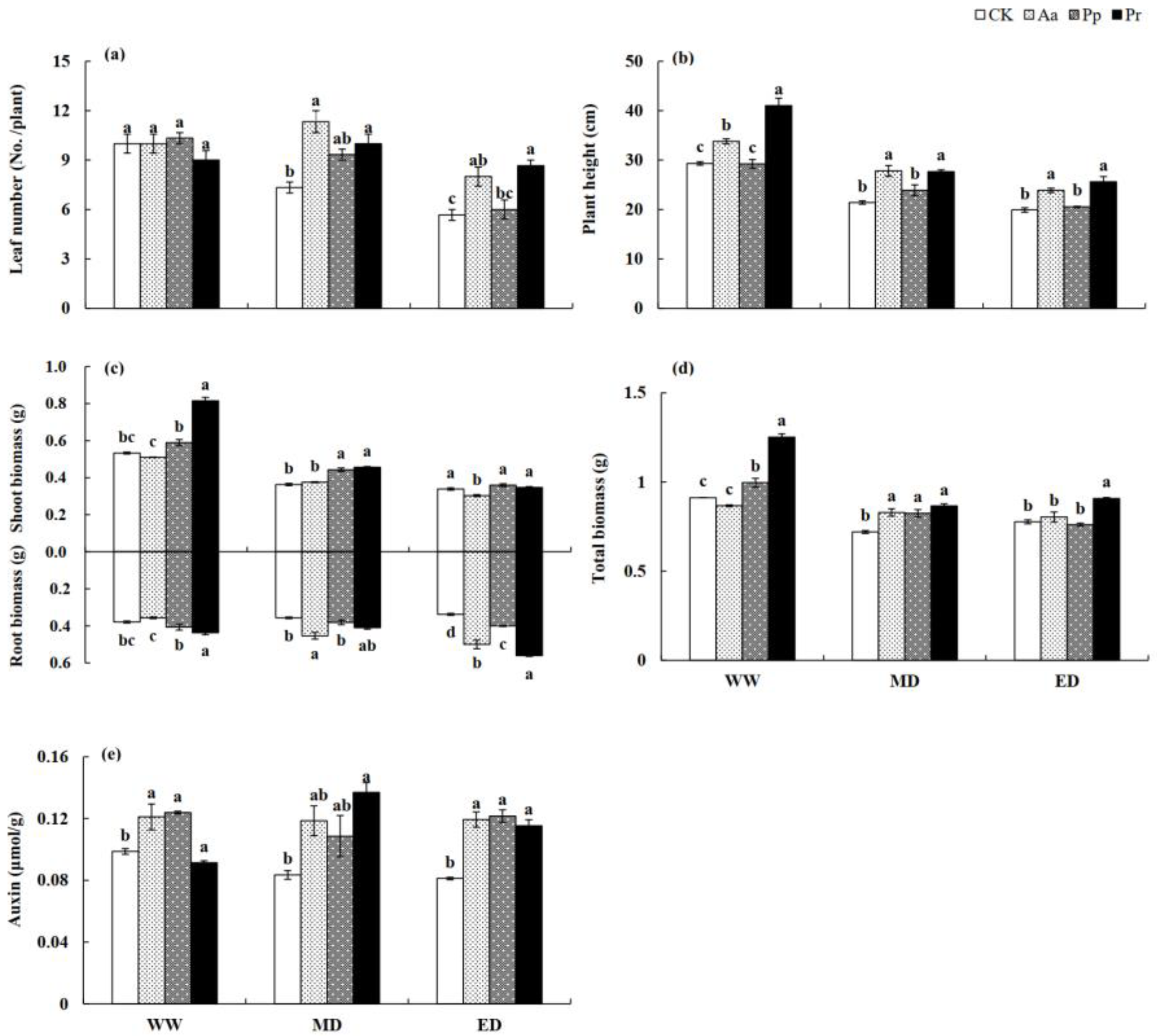
Figure 3.
Influences of DSE on the antioxidant enzyme system of wheat leaves under three water treatments. (a) GSH, glutathione content. (b) SOD, superoxide dismutase. (c) MDA, malondialdehyde content. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups (p < 0.05). WW, well-watered. MD, mild drought stress. ED, severe drought stress.
Figure 3.
Influences of DSE on the antioxidant enzyme system of wheat leaves under three water treatments. (a) GSH, glutathione content. (b) SOD, superoxide dismutase. (c) MDA, malondialdehyde content. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups (p < 0.05). WW, well-watered. MD, mild drought stress. ED, severe drought stress.

Figure 4.
Influences of DSE on the contents of soluble protein and sugar and proline in wheat leaves under different water treatments. (a) soluble protein; (b) soluble sugar; (c) proline. WW, well-watered. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups (p < 0.05). MD, mild drought stress. ED, severe drought stress.
Figure 4.
Influences of DSE on the contents of soluble protein and sugar and proline in wheat leaves under different water treatments. (a) soluble protein; (b) soluble sugar; (c) proline. WW, well-watered. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups (p < 0.05). MD, mild drought stress. ED, severe drought stress.

Figure 5.
The influences of the DSE treatment on the performance of wheat under three levels of water treatment based on principal component analysis. (a–c) represent principal component analysis of the DSE’s influence on the fitness of wheat seedlings under WW (a), MD (b), and ED (c), respectively. Leaf, leaf number. Height, plant height. GSH, glutathione content. SOD, superoxide dismutase. MDA, malondialdehyde content. SS, soluble sugar content. SP, content of soluble protein. Pro, proline contents. Gs, stomatal conductance. Pn, photosynthetic rate. Ci, intercellular CO2 concentration. Tr, transpiration rate. SDW, shoot biomass. RDW, root biomass. TDW, total biomass. ○, ◇, ⟡, and △ indicate CK, Aa, Pp, and Pr, respectively.
Figure 5.
The influences of the DSE treatment on the performance of wheat under three levels of water treatment based on principal component analysis. (a–c) represent principal component analysis of the DSE’s influence on the fitness of wheat seedlings under WW (a), MD (b), and ED (c), respectively. Leaf, leaf number. Height, plant height. GSH, glutathione content. SOD, superoxide dismutase. MDA, malondialdehyde content. SS, soluble sugar content. SP, content of soluble protein. Pro, proline contents. Gs, stomatal conductance. Pn, photosynthetic rate. Ci, intercellular CO2 concentration. Tr, transpiration rate. SDW, shoot biomass. RDW, root biomass. TDW, total biomass. ○, ◇, ⟡, and △ indicate CK, Aa, Pp, and Pr, respectively.

Figure 6.
Influence of DSE on the leaf photosynthetic indexes and chlorophyll content under different water treatments. (a) Pn, leaf photosynthetic rate; (b) Gs, leaf stomatal conductance; (c) Tr, leaf transpiration rate; (d) Ci, leaf intercellular CO2 concentration; (e) Chlorophyll content. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups. WW, well-watered. MD, mild drought stress. ED, severe drought stress.
Figure 6.
Influence of DSE on the leaf photosynthetic indexes and chlorophyll content under different water treatments. (a) Pn, leaf photosynthetic rate; (b) Gs, leaf stomatal conductance; (c) Tr, leaf transpiration rate; (d) Ci, leaf intercellular CO2 concentration; (e) Chlorophyll content. Different letters under a certain water condition indicate significant differences among the four DSE treatment groups. WW, well-watered. MD, mild drought stress. ED, severe drought stress.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The influences of DSE (DSE) and water (Water) treatments on the growth parameters of wheat seedlings based on two-way analysis of variance (ANOVA).
Table 1.
The influences of DSE (DSE) and water (Water) treatments on the growth parameters of wheat seedlings based on two-way analysis of variance (ANOVA).
| Leaf Number | Plant Height | Shoot Biomass | Root Biomass | Total Biomass | Auxin Content | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | |
| DSE | 9.91 | <0.001 | 62.04 | 0.000 | 159.50 | <0.001 | 53.19 | <0.001 | 98.98 | <0.001 | 17.76 | <0.001 |
| Water | 36.04 | <0.001 | 199.08 | 0.000 | 1033.79 | <0.001 | 27.76 | <0.001 | 202.50 | <0.001 | 0.30 | 0.747 |
| DSE × Water | 5.62 | 0.001 | 6.16 | 0.001 | 62.01 | <0.001 | 22.20 | <0.001 | 25.70 | <0.001 | 5.89 | 0.001 |
Note: Significant p values (p < 0.05) are shown in bold.
Table 2.
The influences of DSE (DSE) and water (Water) treatments on the physiological indexes of wheat leaves based on two-way analysis of variance (ANOVA).
Table 2.
The influences of DSE (DSE) and water (Water) treatments on the physiological indexes of wheat leaves based on two-way analysis of variance (ANOVA).
| GSH | SOD | MDA | Soluble Protein | Soluble Sugar | Proline | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | |
| DSE | 5.62 | 0.005 | 55.76 | <0.001 | 16.78 | 0.000 | 16.85 | <0.001 | 109.74 | <0.001 | 107.81 | <0.001 |
| Water | 376.29 | <0.001 | 265.38 | <0.001 | 7.84 | 0.002 | 21.41 | <0.001 | 320.68 | <0.001 | 299.35 | <0.001 |
| DSE × Water | 7.68 | <0.001 | 23.11 | <0.001 | 15.36 | 0.000 | 3.92 | 0.007 | 25.07 | <0.001 | 19.32 | <0.001 |
Note: Significant p values (p < 0.05) are shown in bold. GSH, glutathione content. SOD, superoxide dismutase. MDA, malondialdehyde content.
Table 3.
The influences of DSE (DSE) and water (Water) treatments on the photosynthetic indexes and chlorophyll content of wheat leaves based on two-way analysis of variance (ANOVA).
Table 3.
The influences of DSE (DSE) and water (Water) treatments on the photosynthetic indexes and chlorophyll content of wheat leaves based on two-way analysis of variance (ANOVA).
| Photosynthetic Rate | Stomatal Conductance | Transpiration Rate | Intercellular CO2 Concentration | Chlorophyll | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | |
| DSE | 24.52 | <0.001 | 14.50 | <0.001 | 14.43 | <0.001 | 19.54 | <0.001 | 33.53 | <0.001 |
| Water | 167.07 | <0.001 | 856.99 | <0.001 | 406.11 | <0.001 | 57.48 | <0.001 | 14.88 | <0.001 |
| DSE × Water | 19.11 | <0.001 | 8.57 | <0.001 | 5.34 | 0.001 | 5.74 | 0.001 | 8.80 | <0.001 |
Note: Significant p values (p < 0.05) are shown in bold. Pn, leaf photosynthetic rate. Gs, leaf stomatal conductance. Tr, leaf transpiration rate. Ci, leaf intercellular CO2 concentration.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, X.; Liu, Y.; Ye, Q.; Xu, M.; He, X. Application of Desert DSEs to Nonhost Plants: Potential to Promote Growth and Alleviate Drought Stress of Wheat Seedlings. Agriculture 2022, 12, 1539. https://doi.org/10.3390/agriculture12101539
AMA Style
Li X, Liu Y, Ye Q, Xu M, He X. Application of Desert DSEs to Nonhost Plants: Potential to Promote Growth and Alleviate Drought Stress of Wheat Seedlings. Agriculture. 2022; 12(10):1539. https://doi.org/10.3390/agriculture12101539
Chicago/Turabian StyleLi, Xia, Yanxia Liu, Qiannan Ye, Minghui Xu, and Xueli He. 2022. "Application of Desert DSEs to Nonhost Plants: Potential to Promote Growth and Alleviate Drought Stress of Wheat Seedlings" Agriculture 12, no. 10: 1539. https://doi.org/10.3390/agriculture12101539
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.