
Improved Satellite Cell Proliferation Induced by L-Carnosine Benefits Muscle Growth of Pigs in Part through Activation of the Akt/mTOR/S6K Signaling Pathway

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Handling and Feeding Pigs
2.2. Primary pig Skeletal Muscle SCs Culture
2.3. Proliferation Activity Analyses
2.3.1. MTT Assay
2.3.2. Crystal Violet Assay
2.3.3. Flow Cytometry
2.4. Western Blot
2.5. Statistical Analyses
3. Results
3.1. Effect of L-Carnosine on Growth Performance of Finishing Pigs
3.2. Effect of L-Carnosine on the mTOR Signaling Pathway in the Longissimus Dorsi Muscle of Finishing Pigs
3.3. Effect of L-Carnosine on Satellite Cell Proliferation
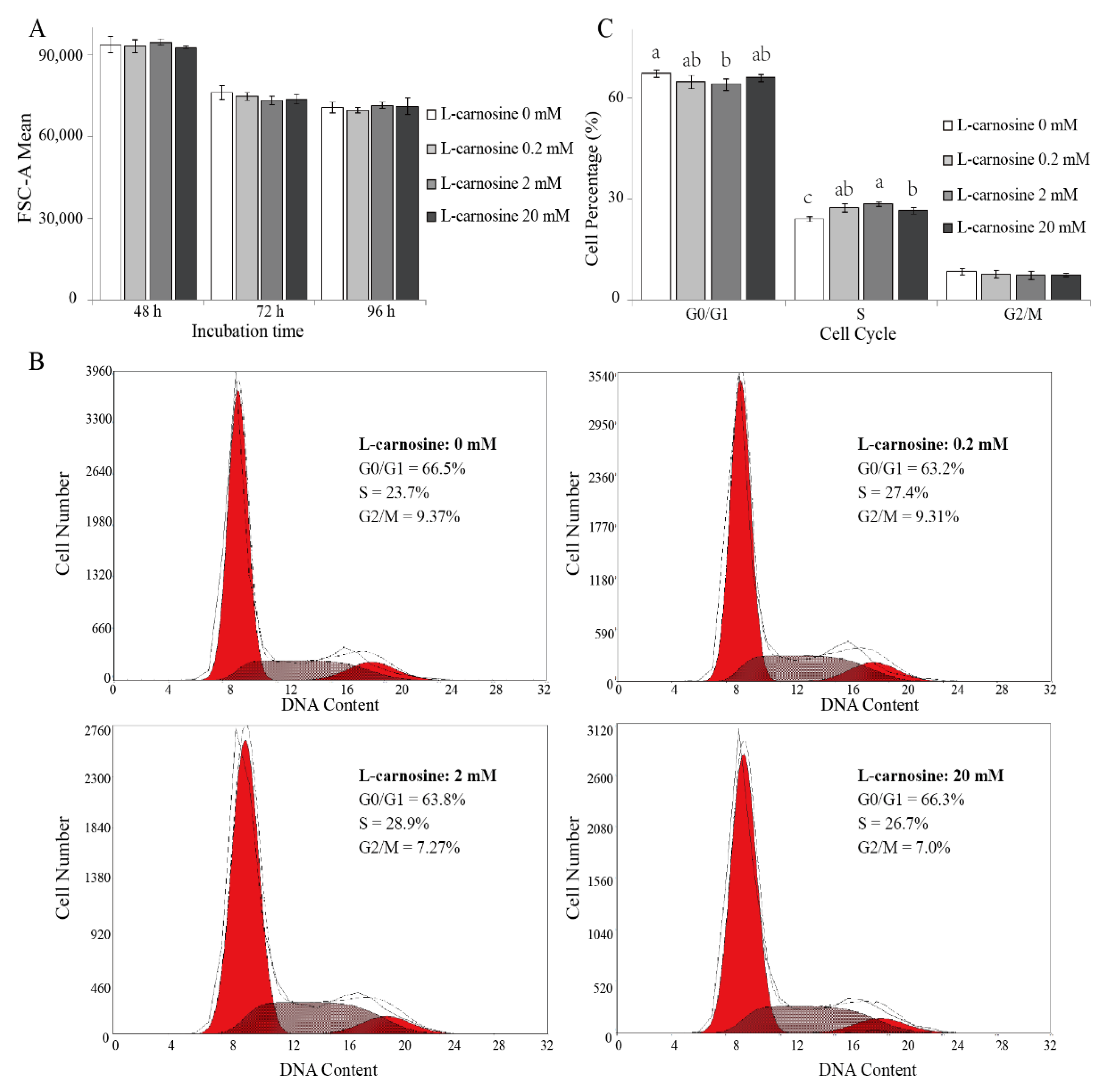
3.4. Effect of L-Carnosine on Cell Size and Cell Cycle of Satellite Cells
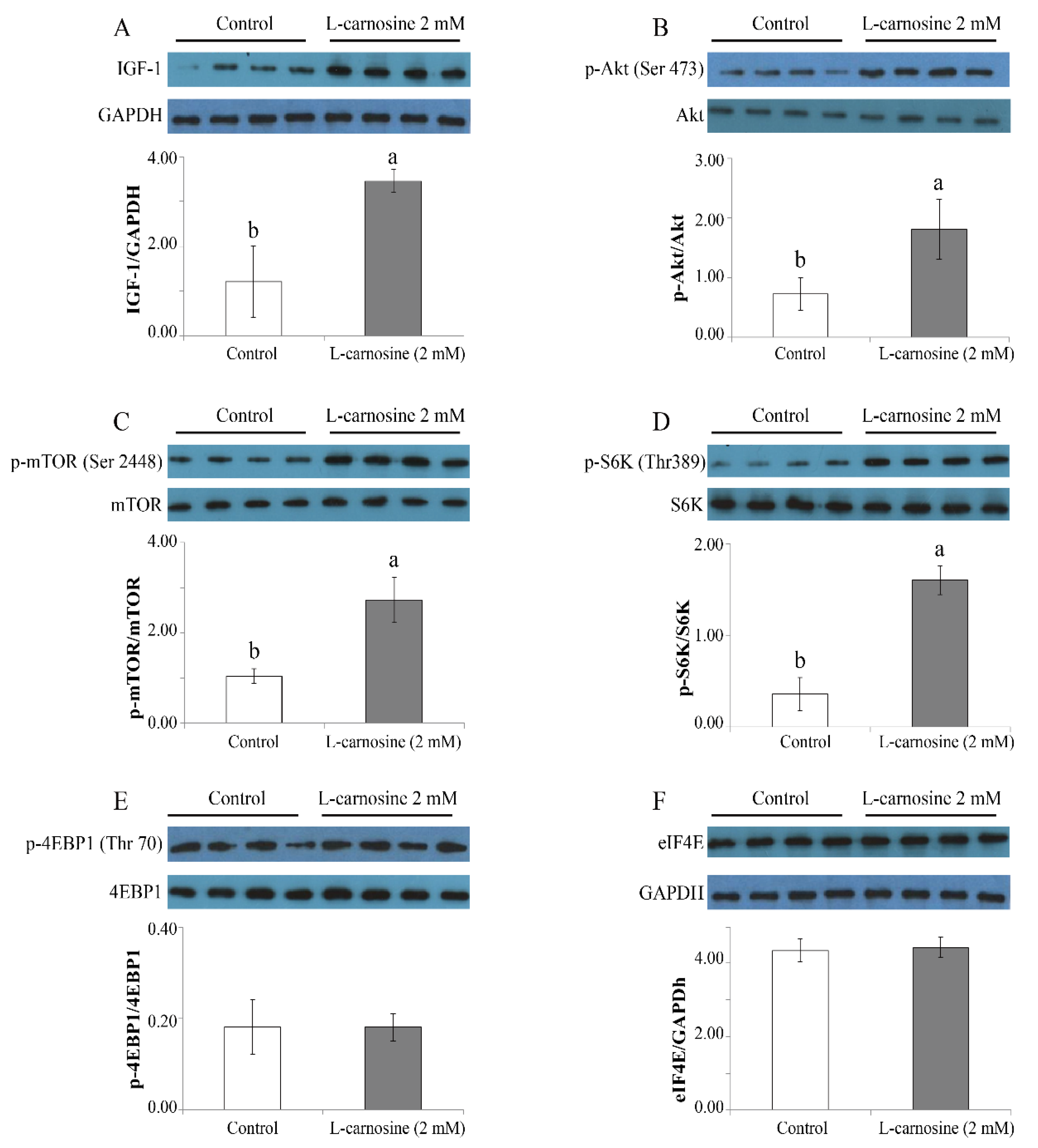
3.5. L-carnosine Activated IGF-1/PI3K/S6K Signaling Pathway in Cultured Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Li, X.; Yang, X.; Sun, Q.; Huang, R.; Xing, J.; Zhao, R. Maternal dietary protein induces opposite myofiber type transition in Meishan pigs at weaning and finishing stages. Meat Sci. 2011, 89, 221–227. [Google Scholar] [CrossRef]
- Rowlerson, A.; Veggetti, A. Cellular mechanisms of post-embryonic muscle growth in aquaculture species. Fish Physiol. 2001, 18, 103–140. [Google Scholar]
- Berard, J.; Kalbe, C.; Losel, D.; Tuchscherer, A.; Rehfeldt, C. Potential sources of early-postnatal increase in myofibre number in pig skeletal muscle. Histochem. Cell Biol. 2011, 136, 217–225. [Google Scholar] [CrossRef]
- Qi, R.; Chen, Y.; Peng, H.; Jiang, S.; Xiao, R.; Huang, J. Conjugated linoleic acid supplementation during late gestation and lactation of sows affects rnyofiber type in their litters. Livest. Sci. 2015, 178, 322–329. [Google Scholar] [CrossRef]
- Chang, N.C.; Chevalier, F.P.; Rudnicki, M.A. Satellite cells in muscular dystrophy-lot in polarity. Trends Mol. Med. 2016, 22, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Yajima, H.; Motohashi, N.; Ono, Y.; Sato, S.; Ikeda, K.; Masuda, S.; Yada, E.; Kanesaki, H.; Miyagoe-Suzuki, Y.; Takeda, S.I.; et al. Six family genes control the proliferation and differentiation of muscle satellite cells. Exp. Cell Res. 2010, 316, 2932–2944. [Google Scholar] [CrossRef]
- Hawke, T.J.; Garry, D.J. Myogenic satellite cells: Physiology to molecular biology. J. Appl. Physiol. 2001, 91, 534–551. [Google Scholar]
- Tierney, M.T.; Sacco, A. Satellite Cell Heterogeneity in Skeletal Muscle Homeostasis. Trends Cell Biol. 2016, 26, 434–444. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.Q.; Zhao, Y.L.; Li, H.C.; Sui, W.G.; Yan, H.C.; Wang, X.Q. Heat stress inhibits proliferation, promotes growth, and induces apoptosis in cultured Lantang swine skeletal muscle satellite cells. J. Zhejiang Univ. Sci. B 2015, 16, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, K.; Nagata, Y.; Wada, E.; Zammit, P.S.; Shiozuka, M.; Matsuda, R. Zinc promotes proliferation and activation of myogenic cells via the PI3K/Akt and ERK signaling cascade. Exp. Cell Res. 2015, 333, 228–237. [Google Scholar] [CrossRef]
- Torres, A.S.; Holz, M.K. Unraveling the multifaceted nature of the nuclear function of mTOR. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118907. [Google Scholar] [CrossRef]
- Goodman, C.A. The role of mTORC1 in regulating protein synthesis and skeletal muscle mass in response to various mechanical stimuli. Rev. Physiol. Biochem. Pharmacol. 2014, 166, 43–95. [Google Scholar] [CrossRef]
- Pallafacchina, G.; Blaauw, B.; Schiaffino, S. Role of satellite cells in muscle growth and maintenance of muscle mass. Nutr. Metab. Cardiovasc. Dis. 2013, 23 (Suppl. S1), S12–S18. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Abbate, J.M.; Macri, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of Protein Hydrolysates from Anchovy (Engraulis Encrasicolus) Waste for Twelve Weeks Decreases Metabolic Dysfunction-Associated Fatty Liver Disease Severity in ApoE(−/−)Mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef]
- Avondo, M.; Pagano, R.I.; Guastella, A.M.; Criscione, A.; Di Gloria, M.; Valenti, B.; Piccione, G.; Pennisi, P. Diet selection and milk production and composition in Girgentana goats with different alpha s1-casein genotype. J. Dairy Res. 2009, 76, 202–209. [Google Scholar] [CrossRef]
- Vazzana, I.; Rizzo, M.; Giambelluca, S.; Zumbo, A.; Piccione, G.; Monteverde, V. The response of some blood constituents after administration of two different diets in goats. Comp. Clin. Pathol. 2013, 23, 1587–1591. [Google Scholar] [CrossRef]
- Agrawal, A.; Rathor, R.; Kumar, R.; Singh, S.N.; Kumar, B.; Suryakumar, G. Endogenous dipeptide-carnosine supplementation ameliorates hypobaric hypoxia-induced skeletal muscle loss via attenuating endoplasmic reticulum stress response and maintaining proteostasis. IUBMB Life 2022, 74, 101–116. [Google Scholar] [CrossRef]
- Cong, J.; Zhang, L.; Li, J.; Wang, S.; Gao, F.; Zhou, G. Effects of dietary supplementation with carnosine on growth performance, meat quality, antioxidant capacity and muscle fiber characteristics in broiler chickens. J. Sci. Food Agric. 2017, 97, 3733–3741. [Google Scholar] [CrossRef]
- Bao, Y.; Gao, C.; Hao, W.; Ji, C.; Zhao, L.; Zhang, J.; Liu, T.; Ma, Q. Effects of Dietary L-carnosine and Alpha-lipoic Acid on Growth Performance, Blood Thyroid Hormones and Lipid Profiles in Finishing Pigs. Asian-Australas. J. Anim. Sci. 2015, 28, 1465–1470. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.X.; Hongtrakul, K.; Ji, C.; Ma, Q.G.; Guan, S.; Song, C.L.; Zhang, Y.; Zhao, L.H. Effect of Carnosine on Growth Performance, Carcass Characteristics, Meat Quality and Oxidative Stability in Broiler Chickens. J. Poult. Sci. 2009, 46, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Kadooka, K.; Fujii, K.; Matsumoto, T.; Sato, M.; Morimatsu, F.; Tashiro, K.; Kuhara, S.; Katakura, Y. Mechanisms and consequences of carnosine-induced activation of intestinal epithelial cells. J. Funct. Foods 2015, 13, 32–37. [Google Scholar] [CrossRef]
- Stuerenburg, H.J.; Kunze, K. Concentrations of free carnosine (a putative membrane-protective antioxidant) in human muscle biopsies and rat muscles. Arch. Gerontol. Geriatr. 1999, 29, 107–113. [Google Scholar] [CrossRef]
- Limonta, E.; Invernizzi, P.L.; Rampichini, S.; Bisconti, A.V.; Ce, E.; Longo, S.; Coratella, G.; Iaia, F.M.; Esposito, F. Acute carnosine and beta-alanine supplementation increase the compensated part of the ventilation versus work rate relationship during a ramp incremental cycle test in physically active men. J. Sports Med. Phys. Fit. 2021, 61, 37–43. [Google Scholar] [CrossRef]
- Wang, X.Q.; Yang, W.J.; Yang, Z.; Shu, G.; Wang, S.B.; Jiang, Q.Y.; Yuan, L.; Wu, T.S. The differential proliferative ability of satellite cells in Lantang and Landrace pigs. PLoS ONE 2012, 7, e32537. [Google Scholar] [CrossRef] [Green Version]
- Slack-Davis, J.K.; Martin, K.H.; Tilghman, R.W.; Iwanicki, M.; Ung, E.J.; Autry, C.; Luzzio, M.J.; Cooper, B.; Kath, J.C.; Roberts, W.G.; et al. Cellular characterization of a novel focal adhesion kinase inhibitor. J. Biol. Chem. 2007, 282, 14845–14852. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.H.; Song, J.L.; Delafontaine, P.; Godard, M.P. The therapeutic potential of IGF-I in skeletal muscle repair. Trends Endocrinol. Metab. 2013, 24, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, K.; Berneman, Z.N.; Van Bockstaele, D.R. Cell cycle and apoptosis. Cell Prolif. 2003, 36, 165–175. [Google Scholar] [CrossRef]
- Jewell, J.L.; Kim, Y.C.; Russell, R.C.; Yu, F.X.; Park, H.W.; Plouffe, S.W.; Tagliabracci, V.S.; Guan, K.L. Metabolism. Differential regulation of mTORC1 by leucine and glutamine. Science 2015, 347, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Wolfson, R.L.; Chantranupong, L.; Saxton, R.A.; Shen, K.; Scaria, S.M.; Cantor, J.R.; Sabatini, D.M. Sestrin2 is a leucine sensor for the mTORC1 pathway. Science 2016, 351, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.L.; Ye, J.L.; Yang, J.; Gao, C.Q.; Yan, H.C.; Li, H.C.; Wang, X.Q. mTORC1 mediates lysine-induced satellite cell activation to promote skeletal muscle growth. Cells 2019, 8, 1549. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Zhang, T.; Cao, Y.; Deng, B.; Zhang, J.; Zhao, J. Effects of dietary sea buckthorn pomace supplementation on skeletal muscle mass and meat quality in lambs. Meat Sci. 2020, 166, 108141. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R.; Zlotchenko, E.; Scrimgeour, A.; Lawrence, J.C.; Glass, D.J.; et al. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef]
- Murphy, M.M.; Lawson, J.A.; Mathew, S.J.; Hutcheson, D.A.; Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 2011, 138, 3625–3637. [Google Scholar] [CrossRef] [Green Version]
- Brigant, B.; Demont, Y.; Ouled-Haddou, H.; Metzinger-Le Meuth, V.; Testelin, S.; Garcon, L.; Metzinger, L.; Rochette, J. TRIM37 is highly expressed during mitosis in CHON-002 chondrocytes cell line and is regulated by miR-223. Bone 2020, 137, 115393. [Google Scholar] [CrossRef]
- Vishnyakova, K.S.; Babizhayev, M.A.; Aliper, A.M.; Buzdin, A.A.; Kudryavzeva, A.V.; Yegorov, Y.E. Stimulation of cell proliferation by carnosine: Cell and transcriptome approaches. Mol. Biol. 2014, 48, 718–726. [Google Scholar] [CrossRef]
- Zhang, Z.; Miao, L.; Wu, X.; Liu, G.; Peng, Y.; Xin, X.; Jiao, B.; Kong, X. Carnosine Inhibits the Proliferation of Human Gastric Carcinoma Cells by Retarding Akt/mTOR/p70S6K Signaling. J. Cancer 2014, 5, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Qi, X.; Fang, S.; Jin, Y.; Han, X.; Wang, Y.; Wang, A.; Zhou, H. Carnosine inhibits high glucose-induced mesangial cell proliferation through mediating cell cycle progression. Regul. Pept. 2009, 154, 69–76. [Google Scholar] [CrossRef]
- Palin, M.F.; Lapointe, J.; Gariepy, C.; Beaudry, D.; Kalbe, C. Characterisation of intracellular molecular mechanisms modulated by carnosine in porcine myoblasts under basal and oxidative stress conditions. PLoS ONE 2020, 15, e0239496. [Google Scholar] [CrossRef]
- Zhang, P.; Liang, X.; Shan, T.; Jiang, Q.; Deng, C.; Zheng, R.; Kuang, S. mTOR is necessary for proper satellite cell activity and skeletal muscle regeneration. Biochem. Biophys. Res. Commun. 2015, 463, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Hu, W.; de Stanchina, E.; Teresky, A.K.; Jin, S.; Lowe, S.; Levine, A.J. The regulation of AMPK beta1, TSC2, and PTEN expression by p53: Stress, cell and tissue specificity, and the role of these gene products in modulating the IGF-1-AKT-mTOR pathways. Cancer Res. 2007, 67, 3043–3053. [Google Scholar] [CrossRef] [Green Version]
- Go, G.Y.; Jo, A.; Seo, D.W.; Kim, W.Y.; Kim, Y.K.; So, E.Y.; Chen, Q.; Kang, J.S.; Bae, G.U.; Lee, S.J. Ginsenoside Rb1 and Rb2 upregulate Akt/mTOR signaling-mediated muscular hypertrophy and myoblast differentiation. J. Ginseng Res. 2020, 44, 435–441. [Google Scholar] [CrossRef]
- Cong, X.X.; Gao, X.K.; Rao, X.S.; Wen, J.; Liu, X.C.; Shi, Y.P.; He, M.Y.; Shen, W.L.; Shen, Y.; Ouyang, H.; et al. Rab5a activates IRS1 to coordinate IGF-AKT-mTOR signaling and myoblast differentiation during muscle regeneration. Cell Death Differ. 2020, 27, 2344–2362. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | % | Nutrition Component | Content |
|---|---|---|---|
| Corn | 46.90 | ME (MJ/kg) | 13.24 |
| Corn gluten meal | 5.70 | CP (%) | 22.7 |
| Extruded soybean | 20.00 | Ca (%) | 0.98 |
| Soybean meal | 20.00 | Total phosphorus (%) | 0.64 |
| Limestone | 1.20 | Available phosphorus (%) | 0.44 |
| Dicalcium phosphate | 1.70 | Met (%) | 0.50 |
| Salt | 0.30 | Met+Cys (%) | 0.90 |
| Corn oil | 2.70 | Lys (%) | 1.12 |
| Premix | 1.00 | ME (MJ/kg) | 13.24 |
| Bentonite | 0.50 | ||
| Total | 100.00 |
| Control | ALA | HIS | ALA+HIS | L-Carnosine | SEM | p-Value | |
|---|---|---|---|---|---|---|---|
| ADFI (kg) | 2.86 ab | 2.72 ab | 2.49 b | 2.57 ab | 2.96 a | 0.14 | 0.036 |
| ADG (kg) | 0.773 b | 0.780 b | 0.769 b | 0.862 a | 0.875 a | 0.02 | 0.015 |
| Feed: gain ratio | 3.71 | 3.48 | 3.23 | 2.99 | 3.36 | 0.32 | 0.078 |
| Loin eye area (cm2) | 38.74 | 39.35 | 38.65 | 40.67 | 41.82 | 2.34 | 0.321 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Shen, W.; Liu, T.; Mosenthin, R.; Bao, Y.; Chen, P.; Hao, W.; Zhao, L.; Zhang, J.; Ji, C.; et al. Improved Satellite Cell Proliferation Induced by L-Carnosine Benefits Muscle Growth of Pigs in Part through Activation of the Akt/mTOR/S6K Signaling Pathway. Agriculture 2022, 12, 988. https://doi.org/10.3390/agriculture12070988
Liu Y, Shen W, Liu T, Mosenthin R, Bao Y, Chen P, Hao W, Zhao L, Zhang J, Ji C, et al. Improved Satellite Cell Proliferation Induced by L-Carnosine Benefits Muscle Growth of Pigs in Part through Activation of the Akt/mTOR/S6K Signaling Pathway. Agriculture. 2022; 12(7):988. https://doi.org/10.3390/agriculture12070988
Chicago/Turabian StyleLiu, Yaojun, Wenqiang Shen, Tao Liu, Rainer Mosenthin, Yinghui Bao, Peng Chen, Wenbo Hao, Lihong Zhao, Jianyun Zhang, Cheng Ji, and et al. 2022. "Improved Satellite Cell Proliferation Induced by L-Carnosine Benefits Muscle Growth of Pigs in Part through Activation of the Akt/mTOR/S6K Signaling Pathway" Agriculture 12, no. 7: 988. https://doi.org/10.3390/agriculture12070988