Eliminating the Pathogen Xanthomonas hortorum pv. carotae from Carrot Seeds Using Different Types of Nanoparticles

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Xhc Strain and Growth Conditions
2.2. In Vitro Experiment: Evaluation of Nanomaterials for Antibacterial Activity under In Vitro Conditions
2.3. In Planta Experiment
2.3.1. Artificial Inoculation of Carrot Seeds with Xhc Strain
2.3.2. Treatment and Germination of Seeds with Nanomaterials
2.4. Detection of Xhc in Carrot Seedlings
2.5. Statistical Analysis
3. Results
3.1. Antibacterial Activity of Tested Nanomaterials In Vitro
3.1.1. MIC and MBC Determination
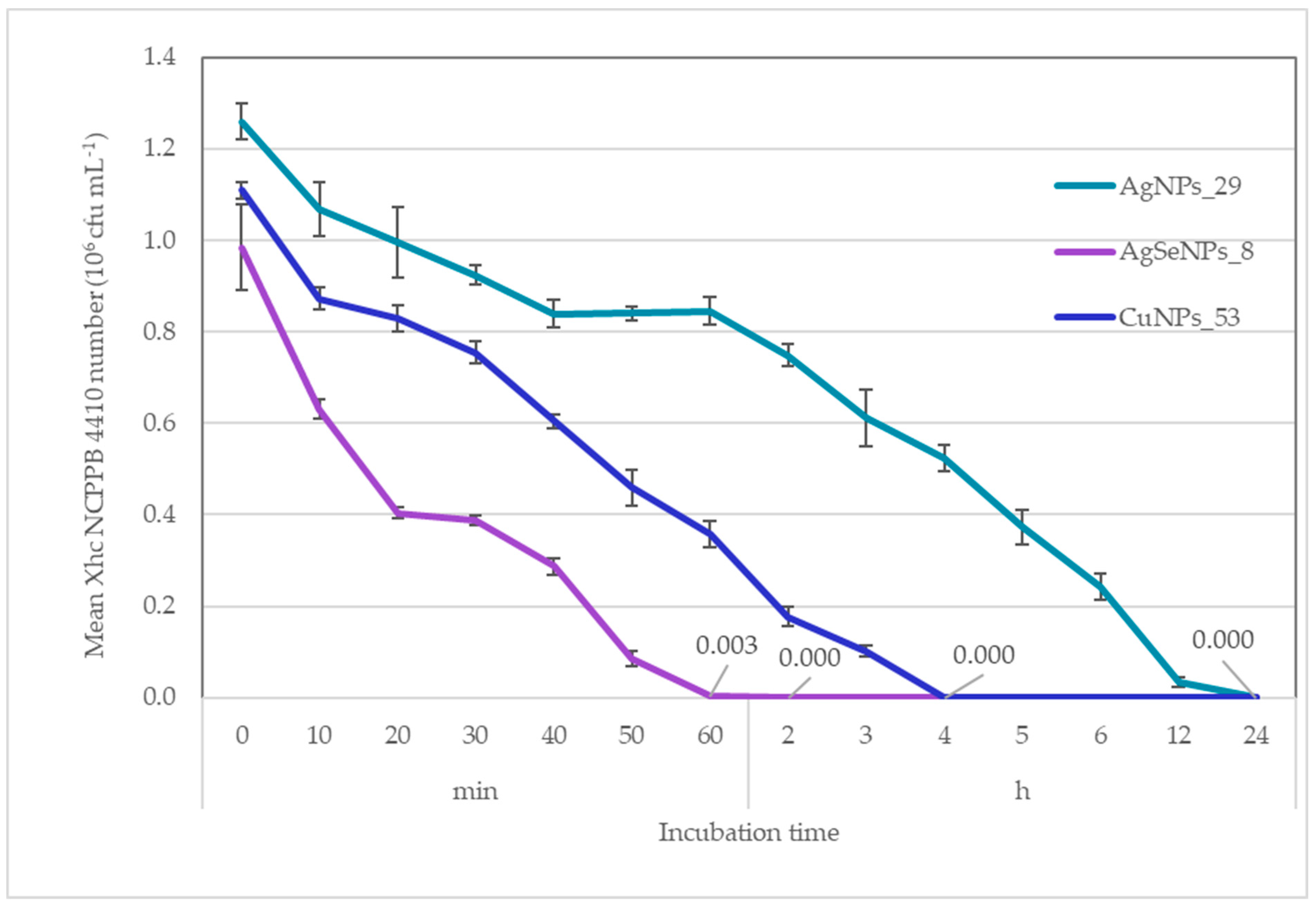
3.1.2. Treatment Duration Determined by Time-Kill Assay
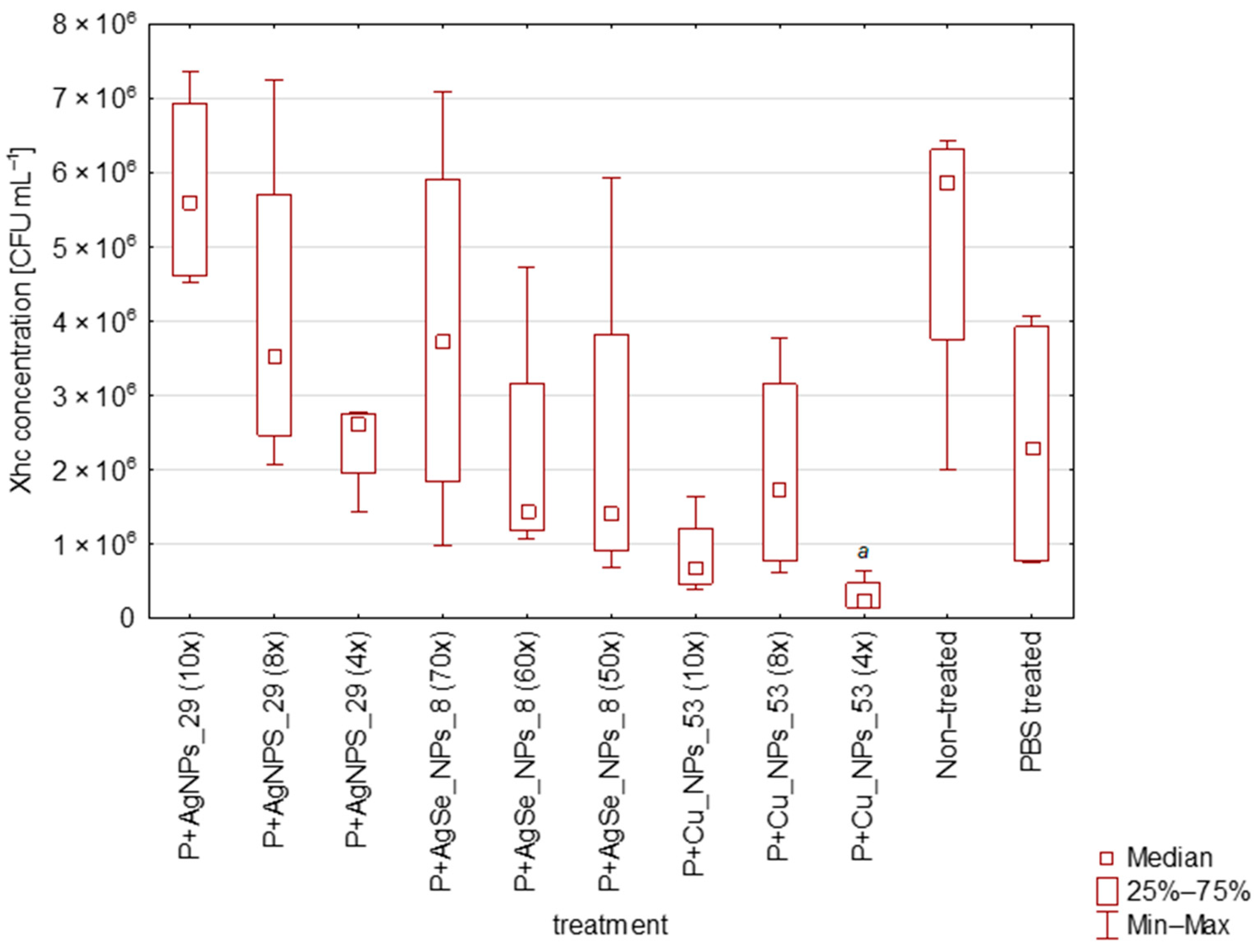
3.2. Effectiviteness of AgNPs_29, CuNPs_53, and AgSeNPs_8 in Elimination of Xhc in Planta
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Dia, N.C.; Morinière, L.; Cottyn, B.; Bernal, E.; Jacobs, J.M.; Koebnik, R.; Osdaghi, E.; Potnis, N.; Pothier, J.F. Xanthomonas hortorum–beyond gardens: Current taxonomy, genomics, and virulence repertoires. Mol. Plant Pathol. 2022, 23, 597–621. [Google Scholar] [CrossRef]
- Pruvost, O.; Boyer, C.; Robène-Soustrade, I.; Jouen, E.; Saison, A.; Hostachy, B.; Benimadhu, S. First report of Xanthomonas hortorum pv. carotae causing bacterial leaf blight of carrot in Mauritius. Plant Dis. 2010, 94, 1069. [Google Scholar] [CrossRef]
- Scott, J.C.; Dung, J.K. Distribution of Xanthomonas hortorum pv. carotae populations in naturally infested carrot seed lots. Plant Dis. 2020, 104, 2144–2148. [Google Scholar] [CrossRef]
- Christianson, C.E.; Jones, S.S.; du Toit, L.J. Screening carrot germplasm for resistance to Xanthomonas hortorum pv. carotae. HortScience 2015, 50, 341–350. [Google Scholar] [CrossRef]
- Medlock, J.M.; Leach, S.A. Effect of climate change on vector-borne disease risk in the UK. Lancet Infect. Dis. 2015, 15, 721–730. [Google Scholar] [CrossRef]
- Moumni, M.; Brodal, G.; Romanazzi, G. Recent innovative seed treatment methods in the management of seedborne pathogens. Food Secur. 2023, 15, 1365–1382. [Google Scholar] [CrossRef]
- Nega, E.; Ulrich, R.; Werner, S.; Jahn, M. Hot water treatment of vegetable seed—An alternative seed treatment method to control seed-borne pathogens in organic farming/Heißwasserbehandlung von Gemüsesaatgut—Eine alternative Saatgutbehandlungsmethode zur Bekämpfung samenbürtiger Pathogene im ökologischen Landbau. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz/J. Plant Dis. Prot. 2003, 110, 220–234. [Google Scholar]
- Mandiriza, G.; Kritzinger, Q.; Aveling, T. The evaluation of plant extracts, biocontrol agents and hot water as seed treatments to control black rot of rape in South Africa. Crop. Prot. 2018, 114, 129–136. [Google Scholar] [CrossRef]
- Pečenka, J.; Bytešníková, Z.; Kiss, T.; Peňázová, E.; Baránek, M.; Eichmeier, A.; Tekielska, D.; Richtera, L.; Pokluda, R.; Adam, V. Silver nanoparticles eliminate Xanthomonas campestris pv. campestris in cabbage seeds more efficiently than hot water treatment. Mater. Today Commun. 2021, 27, 102284. [Google Scholar] [CrossRef]
- Carisse, O.; Ouimet, A.; Toussaint, V.; Philion, V. Evaluation of the effect of seed treatments, bactericides, and cultivars on bacterial leaf spot of lettuce caused by Xanthomonas campestris pv. vitians. Plant Dis. 2000, 84, 295–299. [Google Scholar] [CrossRef]
- Kumar, A.; Choudhary, A.; Kaur, H.; Guha, S.; Mehta, S.; Husen, A. Potential applications of engineered nanoparticles in plant disease management: A critical update. Chemosphere 2022, 295, 133798. [Google Scholar] [CrossRef]
- Skrzyniarz, K.; Sanchez-Nieves, J.; de la Mata, F.J.; Łysek-Gładysińska, M.; Lach, K.; Ciepluch, K. Mechanistic insight of lysozyme transport through the outer bacteria membrane with dendronized silver nanoparticles for peptidoglycan degradation. Int. J. Biol. Macromol. 2023, 237, 124239. [Google Scholar] [CrossRef]
- Soares, E.V.; Soares, H.M. Harmful effects of metal (loid) oxide nanoparticles. Appl. Microbiol. Biotechnol. 2021, 105, 1379–1394. [Google Scholar] [CrossRef] [PubMed]
- Gahlawat, G.; Shikha, S.; Chaddha, B.S.; Chaudhuri, S.R.; Mayilraj, S.; Choudhury, A.R. Microbial glycolipoprotein-capped silver nanoparticles as emerging antibacterial agents against cholera. Microb. Cell Factories 2016, 15, 1–14. [Google Scholar] [CrossRef]
- Lee, B.; Lee, M.J.; Yun, S.J.; Kim, K.; Choi, I.-H.; Park, S. Silver nanoparticles induce reactive oxygen species-mediated cell cycle delay and synergistic cytotoxicity with 3-bromopyruvate in Candida albicans, but not in Saccharomyces cerevisiae. Int. J. Nanomed. 2019, 14, 4801–4816. [Google Scholar] [CrossRef] [PubMed]
- Mikhailova, E.O. Silver nanoparticles: Mechanism of action and probable bio-application. J. Funct. Biomater. 2020, 11, 84. [Google Scholar] [CrossRef] [PubMed]
- Adisa, I.O.; Pullagurala, V.L.R.; Peralta-Videa, J.R.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Recent advances in nano-enabled fertilizers and pesticides: A critical review of mechanisms of action. Environ. Sci. Nano 2019, 6, 2002–2030. [Google Scholar] [CrossRef]
- Ali, M.A.; Ahmed, T.; Wu, W.; Hossain, A.; Hafeez, R.; Islam Masum, M.M.; Wang, Y.; An, Q.; Sun, G.; Li, B. Advancements in plant and microbe-based synthesis of metallic nanoparticles and their antimicrobial activity against plant pathogens. Nanomaterials 2020, 10, 1146. [Google Scholar] [CrossRef] [PubMed]
- Strayer, A.; Ocsoy, I.; Tan, W.; Jones, J.; Paret, M. Low concentrations of a silver-based nanocomposite to manage bacterial spot of tomato in the greenhouse. Plant Dis. 2016, 100, 1460–1465. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, N.S.; Silva, A.C.A.; Tebaldi, N.D. Simonkolleite nanoparticles for seed treatment and control of tomato bacterial spot caused by Xanthomonas hortorum pv. gardneri. Ciênc. Agrotecnologia 2023, 47, e000623. [Google Scholar] [CrossRef]
- Norman, D.J.; Chen, J. Effect of foliar application of titanium dioxide on bacterial blight of geranium and Xanthomonas leaf spot of poinsettia. HortScience 2011, 46, 426–428. [Google Scholar] [CrossRef]
- Wohlmuth, J.; Tekielska, D.; Čechová, J.; Baránek, M. Interaction of the nanoparticles and plants in selective growth stages—Usual effects and resulting impact on usage perspectives. Plants 2022, 11, 2405. [Google Scholar] [CrossRef] [PubMed]
- Cadena, M.B.; Preston, G.M.; Van der Hoorn, R.A.; Flanagan, N.A.; Townley, H.E.; Thompson, I.P. Enhancing cinnamon essential oil activity by nanoparticle encapsulation to control seed pathogens. Ind. Crop. Prod. 2018, 124, 755–764. [Google Scholar] [CrossRef]
- Kaur, P.; Thakur, R.; Choudhary, A. An in vitro study of the antifungal activity of silver/chitosan nanoformulations against important seed borne pathogens. Int. J. Sci. Technol. Res. 2012, 1, 83–86. [Google Scholar]
- Prasad, T.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.; Sajanlal, P.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Sahab, A.; Waly, A.; Sabbour, M.; Nawar, L.S. Synthesis, antifungal and insecticidal potential of Chitosan (CS)-g-poly (acrylic acid)(PAA) nanoparticles against some seed borne fungi and insects of soybean. Int. J. ChemTech Res. 2015, 8, 589–598. [Google Scholar]
- Mawale, K.S.; Nandini, B.; Giridhar, P. Copper and Silver Nanoparticle Seed Priming and Foliar Spray Modulate Plant Growth and Thrips Infestation in Capsicum spp. ACS Omega 2024, 9, 3430–3444. [Google Scholar] [CrossRef]
- Štůsková, K.; Pečenka, J.; Tekielska, D.A.; Špetík, M.; Bytešníková, Z.; Švec, P.; Ondreáš, F.; Ridošková, A.; Richtera, L.; Adam, V. The in vitro effects of selected substances and nanomaterials against Diaporthe eres, Diplodia seriata and Eutypa lata. Ann. Appl. Biol. 2023, 182, 226–237. [Google Scholar] [CrossRef]
- Bytešníková, Z.; Pečenka, J.; Tekielska, D.; Kiss, T.; Švec, P.; Ridošková, A.; Bezdička, P.; Pekárková, J.; Eichmeier, A.; Pokluda, R. Reduced graphene oxide-based nanometal-composite containing copper and silver nanoparticles protect tomato and pepper against Xanthomonas euvesicatoria infection. Chem. Biol. Technol. Agric. 2022, 9, 1–16. [Google Scholar] [CrossRef]
- Bytešníková, Z.; Koláčková, M.; Dobešová, M.; Švec, P.; Ridošková, A.; Pekárková, J.; Přibyl, J.; Cápal, P.; Húska, D.; Adam, V. New insight into the biocompatibility/toxicity of graphene oxides and their reduced forms on Chlamydomonas reinhardtii. NanoImpact 2023, 31, 100468. [Google Scholar] [CrossRef]
- da Silva, R.S.; de Oliveira, M.M.G.; de Melo, J.O.; Blank, A.F.; Corrêa, C.B.; Scher, R.; Fernandes, R.P.M. Antimicrobial activity of Lippia gracilis essential oils on the plant pathogen Xanthomonas campestris pv. campestris and their effect on membrane integrity. Pestic. Biochem. Physiol. 2019, 160, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Standards, N.C.f.C.L.; Barry, A.L. Methods for Determining Bactericidal Activity of Antimicrobial Agents: Approved Guideline; National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 1999; Volume 19. [Google Scholar]
- Roberts, S.; Brough, J.; Hunter, P. Modelling the spread of Xanthomonas campestris pv. campestris in module-raised brassica transplants. Plant Pathol. 2007, 56, 391–401. [Google Scholar] [CrossRef]
- Peňázová, E.; Dvořák, M.; Ragasová, L.; Kiss, T.; Pečenka, J.; Čechová, J.; Eichmeier, A. Multiplex real-time PCR for the detection of Clavibacter michiganensis subsp. michiganensis, Pseudomonas syringae pv. tomato and pathogenic Xanthomonas species on tomato plants. PLoS ONE 2020, 15, e0227559. [Google Scholar] [CrossRef]
- Du Toft, L.; Crowe, F.; Derie, M.; Simmons, R.; Pelter, G. Bacterial blight of carrot seed crops in the Pacific Northwest. Phytopathology 2004, 94, S26. [Google Scholar] [CrossRef]
- Belvoir, T.; Scott, J.C.; Dung, J.K. Identification and screening of bacteria for the biocontrol of Xanthomonas hortorum pv. carotae in carrot seed crops. In Proceedings of the Plant Health 2019, APS Annual Meeting, St Paul, MN, USA, 3–7 August 2019. [Google Scholar]
- Clement, J.L.; Jarrett, P.S. Antibacterial silver. Met. Based Drugs 1994, 1, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Elamawi, R.M.; Al-Harbi, R.E. Effect of biosynthesized silver nanoparticles on Fusarium oxysporum fungus the cause of seed rot disease of faba bean, tomato and barley. J. Plant Prot. Pathol. 2014, 5, 225–237. [Google Scholar] [CrossRef]
- Jo, Y.-K.; Cromwell, W.; Jeong, H.-K.; Thorkelson, J.; Roh, J.-H.; Shin, D.-B. Use of silver nanoparticles for managing Gibberella fujikuroi on rice seedlings. Crop. Prot. 2015, 74, 65–69. [Google Scholar] [CrossRef]
- Tariq, M.; Mohammad, K.N.; Ahmed, B.; Siddiqui, M.A.; Lee, J. Biological synthesis of silver nanoparticles and prospects in plant disease management. Molecules 2022, 27, 4754. [Google Scholar] [CrossRef]
- Pariona, N.; Mtz-Enriquez, A.I.; Sánchez-Rangel, D.; Carrión, G.; Paraguay-Delgado, F.; Rosas-Saito, G. Green-synthesized copper nanoparticles as a potential antifungal against plant pathogens. RSC Adv. 2019, 9, 18835–18843. [Google Scholar] [CrossRef]
- Giannousi, K.; Avramidis, I.; Dendrinou-Samara, C. Synthesis, characterization and evaluation of copper based nanoparticles as agrochemicals against Phytophthora infestans. RSC Adv. 2013, 3, 21743–21752. [Google Scholar] [CrossRef]
- Chen, J.; Mao, S.; Xu, Z.; Ding, W. Various antibacterial mechanisms of biosynthesized copper oxide nanoparticles against soilborne Ralstonia solanacearum. RSC Adv. 2019, 9, 3788–3799. [Google Scholar] [CrossRef]
- Banik, S.; Pérez-de-Luque, A. In vitro effects of copper nanoparticles on plant pathogens, beneficial microbes and crop plants. Span. J. Agric. Res. 2017, 15, e1005. [Google Scholar] [CrossRef]
- Reddy, J.R.S.M.; Kannan, K.P.; Sankaran, K.; Rengasamy, G.; Priya, V.V.; Sathishkumar, P. Eradication of dental pathogens using flavonoid rutin mediated silver-selenium nanoparticles. Inorg. Chem. Commun. 2023, 157, 111391. [Google Scholar] [CrossRef]
- Mittal, A.K.; Thanki, K.; Jain, S.; Banerjee, U.C. Comparative studies of anticancer and antimicrobial potential of bioinspired silver and silver-selenium nanoparticles. J. Mater. NanoSci. 2016, 3, 22–27. [Google Scholar]
- Serov, D.A.; Khabatova, V.V.; Vodeneev, V.; Li, R.; Gudkov, S.V. A review of the antibacterial, fungicidal and antiviral properties of selenium nanoparticles. Materials 2023, 16, 5363. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Yahia, L.H.; Sacher, E. Antimicrobial properties of the Ag, Cu nanoparticle system. Biology 2021, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Miranda, R.M.d.; Dias, D.C.F.d.S.; Picoli, E.A.d.T.; Silva, P.P.d.; Nascimento, W.M. Physiological quality, anatomy and histochemistry during the development of carrot seeds (Daucus carota L.). Ciênc. Agrotecnologia 2017, 41, 169–180. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Salem, S.S.; Khalil, A.M.; El-Wakil, D.A.; Fouda, H.M.; Hashem, A.H. Potential of biosynthesized zinc oxide nanoparticles to control Fusarium wilt disease in eggplant (Solanum melongena) and promote plant growth. BioMetals 2022, 35, 601–616. [Google Scholar] [CrossRef]
- Varympopi, A.; Dimopoulou, A.; Papafotis, D.; Avramidis, P.; Sarris, I.; Karamanidou, T.; Kerou, A.K.; Vlachou, A.; Vellis, E.; Giannopoulos, A. Antibacterial activity of copper nanoparticles against Xanthomonas campestris pv. vesicatoria in tomato plants. Int. J. Mol. Sci. 2022, 23, 4080. [Google Scholar] [CrossRef]
- Govindan, R.; Chackaravarthi, G.; Ramachandran, G.; Chelliah, C.K.; Muthuchamy, M.; Quero, F.; Mothana, R.A.; Noman, O.M.; Siddiqui, N.A.; Li, W.-J. Effective removal of biofilm formation in Acinetobacter baumannii using chitosan nanoparticles loaded plant essential oils. J. King Saud Univ. Sci. 2022, 34, 101845. [Google Scholar] [CrossRef]
- Yang, W.; Fortunati, E.; Gao, D.; Balestra, G.M.; Giovanale, G.; He, X.; Torre, L.; Kenny, J.M.; Puglia, D. Valorization of acid isolated high yield lignin nanoparticles as innovative antioxidant/antimicrobial organic materials. ACS Sustain. Chem. Eng. 2018, 6, 3502–3514. [Google Scholar] [CrossRef]
- Bayat, M.; Zargar, M.; Chudinova, E.; Astarkhanova, T.; Pakina, E. In vitro evaluation of antibacterial and antifungal activity of biogenic silver and copper nanoparticles: The first report of applying biogenic nanoparticles against Pilidium concavum and Pestalotia sp. fungi. Molecules 2021, 26, 5402. [Google Scholar] [CrossRef]
- Park, S.; Ahn, Y.-J. Multi-walled carbon nanotubes and silver nanoparticles differentially affect seed germination, chlorophyll content, and hydrogen peroxide accumulation in carrot (Daucus carota L.). Biocatal. Agric. Biotechnol. 2016, 8, 257–262. [Google Scholar] [CrossRef]
- Bayat, M.; Zargar, M.; Murtazova, K.M.-S.; Nakhaev, M.R.; Shkurkin, S.I. Ameliorating seed germination and seedling growth of nano-primed wheat and flax seeds using seven biogenic metal-based nanoparticles. Agronomy 2022, 12, 811. [Google Scholar] [CrossRef]
- Kadri, O.; Karmous, I.; Kharbech, O.; Arfaoui, H.; Chaoui, A. Cu and CuO nanoparticles affected the germination and the growth of barley (Hordeum vulgare L.) seedling. Bull. Environ. Contam. Toxicol. 2022, 108, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Essa, H.L.; Abdelfattah, M.S.; Marzouk, A.S.; Shedeed, Z.; Guirguis, H.A.; El-Sayed, M.M. Biogenic copper nanoparticles from Avicennia marina leaves: Impact on seed germination, detoxification enzymes, chlorophyll content and uptake by wheat seedlings. PLoS ONE 2021, 16, e0249764. [Google Scholar] [CrossRef] [PubMed]
- Santás-Miguel, V.; Arias-Estévez, M.; Rodríguez-Seijo, A.; Arenas-Lago, D. Use of metal nanoparticles in agriculture. A review on the effects on plant germination. Environ. Pollut. 2023, 334, 122222. [Google Scholar] [CrossRef]
- Guo, H.; Liu, Y.; Chen, J.; Zhu, Y.; Zhang, Z. The effects of several metal nanoparticles on seed germination and seedling growth: A meta-analysis. Coatings 2022, 12, 183. [Google Scholar] [CrossRef]
- Gilbertson, R. Bacterial Leaf Blight of Carrot; Compendium of Umbelliferous Crop Diseases; American Phytopathological Society: St. Paul, MN, USA, 2002; pp. 11–12. [Google Scholar]
- Umesh, K.; Davis, R.; Gilbertson, R. Seed contamination thresholds for development of carrot bacterial blight caused by Xanthomonas campestris pv. carotae. Plant Dis. 1998, 82, 1271–1275. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Nanomaterial | Size | Shape | Concentration of Stock Solution (mg L−1) | Dilution Factor of Stock Solution |
|---|---|---|---|---|
| AgNPs_29 [11] | 2 nm | Spherical | 100 Ag | 10×–100× ** |
| AgNPs_30 [11] | 22 nm | Spherical | 100 Ag | 2×–8× * 10×–100× |
| AgSeNPs_8 [30] | 60 nm | Cluster plates | 3703 Ag 748 Se | 10×–100× |
| 200×–1000× *** | ||||
| CuNPs_50 [30] | 10 nm | Spherical | 545 Cu | 2×–8× |
| CuNPs_53 [30] | 100 nm | Spherical | 3974 Cu | 10×–100× |
| rGO-Cu_25 [31] | Flakes, 1×–10 nm thickness | Plate | 1694 Cu | 2×–8× |
| rGO [32] | Flakes, 1×–10 nm thickness | Plate | 3100 rGO | 2×–8× |
| SeNPs_40 [30] | 150 nm | Spherical | 517 Se | 2×–8× |
| Nanomaterial | Df | Concentration (mg L−1) | |||
|---|---|---|---|---|---|
| AgNPs_29 | 10× | 10 | Ag | ||
| 8× | 13 | ||||
| 4× | 25 | ||||
| CuNPs_53 | 10× | 397 | Cu | ||
| 8× | 497 | ||||
| 4× | 993 | ||||
| AgSeNPs_8 | 70× | 53 | Ag | 11 | Se |
| 60× | 62 | 13 | |||
| 50× | 74 | 15 | |||
| Nanomaterial | MIC (mg L−1) | MBC (mg L−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Df | Ag | Cu | Se | Df | Ag | Cu | Se | |
| AgNPs_29 | 40× | 3 | - | - | 10× | 10 | - | - |
| AgNPs_30 | 40× | 3 | - | - | NE | - | - | - |
| AgSeNPs_8 | 200× | 19 | - | 4 | 70× | 53 | - | 11 |
| CuNPs_50 | NE | - | - | - | NE | - | - | - |
| CuNPs_53 | 20× | - | 199 | - | 10× | - | 397 | - |
| rGO-Cu_25 | 10× | - | 169 | - | 2× | - | 847 | - |
| rGO | NE | - | - | - | NE | - | - | - |
| SeNPs_40 | NE | - | - | - | NE | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wohlmuth, J.; Tekielska, D.; Hakalová, E.; Čechová, J.; Bytešníková, Z.; Richtera, L.; Baránek, M. Eliminating the Pathogen Xanthomonas hortorum pv. carotae from Carrot Seeds Using Different Types of Nanoparticles. Agriculture 2024, 14, 498. https://doi.org/10.3390/agriculture14030498
Wohlmuth J, Tekielska D, Hakalová E, Čechová J, Bytešníková Z, Richtera L, Baránek M. Eliminating the Pathogen Xanthomonas hortorum pv. carotae from Carrot Seeds Using Different Types of Nanoparticles. Agriculture. 2024; 14(3):498. https://doi.org/10.3390/agriculture14030498
Chicago/Turabian StyleWohlmuth, Jan, Dorota Tekielska, Eliška Hakalová, Jana Čechová, Zuzana Bytešníková, Lukáš Richtera, and Miroslav Baránek. 2024. "Eliminating the Pathogen Xanthomonas hortorum pv. carotae from Carrot Seeds Using Different Types of Nanoparticles" Agriculture 14, no. 3: 498. https://doi.org/10.3390/agriculture14030498