
The Effect of Plant Growth-Promoting Rhizobacteria on Soil Properties and the Physiological and Anatomical Characteristics of Wheat under Water-Deficit Stress Conditions
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Soil
2.2. Soil Analysis
2.3. Plant Growth and Physiological Traits
2.4. Anatomical Structure of Flag Leaf
2.5. Yield and Yield Parameters
2.6. Statistical Analysis
3. Results
3.1. Impact of PGPR on Soil Characters
3.2. Effect of Water Stress, Inorganic N Fertilization, and PGPR on Wheat Growth
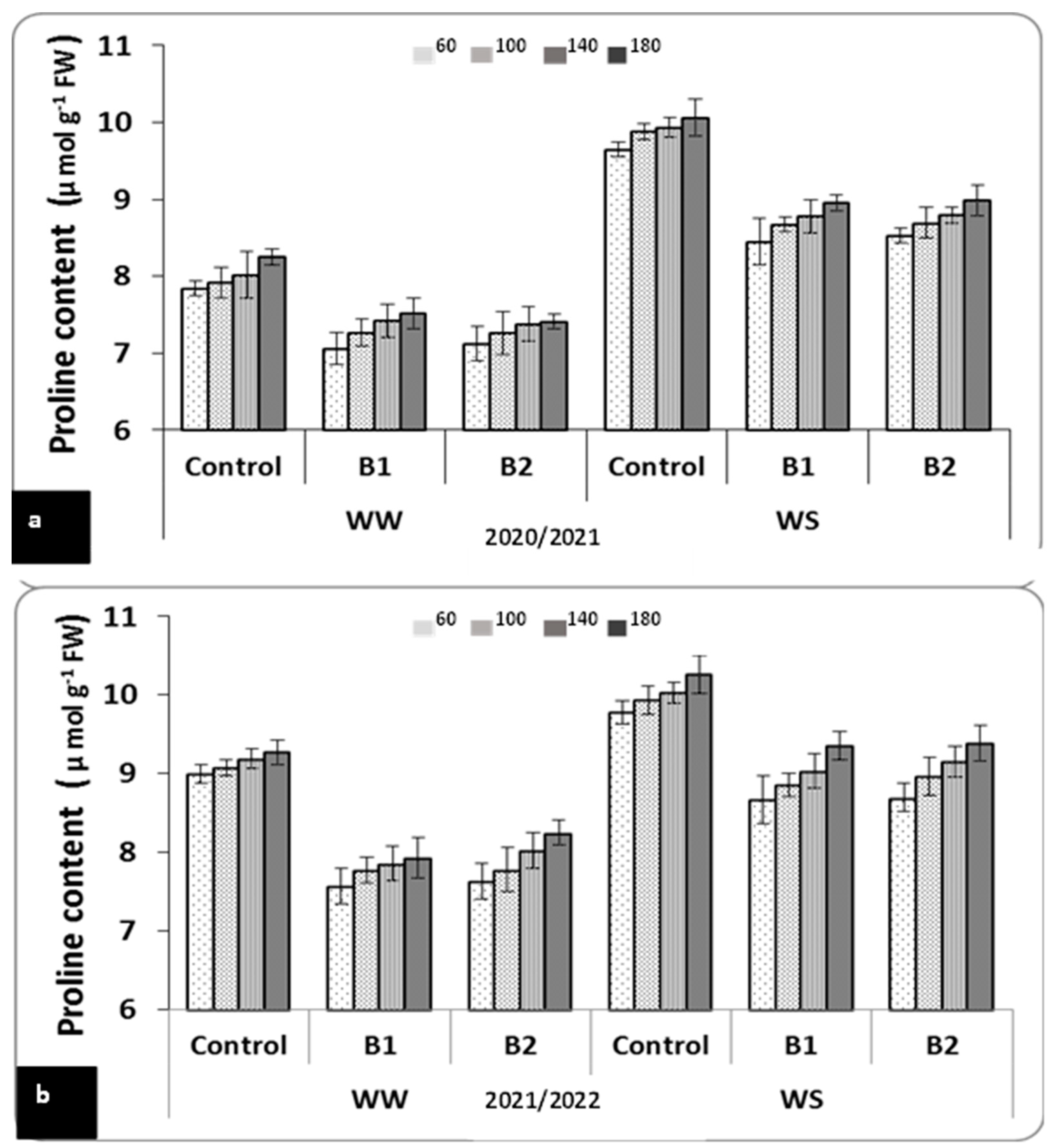
3.3. Effect of Water Stress, Inorganic N Fertilization, and PGPR on Some Physiological Traits of Wheat
3.4. Effect of Water Stress, Inorganic N Fertilization, and PGPR on Yield and Related Traits of Wheat
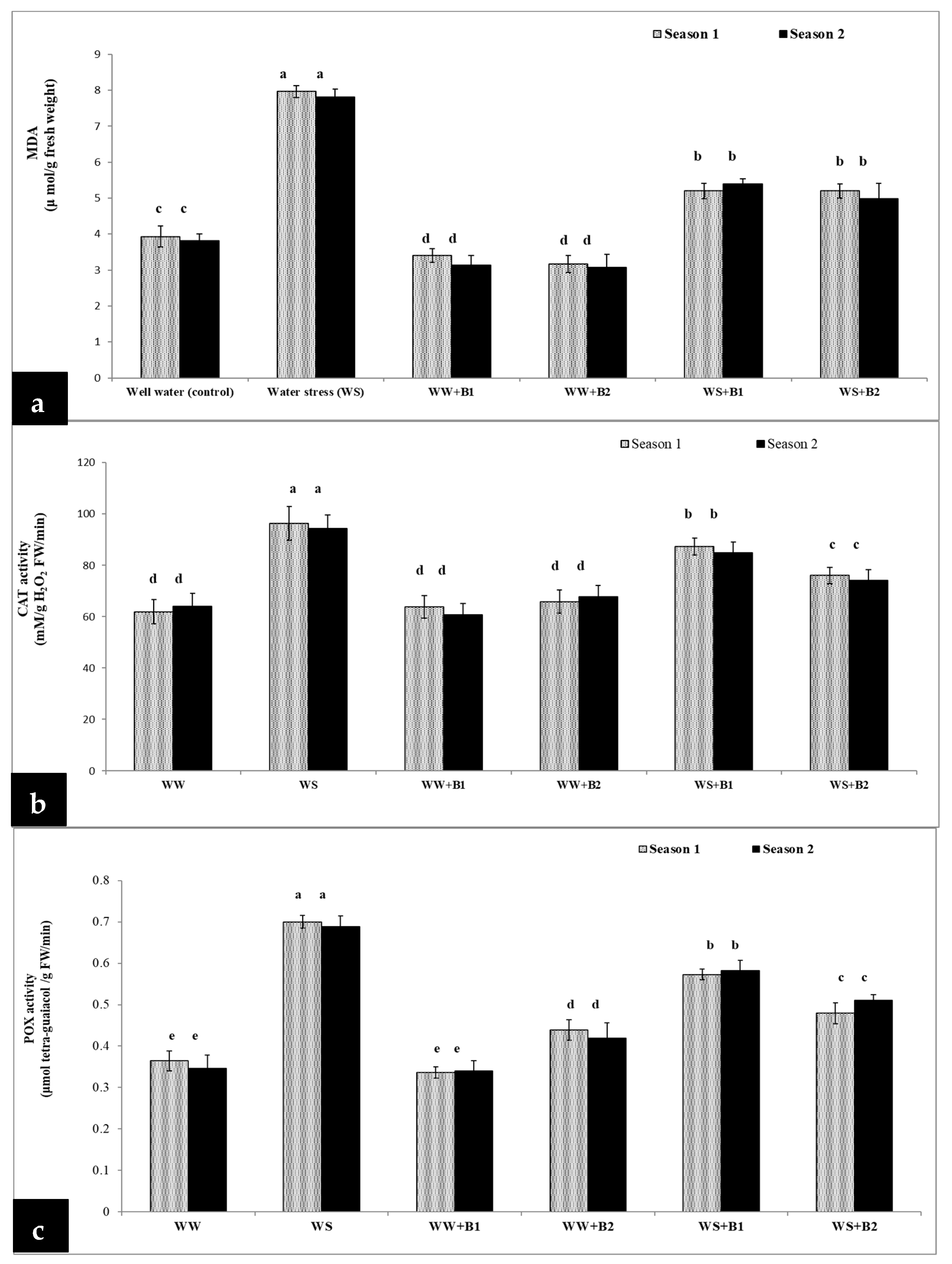
3.5. Effect of Water Stress, N Levels, and PGPR on CAT, POX and Malondialdehyde (MDA) in Wheat Plants
3.6. Effect of Water Stress, N Levels, and PGPR on Anatomical Leaf Structure
3.7. Effect of Water Stress, N Levels, and PGPR on Grain N Uptake and NUE in Wheat
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Egypt-Wheat Sector Review. 2019. Available online: http://www.fao.org/3/a-i4898e.pdf (accessed on 10 October 2022).
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb. Ecol. 2009, 58, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) growth enhancement by Azospirillum sp. under drought stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. PGPRs function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Arough, Y.K.H.; Raouf, S.S.; Reza, S.S. Bio fertilizers and zinc effects on some physiological parameters of triticale under water-limitation condition. J. Plant Interact. 2016, 11, 167–177. [Google Scholar] [CrossRef]
- Hafez, Y.M.; Attia, K.A.; Kamel, S.; Alamery, S.; El-Gendy, S.; Al-Dosse, A.; Mehiar, F.; Ghazy, A.; Abdelaal, K.A.A. Bacillus subtilis as a bio-agent combined with nano molecules can control powdery mildew disease through histochemical and physiobiochemical changes in cucumber plants. Physiol. Mol. Plant Pathol. 2020, 111, 101489. [Google Scholar] [CrossRef]
- ALKahtani, M.D.F.; Fouda, A.; Attia, K.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.; Hijri, M.; St-Arnaud, M.; Hassan, S.; Khan, N.; et al. Isolation and Characterization of Plant Growth Promoting Endophytic Bacteria from Desert Plants and Their Application as Bioinoculants for Sustainable Agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Schütz, L.; Gattinger, A.; Meier, M.; Müller, A.; Boller, T.; Mäder, P.; Mathimaran, N. Improving crop yield and nutrient use efficiency via biofertilization—A global meta-analysis. Front. Plant Sci. 2018, 8, 22–31. [Google Scholar] [CrossRef]
- Varga, B.; Vida, G.; Varga, E.; Bencze, S.; Veisz, O. Effect of simulating drought in various phenophases on the water use efficiency of winter wheat. J. Agron. Crop Sci. 2015, 201, 1–9. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A. Effect of Salicylic acid and Abscisic acid on morpho-physiological and anatomical characters of faba bean plants (Vicia faba L.) under drought stress. J. Plant Prod. Mansoura Univ. 2015, 6, 1771–1788. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Hafez, Y.M.; El-Afry, M.M.; Tantawy, D.S.; Alshaal, T. Effect of some osmoregulators on photosynthesis, lipid peroxidation, antioxidative capacity and productivity of barley (Hordeum vulgare L.) under water deficit stress. Environ. Sci. Pollut. Res. 2018, 25, 30199–30211. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.-S.E.; Abu-Elsaoud, M.A.; Hafez, Y.M. Exogenous Application of Proline and Salicylic Acid can Mitigate the Injurious Impacts of Drought Stress on Barley Plants Associated with Physiological and Histological Characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef]
- Helaly, M.N.; Mohammed, Z.; El-Shaeery, N.I.; Abdelaal, K.A.A.; Nofal, I.E. Cucumber grafting onto pumpkin can represent an interesting tool to minimize salinity stress. Physiological and anatomical studies. Middle East J. Agric. Res. 2017, 6, 953–975. [Google Scholar]
- Abdelaal, K.A.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef]
- El-Shawa, G.M.R.; Rashwan, E.M.; Abdelaal, K.A.A. Mitigating salt stress effects by exogenous application of proline and yeast extract on morphophysiological, biochemical and anatomical characters of calendula plants. Sci. J. Flowers Ornam. Plants 2020, 7, 461–482. [Google Scholar] [CrossRef]
- Hafez, Y.M.; Abdelaal, K.A.A. Investigation of susceptibility and resistance mechanisms of some Egyptian wheat cultivars (Triticum aestivum L.) inoculated with Blumeria graminis f. sp. tritici using certain biochemical, molecular characterization and SEM. J. Plant Prot. Pathol. Mansoura Univ. 2015, 6, 443–454. [Google Scholar]
- Omara, R.I.; El-Kot, G.; Fadel, F.M.; Abdelaal, K.A.A.; Saleh, E. Efficacy of certain bioagents on patho-physiological characters of wheat plants under wheat leaf rust stress. Physiol. Mol. Plant Pathol. 2019, 106, 102–108. [Google Scholar] [CrossRef]
- Hafez, E.M.; Gharib, H.S. Effect of exogenous application of ascorbic acid on physiological and biochemical characteristics of wheat under water stress. Int. J. Plant Prod. 2016, 10, 579–596. [Google Scholar]
- Abdelaal, K.A.A.; Hafez, Y.M.; El Sabagh, A.; Saneoka, H. Ameliorative effects of Abscisic acid and yeast on morpho-physiological and yield characteristics of maize plant (Zea mays L.) under water deficit conditions. Fresenius Environ. Bull. 2017, 26, 7372–7383. [Google Scholar]
- Abdelaal, K.A.A.; Elafry, M.; Abdel-Latif, I.; Elshamy, R.; Hassan, M.; Hafez, Y. Pivotal role of yeast and ascorbic acid in improvement the morpho-physiological characters of two wheat cultivars under water deficit stress in calcareous soil. Fresenius Environ. Bull. 2021, 30, 2554–2565. [Google Scholar]
- Hafez, E.M.; Kobata, T. The effect of different nitrogen sources from urea and ammonium sulfate on the spikelet number in Egyptian spring wheat cultivars on well-watered pot soils. Plant Prod. Sci. 2012, 15, 332–338. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; AlKahtani, M.D.F.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The pivotal role of plant growth promoting bacteria in alleviating the adverse effects of drought on plants. Biology 2021, 10, 520. [Google Scholar] [CrossRef] [PubMed]
- AlKahtani, M.D.F.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Husnain, L.A.; AlGwaiz, H.I.M.; Abdelaal, K.A.A. Evaluation of Silicon and Proline Application on the Oxidative Machinery in Drought-Stressed Sugar Beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef] [PubMed]
- Kahiluoto, H.; Kuisma, M.; Kuokkanen, A.; Mikkilä, M.; Linnanen, L. Taking planetary nutrient boundaries seriously: Can we feed the people? Glob. Food Sec. 2014, 3, 16–21. [Google Scholar] [CrossRef]
- Klute, A. Methods of Soil Analysis. In Physical and Mineralogical Methods, 2nd ed.; Part 1; ASA, SSSA, Inc.: Madison, WI, USA, 2000. [Google Scholar]
- Yin, X.; Struik, P.C.; Romero, P.; Harbinson, J.; Evers, J.B.; Van, D.P.; Vos, J. Using combined measurements of gas exchange and chlorophyll fluorescence to estimate parameters of a biochemical C photosynthesis model: A critical appraisal and a new integrated approach applied to leaves in wheat (Triticum aestivum) canopy. Plant Cell Environ. 2009, 32, 448–464. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A. Pivotal Role of Bio and Mineral Fertilizer Combinations on Morphological, Anatomical and Yield Characters of Sugar Beet Plant (Beta vulgaris L.). Middle East J. Agric. Res. 2015, 4, 717–734. [Google Scholar]
- Abdelaal, K.A.A.; Badawy, S.; Abdel Aziz, R.M.; Neana, S. Effect of mineral nitrogen levels and biofertilizer on morphophysiological characters of three sweet sorghum varieties (Sorghum bicolor L. Moench). J. Plant Prod. Mansoura Univ. 2015, 6, 189–203. [Google Scholar]
- Hafez, E.M.; Ragab, Y.; Kobata, T. Water-use efficiency and ammonium-N source applied of wheat under irrigated and desiccated conditions. Int. J. Plant Soil Sci. 2014, 3, 1302. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem. 2004, 42, 565. [Google Scholar] [CrossRef]
- ALKahtani, M.D.F.; Attia, K.A.; Hafez, Y.M.; Khan, N.; Eid, A.M.; Ali, M.A.M.; Abdelaal, K.A.A. Chlorophyll Fluorescence Parameters and Antioxidant Defense System Can Display Salt Tolerance of Salt Acclimated Sweet Pepper Plants Treated with Chitosan and Plant Growth Promoting Rhizobacteria. Agronomy 2020, 10, 1180. [Google Scholar] [CrossRef]
- Guarda, G.; Padovan, S.; Delogu, G. Grain yield, nitrogen use efficiency and baking quality of old and modern Italian bread wheat cultivars grown at different nitrogen levels. Eur. J. Agron. 2004, 21, 181–192. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Hawkesford, M.J.; Barraclough, P.B.; Holdsworth, M.J.; Kerr, S.; Kightley, S.; Shewry, P.R. Identifying traits to improve the nitrogen economy of wheat: Recent advances and future prospects. Field Crops Res. 2009, 114, 329–342. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Pierotti, C.F. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Daughtry, C.S.T.; Hollinger, S.E. Costs of measuring leaf area index of corn. Agron. J. 1984, 76, 836–841. [Google Scholar] [CrossRef]
- Alami, Y.; Achouak, W.; Marol, C.; Heulin, T. Rhizosphere soil aggregation and plant growth promotion of sunflowers by an exopolysaccharide-producing Rhizobium sp. strain isolated from sunflower roots. Appl. Environ. Microbiol. 2000, 66, 3393–3398. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, F.J.; de Andrés, E.F.; Tenorio, J.L.; Ayerbe, L. Growth of epicotyls, turgor maintenance and osmotic adjustment in pea plants (Pisum sativum L.) subjected to water stress. Field Crops Res. 2004, 86, 81–90. [Google Scholar] [CrossRef]
- Selvakumar, G.; Panneerselvam, P.; Ganeshamurthy, A.N. Bacterial mediated alleviation of abiotic stress in crops. In Bacteria in Agrobiology, Stress Management; Springer: Berlin/Heidelberg, Germany, 2012; pp. 205–224. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase. “Methods of Enzymatic Analysis”, 3rd ed.; Verlag Chemie: Weinheim, Germany, 1983; p. 273. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Davenport, S.B.; Gallego, S.M.; Benavides, M.P.; Tomaro, M.L. Behaviour of antioxidant defense system in the adaptive response to salt stress in Helianthus annuus L. cells. Plant Growth Regul. 2003, 40, 81–88. [Google Scholar] [CrossRef]
- Nassar, M.A.; El-Sahhar, K.F. Botanical Preparations and Microscopy (Microtechnique); Academic Bookshop: Giza, Egypt, 1998; p. 219. [Google Scholar]
- El-Banna, M.F.; Abdelaal, K.A.A. Response of Strawberry Plants Grown in the Hydroponic System to Pretreatment with H2O2 Before Exposure to Salinity Stress. J. Plant Prod. Mansoura Univ. 2018, 9, 989–1001. [Google Scholar] [CrossRef]
- Abdelaal, K.A.A.; El Menofy, E.M.; Nessem, A.A.; Elhaak, M.A. The allelopathy potential and glyphosate influence on anatomical features of Egyptian clover plant (Trifolium alexandrinum L.) infested with dodder weed (Cuscuta campestris L.). Fresenius Environ. Bull. 2019, 28, 1262–1269. [Google Scholar]
- Abdelaal, K.A.A.; Attia, K.A.; Niedbała, G.; Wojciechowski, T.; Hafez, Y.; Alamery, S.; Alateeq, T.K.; Arafa, S.A. Mitigation of Drought Damages by Exogenous Chitosan and Yeast Extract with Modulating the Photosynthetic Pigments, Antioxidant Defense System and Improving the Productivity of Garlic Plants. Horticulturae 2021, 7, 510. [Google Scholar] [CrossRef]
- Arafa, S.A.; Attia, K.A.; Niedbała, G.; Piekutowska, M.; Alamery, S.; Abdelaal, K.; Alateeq, T.K.; Ali, M.A.M.; Elkelish, A.; Attallah, S.Y. Seed Priming Boost Adaptation in Pea Plants under Drought Stress. Plants 2021, 10, 2201. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.; Omara, A.E.D.; Ahmed, A. The Coupling Effects of Plant Growth Promoting Rhizobacteria and Salicylic Acid on Physiological Modifications, Yield Traits, and Productivity of Wheat under Water Deficient Conditions. Agronomy 2019, 9, 524. [Google Scholar] [CrossRef]
- Moutia, J.F.Y.; Saumtally, S.; Spaepen, S.; Vanderleyden, J. Plant growth promotion by Azospirillum sp. in sugarcane is influenced by genotype and drought stress. Plant Soil 2010, 337, 233–242. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Combaret, C.P. Plant growth promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef]
- Sandhya, V.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Abou-Attia, F.A.M.; Abdelaal, K.A.A. Effect of Bio and Mineral fertilization on the main insect pests and some characters of sugar beet plants. J. Agric. Sci. Mansoura Univ. 2007, 32, 1471–1485. [Google Scholar] [CrossRef]
- Hafez, E.; Badawy, S. Effect of Bio Fertilizers and Inorganic Fertilizers on Growth, Productivity and Quality of Bread Wheat Cultivars. Cercet. Agron. Mold. 2018, 176, 1–16. [Google Scholar] [CrossRef]
- Wang, J.; Fu, P.; Lu, W.; Lu, D. Application of moderate nitrogen levels alleviates yield loss and grain quality deterioration caused by post-silking heat stress in fresh waxy maize. Crop J. 2020, 8, 1081–1092. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.; Rashwan, E.; Kamara, M.M. Synergistic Effect of Biochar and Plant Growth Promoting Rhizobacteria on Alleviation of Water Deficit in Rice Plants under Salt-Affected Soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Abdelaal, K.A.A.; Barutcular, C. Impact of antioxidants supplementation on growth, yield and quality traits of canola (Brassica napus L.) under irrigation intervals in North Nile Delta of Egypt. J. Exp. Biol. Agric. Sci. 2017, 5, 163–172. [Google Scholar] [CrossRef]
- Hota, T.; Pradhan, C.; Rout, G.R. Identification of drought tolerant Sesamum genotypes using biochemical markers. Indian J. Exp. Biol. 2019, 57, 690. [Google Scholar]
- Khaffagy, A.E.; Mazrou, Y.S.A.; Morsy, A.R.; El-Mansoury, M.A.M.; El-Tokhy, A.I.; Hafez, Y.; Abdelaal, K.; Khedr, R.A. Impact of Irrigation Levels and Weed Control Treatments on Annual Weeds, Physiological Traits and Productivity of Soybean under Clay Soil Conditions. Agronomy 2022, 12, 1037. [Google Scholar] [CrossRef]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.A.A.; Hafez, Y.M. Histological and biochemical aspects of compatible and incompatible wheat-Puccinia striiformis interactions. Physiol. Molecular Plant Pathol. 2019, 106, 120–128. [Google Scholar] [CrossRef]
- Hafez, Y.M.; Attia, K.A.; Alamery, S.; Ghazy, A.; Al-Dosse, A.; Ibrahim, E.; Rashwan, E.; El-Maghraby, L.; Awad, A.; Abdelaal, K.A.A. Beneficial Effects of Biochar and Chitosan on Antioxidative Capacity, Osmolytes Accumulation, and Anatomical Characters of Water-Stressed Barley Plants. Agronomy 2020, 10, 630. [Google Scholar] [CrossRef]
- Abdelaal, K.; Mazrou, Y.; Hafez, Y. Effect of slicon and carrot extract on morphophysiological characters of pea (Pisum sativum L.) under salinity stress conditions. Fresenius Environ. Bull. 2022, 31, 608–615. [Google Scholar]
- Al-Shammari, W.B.; Altamimi, H.R.; Abdelaal, K. Improvement in Physiobiochemical and Yield Characteristics of Pea Plants with Nano Silica and Melatonin under Salinity Stress Conditions. Horticulturae 2023, 9, 711. [Google Scholar] [CrossRef]
- El Nahhas, N.; AlKahtani, M.; Abdelaal, K.A.A.; Al Husnain, L.; AlGwaiz, H.; Hafez, Y.M.; Attia, K.; El-Esawi, M.; Ibrahim, M.; Elkelish, A. Biochar and jasmonic acid application attenuates antioxidative systems and improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) irrigated with saline water. Plant Physiol. Biochem. 2021, 166, 807–817. [Google Scholar] [CrossRef]
- Mosalem, M.; Mazrou, Y.; Badawy, S.; Abd Ullah, M.A.; Mubarak, M.G.; Hafez, Y.M.; Abdelaal, K.A. Evaluation of sowing methods and nitrogen levels for grain yield and components of durum wheat under arid regions of Egypt. Rom Biotechnol. Lett. 2021, 26, 3031–3039. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Mazrou, Y.; El-Henawy, A.S.; Nabil, A.M.M.; Moghanam, A.A.; Hafez, Y.; Abdelaal, K. Effect of boron and potassium levels on growth and yield of sugar beet under irrigation deprivation conditions. Fresenius Environ. Bull. 2022, 31, 3623–3632. [Google Scholar]
- Abdelaal, K.; Alsubeie, M.S.; Hafez, Y.; Emeran, A.; Moghanm, F.; Okasha, S.; Omara, R.; Basahi, M.A.; Darwish, D.B.E.; Ibrahim, M.F.M.; et al. Physiological and Biochemical Changes in Vegetable and Field Crops under Drought, Salinity and Weeds Stresses: Control Strategies and Management. Agriculture 2022, 12, 2084. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Naveed, M.; Nawaz, A.; Shahzad, B. Seed priming of Zn with endophytic bacteria improves the productivity and grain biofortification of bread wheat. Eur. J. Agron. 2018, 94, 98–107. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2020/2021 | 2021/2022 | |||||||
|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Precipitation (mm) | RH (%) | Temperature (°C) | Precipitation (mm) | RH (%) | |||
| max | min | max | min | |||||
| December | 25.8 | 10.4 | 0.86 | 31.3 | 26.9 | 11.4 | 0.80 | 30.7 |
| January | 22.6 | 9.4 | 3.54 | 42.3 | 23.5 | 9.4 | 3.50 | 41.4 |
| February | 21.2 | 10.2 | 7.10 | 46.3 | 22.0 | 11.0 | 7.00 | 45.1 |
| March | 23.3 | 15.1 | 0.60 | 39.5 | 24.0 | 16.2 | 0.30 | 37.2 |
| April | 26.5 | 16.1 | 0.50 | 38.0 | 27.4 | 17.2 | 0.20 | 40.2 |
| May | 30.8 | 16.7 | 0.00 | 36.9 | 31.9 | 17.8 | 0.00 | 37.9 |
| Parameters | FC (%) | WP (%) | BD (g cm−3) | Available H2O (Gravimetric, %) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Time | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | |
| Before sowing | 42.2 | 44.0 | 22.9 | 21.7 | 1.2 | 1.3 | 19.2 | 25.7 | |
| At harvest (B1) | 52.1 | 62.7 | 19.6 | 18.4 | 1.0 | 1.1 | 37.2 | 41.6 | |
| At harvest (B2) | 58.7 | 63.7 | 20.6 | 19.9 | 1.0 | 1.2 | 38.1 | 43.8 | |
| S.E.M. | 1.02 | 1.06 | 0.43 | 0.32 | 0.04 | 0.05 | 1.00 | 0.93 | |
| Treatments | LAI | Flag Leaf Area (cm2) | Chlorophyll Content (SPAD) | |||
|---|---|---|---|---|---|---|
| S1 | S2 | S1 | S2 | S1 | S2 | |
| Water treatments (W) | ||||||
| WW | 5.34 a | 3.72 a | 40.42 a | 38.48 a | 40.10 a | 41.92 a |
| WS | 5.11 b | 3.23 b | 37.78 b | 36.81 b | 37.41 b | 37.80 b |
| N-levels (N) (kg ha−1) | ||||||
| 60 | 4.51 d | 3.32 d | 30.14 d | 26.19 d | 31.92 d | 33.79 d |
| 100 | 5.09 c | 3.49 c | 36.37 c | 35.15 c | 36.24 c | 38.23 c |
| 140 | 5.42 b | 3.72 b | 42.42 b | 43.70 b | 41.27 b | 41.94 b |
| 180 | 6.08 a | 3.96 a | 47.36 a | 45.54 a | 45.59 a | 47.45 a |
| PGPR (B) | ||||||
| B1 | 4.19 c | 3.04 b | 36.14 c | 36.45 b | 36.24 c | 37.15 b |
| B2 | 5.23 b | 3.62 ab | 39.05 b | 38.10 a | 38.93 b | 40.84 a |
| ANOVA | ||||||
| W | * | ** | ** | * | ** | ** |
| N | ** | ** | ** | ** | ** | ** |
| B | ** | ** | ** | * | ** | ** |
| W × N | ns | ns | ns | ns | ns | ns |
| W × B | ns | ns | ns | ns | ns | ns |
| N × B | ns | ns | ns | ns | ns | ns |
| W × N × B | ns | ns | ns | ns | ns | ns |
| Treatments | Plant Height (cm) | Spike Length (cm) | No. Spikes m−2 | No. Grains Spike−1 | 1000-Grain Weight (g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | |
| Water treatments (W) | ||||||||||
| WW | 107.99 a | 107.61 a | 13.92 a | 13.83 a | 392.78 a | 379.76 a | 57.37 a | 59.51 a | 52.54 a | 53.30 a |
| WS | 96.01 b | 93.85 b | 10.07 b | 10.03 b | 376.14 b | 364.91 b | 50.90 b | 53.73 b | 47.61 b | 46.89 b |
| Nitrogen (N) (kg ha−1) | ||||||||||
| 60 | 91.22 d | 88.20 d | 9.85 c | 9.70 b | 333.83 c | 302.01 c | 49.39 c | 51.11 c | 48.11 d | 47.31 d |
| 100 | 98.52 c | 97.35 c | 10.79 b | 9.93 b | 358.00 b | 348.03 b | 53.63 b | 53.87 b | 49.02 c | 48.45 c |
| 140 | 106.57 b | 105.91 b | 13.43 a | 13.42 a | 418.02 a | 413.00 a | 56.13 a | 60.64 a | 50.23 b | 51.47 b |
| 180 | 111.69 a | 111.4 a | 13.92 a | 13.88 a | 428.03 a | 426.00 a | 57.39 a | 60.86 a | 52.94 a | 51.15 a |
| PGPR (B) | 99.57 c | 99.31 b | 10.26 b | 10.03 b | 359.71 b | 309.09 b | 50.19 b | 54.32 b | 47.90 b | 47.88 b |
| B1 | 102.32 b | 101.14 a | 12.71 a | 12.78 a | 396.00 a | 395.60 a | 55.89 a | 57.73 a | 51.12 a | 49.82 a |
| B2 | 104.10 a | 101.75 a | 13.02 a | 12.98 a | 398.00 a | 411.81 a | 56.33 a | 57.81 a | 51.20 a | 51.09 a |
| ANOVA | ||||||||||
| W | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| N | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| B | ** | * | * | * | ** | ** | ** | * | ns | ns |
| W N | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| W B | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| NB | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| WNB | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Treatments | Plant Height (cm) | Spike Length (cm) | No. Spikes m−2 | No. Grains Spike−1 | 1000-Grain Weight (g) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | |
| Water treatments (W) | ||||||||||
| WW | 107.99 a | 107.61 a | 13.92 a | 13.83 a | 392.78 a | 379.76 a | 57.37 a | 59.51 a | 52.54 a | 53.30 a |
| WS | 96.01 b | 93.85 b | 10.07 b | 10.03 b | 376.14 b | 364.91 b | 50.90 b | 53.73 b | 47.61 b | 46.89 b |
| Nitrogen (N) (kg ha−1) | ||||||||||
| 60 | 91.22 d | 88.20 d | 9.85 c | 9.70 b | 333.83 c | 302.01 c | 49.39 c | 51.11 c | 48.11 d | 47.31 d |
| 100 | 98.52 c | 97.35 c | 10.79 b | 9.93 b | 358.00 b | 348.03 b | 53.63 b | 53.87 b | 49.02 c | 48.45 c |
| 140 | 106.57 b | 105.91 b | 13.43 a | 13.42 a | 418.02 a | 413.00 a | 56.13 a | 60.64 a | 50.23 b | 51.47 b |
| 180 | 111.69 a | 111.4 a | 13.92 a | 13.88 a | 428.03 a | 426.00 a | 57.39 a | 60.86 a | 52.94 a | 51.15 a |
| PGPR (B) | 99.57 c | 99.31 b | 10.26 b | 10.03 b | 359.71 b | 309.09 b | 50.19 b | 54.32 b | 47.90 b | 47.88 b |
| B1 | 102.32 b | 101.14 a | 12.71 a | 12.78 a | 396.00 a | 395.60 a | 55.89 a | 57.73 a | 51.12 a | 49.82 a |
| B2 | 104.10 a | 101.75 a | 13.02 a | 12.98 a | 398.00 a | 411.81 a | 56.33 a | 57.81 a | 51.20 a | 51.09 a |
| ANOVA | ||||||||||
| W | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| N | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| B | ** | * | * | * | ** | ** | ** | * | ns | ns |
| W N | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| W B | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| NB | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| WNB | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alaskar, A.A.; AL-Shwaiman, H.A. The Effect of Plant Growth-Promoting Rhizobacteria on Soil Properties and the Physiological and Anatomical Characteristics of Wheat under Water-Deficit Stress Conditions. Agriculture 2023, 13, 2042. https://doi.org/10.3390/agriculture13112042
Alaskar AA, AL-Shwaiman HA. The Effect of Plant Growth-Promoting Rhizobacteria on Soil Properties and the Physiological and Anatomical Characteristics of Wheat under Water-Deficit Stress Conditions. Agriculture. 2023; 13(11):2042. https://doi.org/10.3390/agriculture13112042
Chicago/Turabian StyleAlaskar, Abdulaziz A., and Hind A. AL-Shwaiman. 2023. "The Effect of Plant Growth-Promoting Rhizobacteria on Soil Properties and the Physiological and Anatomical Characteristics of Wheat under Water-Deficit Stress Conditions" Agriculture 13, no. 11: 2042. https://doi.org/10.3390/agriculture13112042