
A Genome-Wide Comparative Analysis of AUX1/LAX, PIN, and ABCB Genes Reveals Their Roles in Cucumber Fruit Curving
1
College of Advanced Agriculture and Ecological Environment, Heilongjiang University, Harbin 150080, China
2
Key Laboratory of Sugar Beet Genetic Breeding, Heilongjiang University, Harbin 150080, China
3
Key Laboratory of Biology and Genetic Improvement of Horticultural Crops (Northeast Region), Ministry of Agriculture, College of Horticulture and Landscape Architecture, Northeast Agricultural University, Harbin 150030, China
*
Author to whom correspondence should be addressed.
Agriculture 2024, 14(5), 657; https://doi.org/10.3390/agriculture14050657
Submission received: 2 March 2024
/
Revised: 16 April 2024
/
Accepted: 19 April 2024
/
Published: 24 April 2024
(This article belongs to the Special Issue Agronomic Practices for Improving Growth, Quality, and Yield of Vegetables)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Auxin transport is regulated by the AUX1/LAX, PIN, and ABCB gene families, controlling the distribution of auxin and ultimately fruit curving in cucumbers. However, studies on the differential expression of these auxin transporters and their roles in fruit curving are limited. In this study, we identified 36 auxin transporters from cucumber, including CsLAX1–7, CsPIN1–10, and CsABCB1–19. Basic characteristic analysis revealed that all CsLAX proteins were conservative, and a C-terminal NPNTY motif was found in CsPIN1–4/7–10. CsABCB1/5/11/14/17 were categorized as half-size transporters. Phylogenetic analysis revealed a genetic relationship between auxin transporters in Arabidopsis and cucumber. Exogenous auxin treatment on fruits and qPCR analysis indicated that differential expression patterns of auxin transporters control cucumber fruit curving. Co-expression analysis indicated that CsPIN1 and CsLAX2 were substantially negatively correlated, and they displayed opposite expression patterns in curved fruits. A proposed model suggested that CsLAX2 transports extracellular auxin to the convex side of the fruit; however, CsPIN1 inhibits auxin efflux at the same location. This leads to uneven auxin distribution that results in cucumber fruit curving.
1. Introduction
Asymmetric distribution of auxin is crucial for regulating cell differential growth and controlling tissue curving mechanisms in plants, including phototropism [1], gravitropism [2], and apical hook development [3]. Proteins in the plasma membrane (PM) are responsible for auxin transportation between cells [4]. These proteins belong to the three following separate gene families: auxin resistant (AUX) 1/like AUX1 (LAX) influx carriers, pin-formed (PIN) efflux carriers, and ATP-binding cassette B/multidrug-resistance/P-glycoprotein (ABCB/MDR/PGP) efflux/condition transporters [5]. Polar transport involves the active transportation of auxin molecules in the plant organs, which is influenced by the uneven distribution of AUX1/LAX, PIN, and ABCB proteins [6]. Therefore, comparative analysis of these auxin transporters can help in understanding the potential mechanisms of asymmetric growth in plants.
AUX1/LAX proteins belong to the amino acid/auxin permease family that promotes the absorption and movement of auxin in cells [7]. In Arabidopsis thaliana, the protein products of four conservative genes (AUX1, LAX1, LAX2, and LAX3) resemble to amino acid transporters. AtAUX1 and AtLAX3 exhibit separate functions based on their spatial expression patterns. AtAUX1 promotes entry of auxin to the inner side of the apical hook, whereas AtLAX3 removes auxin towards the root [8]. Both AtAUX1 and AtLAX2 rely on AXR4 to transport auxin across the membranes of cells involved in root growth [9]. AtAUX1 protein exhibits varying expression levels on the PM of the outer integument cells and across the entire endosperm, and it regulates seed size via modulating auxin distribution [10]. Despite numerous reports on the properties and functions of AUX1/LAX, they are frequently overlooked because of the restricted number of family members.
The PIN family members are unevenly distributed on the PM, and their placement dictates the direction of auxin transport [11]. The transporters are classified as typical long PIN proteins (PIN1–PIN4 and PIN7) and short proteins (PIN5 and PIN8) [12]. The long PIN proteins are situated on the PM, whereas the shorter proteins PIN5 and PIN8 are found in the endoplasmic reticulum. PIN6 is present in the PM and endoplasmic reticulum [13]. The five long PIN proteins reestablish the uneven auxin distribution in the root tip of Atpin1/3/4/7 mutants; however, PIN5 and PIN6 proteins do not possess this ability [14]. PIN3, PIN4, and PIN7 have overlapping roles in apical hook development by regulating auxin asymmetric distribution in epidermal cells [8,15]. OsPIN1b is involved in auxin transport to regulate plant morphology and root gravity in rice [16]. Naptalam (NPA) specifically targets PINs to inhibit auxin transport, leading to a change in the open conformation of the PIN1 and PIN8 protein structures [17,18]. Therefore, comparative analysis of PINs contributes to understanding their gene function.
The ABCB gene family is the second largest subfamily of the ABC transporters, primarily situated symmetrically on the PM to aid in either the intake or release of auxin [19]. Proteins are categorized as full-size-type transporters or half-size transporters based on the quantity of transmembrane domains (TMDs) and nucleotide binding domains [20]. The distinctive configuration improves the auxin transportation capacity of different ABCB proteins [21]. It has been identified that ABCB1/4/14/19/21 is involved in the polar transport of auxin [11]. AtABCB4, found in the roots, controls the transport of auxin from the root apex to the epidermis toward the shoot and restricts the elongation of root hair by regulating the efflux of auxin [22,23]. GFP-ABCB19 is mostly found on the inner side of the hook, and the suppression of hypocotyl elongation by phytochrome is due to decreased ABCB19-dependent auxin transport [24,25]. AtABCB6 and AtABCB20 controlled the growth of shoots and roots [26,27]. ABCB15–18 and ABCB22 participate in the transport of auxin from the lateral root cap and epidermis toward the shoot [28].
Fruit curvature of cucumber (Cucumis sativus L.) directly impacts appearance quality and leads to economic losses for farmers. Auxin transport regulates fruit curving via unequal distribution of auxin [29]. However, the potential interactions of auxin transporters in cucumber are unknown. In this study, we aimed to comparatively analyze the gene structure, conserved domains, and cis-regulatory elements in the promoter regions of AUX1/LAX, PIN, and ABCB family genes in cucumber. Furthermore, the co-expression network and gene expression analysis were performed to explore the role of auxin transporters in cucumber fruit curving. This study provided a basis for studying the regulation of auxin transport in cucumber fruit curving.
2. Materials and Methods
2.1. Plant Materials and Treatment
The seeds of “changchunmici” cucumber with a fruit curving ratio of 52.56 were grown under 16 h light (at 29 °C)/8 h dark (at 17 °C) conditions in a greenhouse at Northeast Agricultural University, China. The exocarps from curved and straight fruits were collected at 2, 4, 6, 8, and 10 days post-anthesis (DPA) for RT-qPCR analysis.
Experiments with exogenous hormones on cucumber ovaries were performed at 0 DPA. On one side of the ovaries of straight fruits, 30 µM of synthesized indole-3-acetic acid (IAA), 50 µM of efflux inhibitor N-1-naphtylphthalamic acid (NPA), and 0.1 µM of influx inhibitor 2-naphthoxyacetic acid (NOA) were sprayed [30]. For the curved fruits, the three following treatments were set: spraying 30 µM IAA on the concave side, 50 µM NPA on the convex side, or 0.1 µM NOA on the convex side. The fruits treated by ddH2O were set as the control. The exocarps were collected at 4 DPA to determine auxin content and extract RNA. Three independent biological replicates were set for each treatment. Auxin was extracted as per the method described by Weiler [31], and its concentration was determined using a Plant IAA ELISA kit (Meimian Industrial Co., Ltd., Yancheng, Jiangsu, China). Statistical analysis was performed using one-way ANOVA. p value < 0.05 was considered significant.
2.2. Characterization and Phylogenetic Analysis of Auxin Transporters
To identify the auxin transporters in cucumber, the v2 whole genome protein sequences of cucumber were obtained from the Cucurbit Genome Database (CuGenDB, http://cucurbitgenomics.org/, accessed on 22 August 2023). The Hidden Markov Models (HMM) profiles were analyzed to identify the potential auxin transporters in cucumber, including AUX1/LAX (Pfam 01490), PIN (Pfam 03574), and ABCB (Pfam 00005 and Pfam 00664). Additionally, the protein sequences of Arabidopsis auxin transporter from TAIR (https://www.arabidopsis.org/, accessed on 22 August 2023) were used as queries in BLASTp searches against the cucumber genome (Table S1), with an e-value of ≤10−5 and bit scores of >100. Further, redundant protein sequences were eliminated using the InterPro database (https://www.ebi.ac.uk/interpro/, accessed on 22 August 2023) and SMART (http://smart.embl-heidelberg.de/, accessed on 22 August 2023).
Protein length (aa), molecular weight (kDa), and isoelectric point (pI) were calculated using Pepstats (https://www.ebi.ac.uk/Tools/seqstats/emboss_pepstats/, accessed on 30 August 2023). The WoLF PSORT program was used to predict the subcellular localization of proteins (https://wolfpsort.hgc.jp/, accessed on 18 September 2023) [32]. Multiple sequence alignment analysis was performed using the MUSCLE program, with a gap open value of −2.90 and a gap extension value of 0.00. A phylogenetic tree was constructed in MEGA 11.0 using the neighbor-joining (NJ) tree method with 1000 replicate bootstraps [33] and was visualized using Evolview v2 [34].
2.3. Gene Structure, Genome Distribution, and Duplication Analysis
Conserved motifs of AUX1/LAX, PIN, and ABCB proteins in cucumber were explored using MEME online software version 5.5.4 (https://meme-suite.org/meme/doc/meme.html, accessed on 27 September 2023) and visualized using TBtools version 2.069 [35,36]. The gene structures of the AUX1/LAX, PIN, and ABCB gene families were predicted using TBtools version 2.069 [31]. The TMDs were analyzed using DeepTMHMM version 1.0.24 (https://dtu.biolib.com/DeepTMHMM, accessed on 18 September 2023) [37].
The duplicate gene datasets were obtained from Plant Repeat Gene Database (http://pdgd.njau.edu.cn:8080/, accessed on 19 February 2023), including whole genome duplications (WGD), tandem duplications, transposed duplications (TRD), dispersed duplications (DSD), and proximal duplication (PD). The collinear blocks between cucumber and Arabidopsis were analyzed using MCScanX, with the default parameters of TBtools version 2.069 [38]. The gene duplication and synteny were visualized using TBtools version 2.069 [36].
2.4. Analysis of Cis-Acting Elements of Auxin Transporters
The upstream regions (1500 bp) before the initiation codon (ATG) of the AUX1/LAX, PIN, and ABCB genes of cucumber were obtained from cucumber genome database using TBtools software version 2.069 [36]. These promoter sequences were updated on the PlantCARE website (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 27 March 2024) for searching cis-acting elements. The enrichment of cis-acting elements was analyzed using tidyverse tool and ggplot2 tool in R v4.2.1.
2.5. Co-Expression Analysis of Auxin Transporters
The published transcriptome data by Wang et al. [39] was obtained from the Sequence Read Archive database (accession number: SRP111902), which related to the curved and straight fruits of cucumber at 2 DPA. The transcription factors (TFs) were screened from the above transcriptome data using the PlanTFDB database (http://planttfdb.cbi.pku.edu.cn/, accessed on 30 October 2023). The Pearson correlations between TFs and auxin transporters were calculated using the corrplot tool in R v4.2.1, using readings per kilobase pair per million reads (RPKM > 1). Further, the gene pairs with an absolute correlation coefficient > 0.97 were used to construct co-expression networks. Cytoscape software version 3.10.1 was used for the visualization [40].
2.6. Real-Time Quantitative PCR(RT-qPCR) Analysis
Total RNA was extracted from the exocarps at 2, 4, 6, 8, and 10 DPA using RNA-easy Isolation Reagent (Vazyme, Nanjing, China). The first strand of cDNA was synthesized using the Prime ScriptTM RT reagent Kit with gDNA Eraser (Takara, Shiga, Japan). RT-qPCR analysis was conducted in the Quantagene q225 system (KUBO, Beijing, China) using TB Green Premix (Takara, Shiga, Japan). The amplification program was set as follows: preheating at 95.0 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 35 s. The specific primers are given in Table S2. Relative gene expression was determined using the 2−ΔΔCT method. Elongation factor 1 alpha (CsEF1α) served as the reference gene [29]. Gene expression levels were visualized with clustering heatmaps using TBtools software (version 2.069) [36]. All experiments were conducted with three biological replicates.
3. Results
3.1. Auxin Transport Contributed to Asymmetric Auxin Distribution and Resulted in Curved Fruit
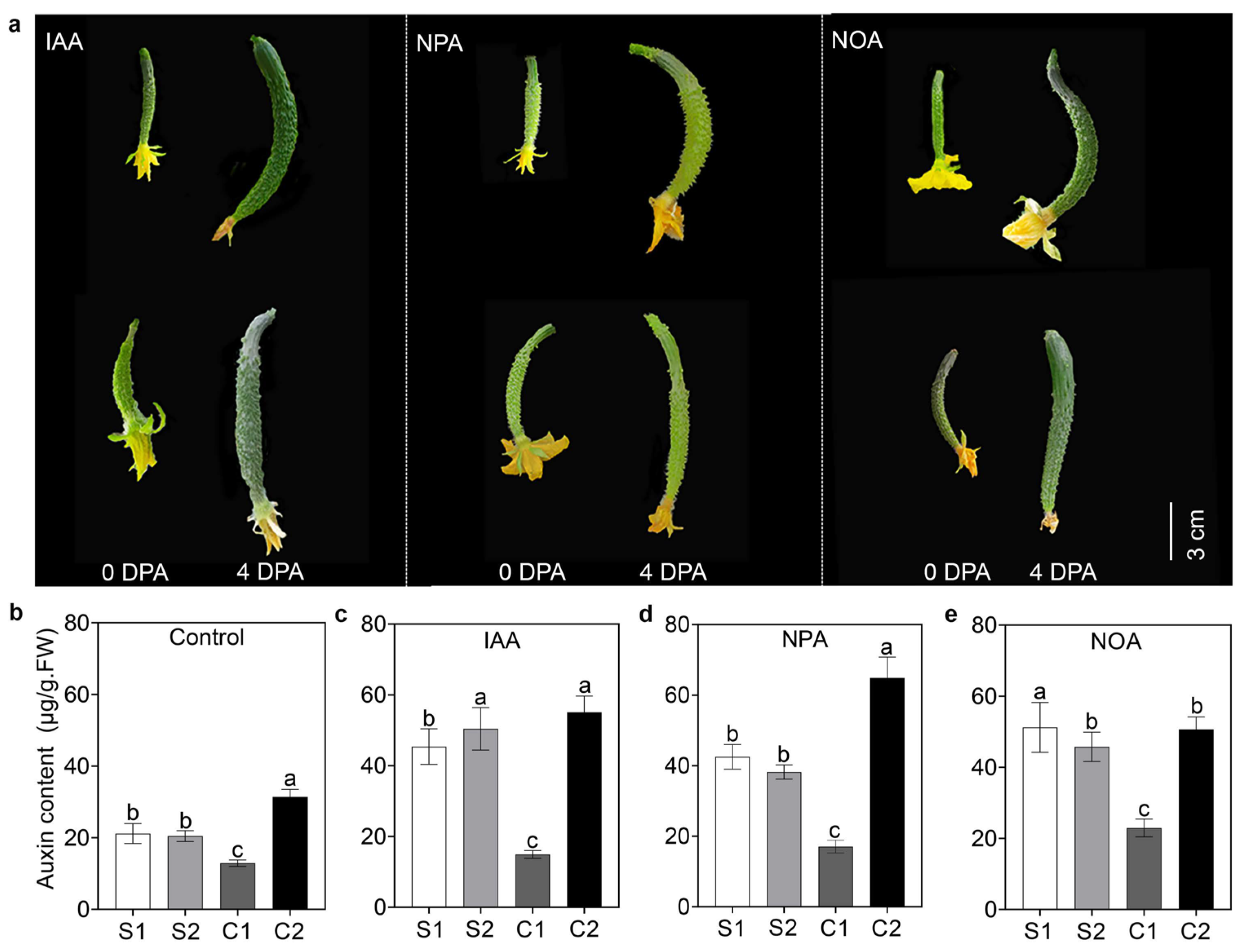
To determine the impact of auxin transport on fruit curvature, we sprayed exogenous auxin (IAA) and auxin transport inhibitors (NOA and NPA) on both straight and curved fruits. Phenotype analysis revealed that spraying IAA, NPA, and NOA on one side of the ovary of straight fruits (0 DPA) led to formation of curved shape at 4 DPA, whereas IAA treatment on the concave side of the ovary of curved fruits led to the development of straight fruit. When NOA and NPA were sprayed on the convex side of the ovaries of curved fruits, straight fruits were produced at 4 DPA (Figure 1a). Further analysis revealed that auxin content was consistent on both sides of the straight fruits, whereas it was higher on the convex side than on the concave side in curved fruits (Figure 1b). Regardless of the total amount of auxin, any unequal distribution of auxin led to fruit curving (Figure 1c–e). These results revealed that auxin transport contributes to the uneven distribution of auxin in cucumber fruit, resulting in fruit curving.
3.2. Genome-Wide Identification of Auxin Transporters
Auxin-transport-related genes, including seven LAX, 10 PIN, and 19 ABCB genes, were identified in cucumber and named according to their location on the chromosomes (CsLAX1–7, CsPIN1–10, and CsABCB1–19). The sizes of CsLAX proteins ranged from 466 to 494 amino acids (aa), with molecular weights ranging from 52.51 to 55.54 kDa and predicted isoelectric points (pIs) ranging from 8.2269 to 9.3645. For CsPIN proteins, protein size ranged from 103 to 645 aa, molecular weights ranged from 10.98 to 70.79 kDa, and the estimated pIs ranged from 4.5119 to 10.0279. For CsABCBs, molecular weights ranged from 34.39 to 160.12 kDa, pIs ranged from 6.1173 to 9.7729, and protein size ranged from 305 to 1464 aa. CsPIN4 was predicted to be present in the vacuolar membrane, CsPIN6, CsABCB5, and CsABCB17 were predicted to be present in the chloroplast and the others in the PM. All CsLAX proteins have 11 transmembrane helices. Except for CsPIN5 and CsPIN6, most CsPIN have 10 transmembrane helices. TMD for CsABCB proteins ranged from 3 to 14 (Table S3).
3.3. Distribution and Duplication Analysis of Auxin Transporters
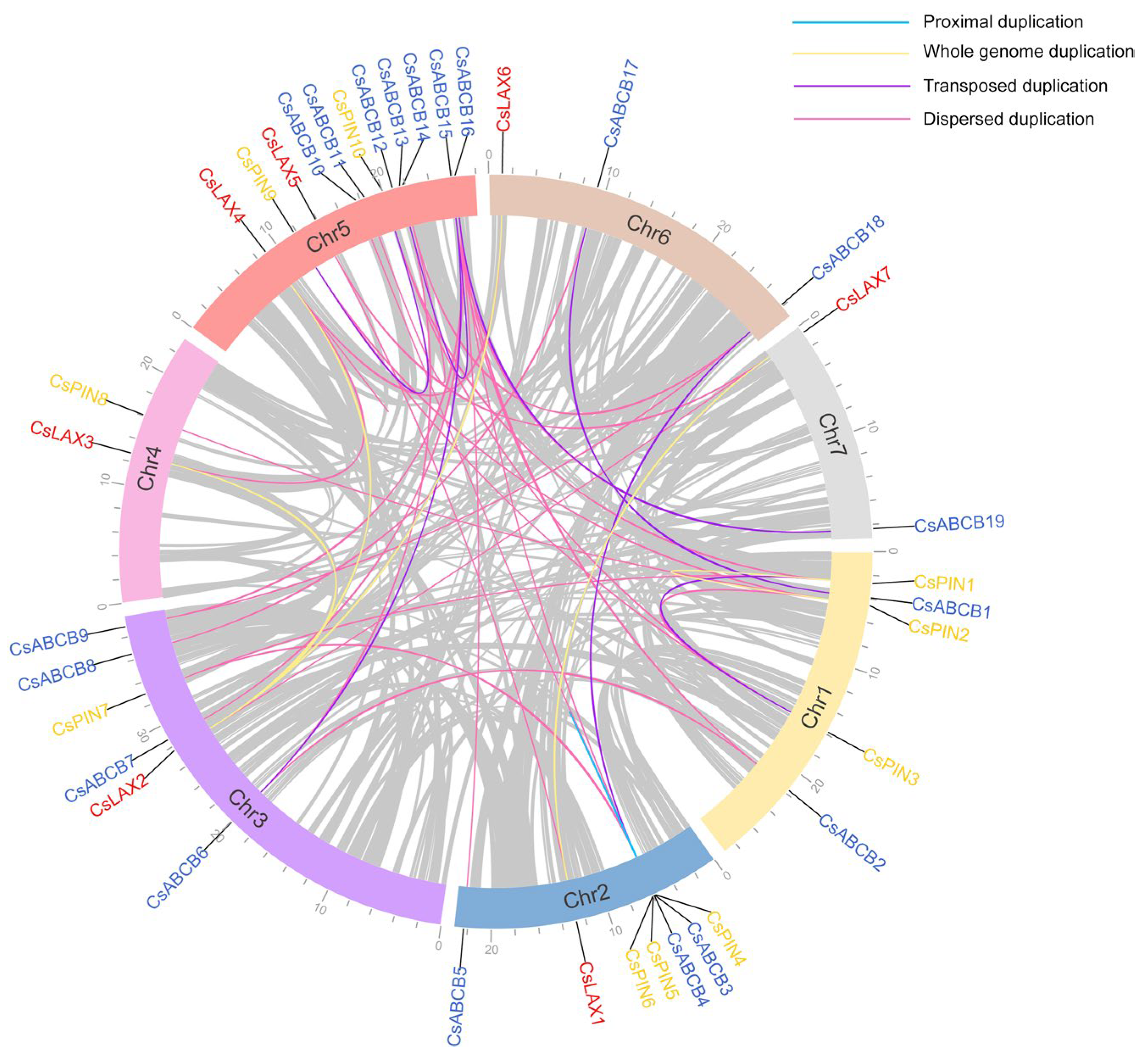
In total, 36 auxin transporters were irregularly distributed on seven chromosomes in cucumber. CsLAXs were identified on all chromosomes except chromosome 1. CsPINs were found on chromosomes 1, 2, 3, 4, and 5. No CsABCB gene was found on chromosome 4. The following four types of duplications were found in cucumber: five pairs of WGD, seven pairs of DNA-TRD, 29 pairs of DSD, and one pair of PD (Figure 2). The CsLAX genes were copied 40% via WGD and 60% via scattered duplication. DNA-TRD and DSD were significant factors in the evolution of the CsPIN and CsABCB families.
3.4. Analyses of Phylogenetic Relationships, Synteny Relationships, Conservative Motif, and Gene Structure
An unrooted NJ phylogenetic tree was constructed with a total of 76 auxin transporters, including four AtAUX/LAXs, eight AtPINs, and 28 AtABCB proteins from Arabidopsis and seven CsLAXs, 10 CsPINs, and 19 CsABCBs from cucumber. A phylogenetic tree of LAX proteins identified the following two subfamilies: I and II (Figure 3a). CsLAX5 and CsLAX7 proteins exhibited 98% and 99% amino acid similarity with the closely related AtLAX3 and AtLAX2, respectively. The phylogenetic tree of the PIN proteins displayed two primary subgroups (I and II) (Figure 3b). Subfamily I consisted of six CsPINs. AtPIN2 and CsPIN3 were categorized together based on their 99% amino acid similarity. In subfamily II, AtPIN5 and AtPIN8 were clustered with CsPIN4 and CsPIN7 due to their closely related sequences. Phylogenetic analysis of ABCB proteins revealed the following four subfamilies: I, II, III, and IV (Figure 3c). Subfamilies I and II both consisted of five cucumber ABCB proteins. Subfamily I consisted of the proteins CsABCB1 and CsABCB14, which exhibited 100% amino acid identity with AtABCB27 and AtABCB26, respectively. In subfamily II, CsABCB16 and AtABCB19, along with CsABCB7 and AtABCB1, were grouped together in a branch with 100% amino acid identity. Subfamilies III and IV contained five and four CsABCB proteins, respectively. Arabidopsis ABCB proteins and cucumber ABCB proteins were not classified in subfamily III together.
In synteny analysis (Figure 3d), 20 orthologous gene pairs were identified among the three auxin transporter families of cucumber and Arabidopsis. Among them, eight pairs of syntenic orthologous genes (one-to-one) were identified, such as AtLAX3–CsLAX5, AtPIN2–CsPIN3, and AtABCB1–CsABCB7. These gene pairs might be derived from a common ancestor. Three AtLAXs and one AtPIN had a one-to-two corresponding relationship with CsLAXs and CsPINs, respectively. Two pairs were two AtABCB genes corresponding to the same CsABCB, such as AtABCB7 and AtABCB9–CsABCB18.
Each CsLAX protein contained 10 motifs because of the 80.8% sequence similarity (Figure 4). CsPIN1–4/7–10 had the C-terminal NPNTY motif, known as motif 8, which is a conserved area in the HL domain of the PINs. CsABCB1, 5, 11, 14, and 17 had three–eight motifs and were categorized as half-size transporters (Table S4). The CsLAX genes had seven to eight exons. The number of exons in CsPINs varied between five and seven, except for CsPIN5 and CsPIN6. Subfamily I exhibited a more conservative gene structure compared with subfamily II. The exon–intron architecture exhibited considerable differences, with the number of exons varying from two to 18. The introns of CsABCB11, CsABCB14, and CsABCB17 were more in number than those of other subfamily members.
3.5. Analysis of Cis-Regulatory Elements in the Promoter Regions of Auxin Transporters
A total of 845 cis-regulatory elements were obtained in the promoter regions (1500 bp) of auxin transporters. Based on their biological functions, these cis-regulatory elements were divided into the following five functional groups: plant growth and development, light response, plant hormone, stress response, and TFs (Figure 5). Particularly, 48 auxin cis-regulatory elements, such as AuxRR-core, as-1, TGA-box, and TGA-element, were identified from auxin transporters and were essential for auxin signaling and transport. The TGA-element is known to be an auxin-responsive element, whereas the TGA-box is a part of the auxin-responsive element. Furthermore, most auxin transporters had both MYB- and MYC-responsive regions, suggesting that MYB and MYC TFs may regulate the expression of these genes.
3.6. Differential Expression of Auxin Transporters Control Fruit Curving
Using qPCR data, a gene expression heatmap was constructed to investigate the role of auxin transporters. The criteria for selecting differentially expressed genes were mean >1 and minimum absolute fold change |log2FC| > 1. Expression patterns of auxin transporters were different between straight and curved fruits. Curved fruits were produced after IAA treatment, which led to increased expression of CsLAX2–7, CsPIN1–2/7/10, and CsABCB1/3/5–7/13/17 on the convex side. However, expressions of these genes were not different on both sides of straight fruits (Figure 6a). LAX1/2/5, PIN1–2/8–10, and ABCB2–4/8/11–13/16/19 were differentially expressed on the concave and convex sides of curved fruits, whereas their expressions were similar on both sides of straight fruits (Figure 6b). These results indicated that differential expression patterns of auxin transporters control fruit curving. Clustering analysis further revealed that CsPIN1–2/9–10 and CsABCB6–7/9–12/14–17 were grouped together (Figure 6b), indicating they have similar functions in fruit curving.
3.7. Potential Roles of Auxin Transporters in Fruit Curving
Based on the published transcriptome data by Wang et al. [39], a co-expression network was constructed using the expression levels of auxin transporters. A total of 14 auxin transporters were significantly correlated with the six guide genes, including CsLAX1 (its expression level was the same as that of CsLAX2 reported by Li et al. [29]) (1), CsLAX2 (2), CsPIN1 (1), CsABCB12 (4), CsABCB16 (4), and CsABCB18 (2) (Figure 7a and Table S5). Among them, CsPIN3 was negatively correlated with both CsABCB12 (r = −0.999) and CsABCB16 (r = −0.994), whereas CsPIN10 was positively correlated with CsABCB16 (r = 0.992) and CsABCB18 (r = 0.999). CsPIN1 was significantly negatively correlated with CsLAX2 (r = −0.999), indicating that they might play opposite roles in fruit curving. Furthermore, CsLAX2 and CsPIN1 exhibited opposite expression patterns, particularly in the concave and convex sides of curved fruits at 4 DPA (Figure 7b). This result was supported by the phenotype of fruits after exogenous auxin treatment. Additionally, during fruit curving, the expression pattern of CsPIN1 was opposite to that of CsABCB16 and CsABCB18.
3.8. Co-Expression Analysis of TFs and Transporters
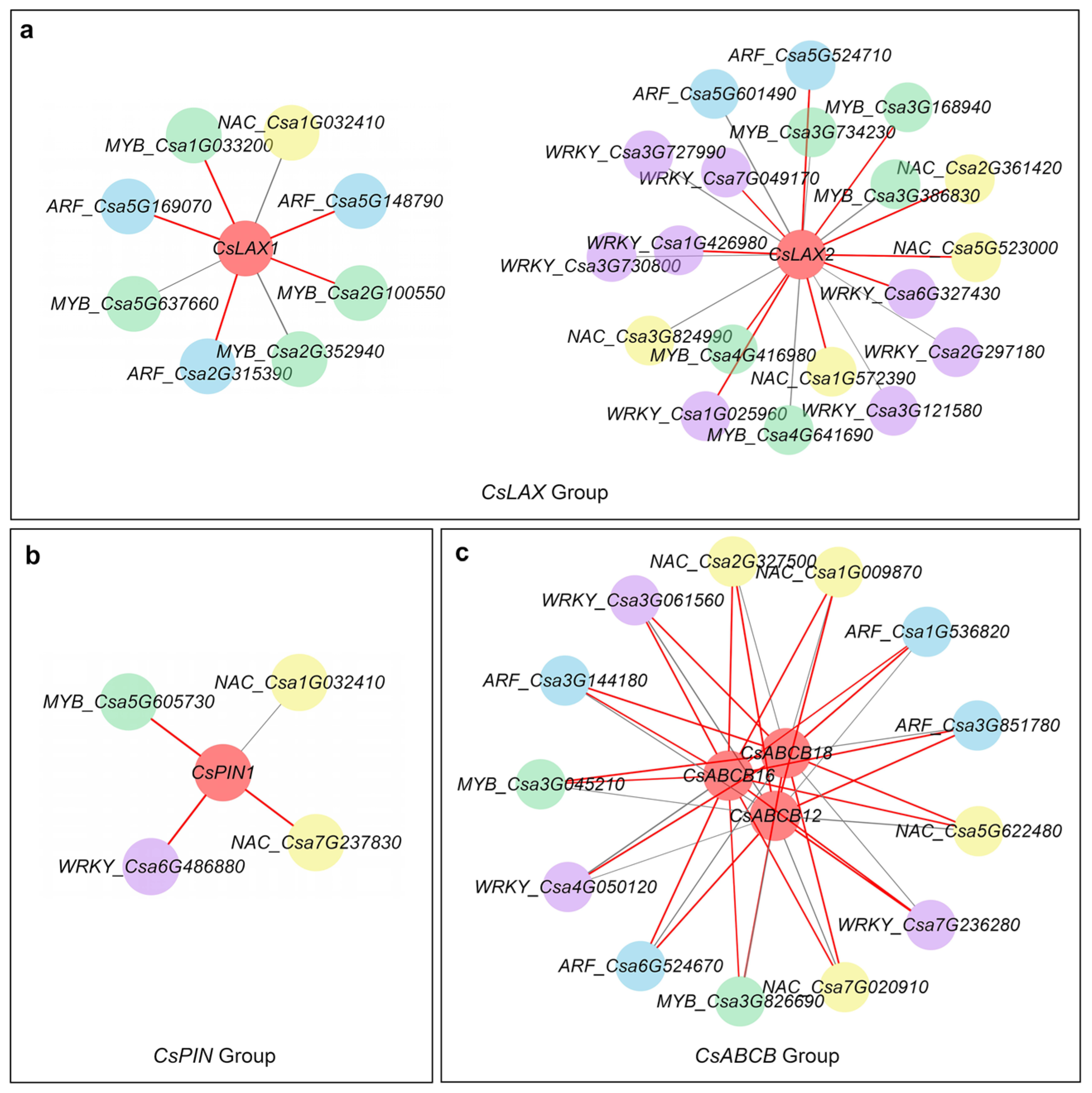
In total, 296 TFs were obtained from the published transcriptome data, among which the most abundant TFs were MYB, WRKY, NAC, and ARF. A co-expression network was constructed using TFs and guide genes. In the LAX group, five TFs, including ARF (3) and MYB (2) (r > 0.99), were strongly correlated with CsLAX1; however, 10 TFs were positively correlated with CsLAX2 (Figure 8a). MYB TFs were significantly correlated with CsPIN1 (Figure 8b). For the ABCB group, 13 TFs were typically related to CsABCB12/16/18 (Figure 8c). These TFs might regulate auxin transporters in fruit curving.
4. Discussion
4.1. Comprehensive Analysis of Auxin Transporters
The analysis of gene structure and motifs contributed to the elucidation of gene function. The results revealed that CsLAXs, like AtLAXs, had a highly conserved motif and gene structure and consisted of 11 transmembrane helices [13]. The PIN protein in Arabidopsis contained two closely conserved transmembrane helices at the N and C termini, linked by a cytoplasmic loop in the center [41]. Transmembrane domain analysis revealed that CsPIN4 and CsPIN7 had short central hydrophilic loops that were clustered with AtPIN5 and AtPIN8, whereas CsPIN1–3 and CsPIN8–10 had long central hydrophilic loops. Previous studies have reported the essential function of the NPNTY motif in facilitating interactions between membrane proteins and receptor proteins [42]. The motif was found to be conserved in cucumber (motif 8), except for CsPIN5 and CsPIN6. ABCB proteins were categorized into the following two classes: half- and full-size proteins. The full-size CcABCBs in Chinese hickory have a higher number of transmembrane regions than the half-size CcABCBs [21]. The same outcome was observed in cucumber (Table S3). These results suggested that the functions of LAX, PIN, and ABCB proteins were similar in cucumber and Arabidopsis.
4.2. Potential Roles of Auxin Transporters
The polar transit of auxin leads to an unequal distribution of AUX/LAX and PIN proteins in cells and tissues. This regulation is essential for conducting the roles of auxin in governing various plant developmental processes [5]. AUX/LAX and PIN proteins typically have contrasting roles in auxin accumulation. AtAUX1 and AtPIN2 exhibited opposite expression patterns in regulating auxin levels for root hair elongation under phosphorus stress [43]. Interestingly, AtAUX1 and AtPIN2 facilitated the transport of auxin to the root hair meristem for elongation growth in response to low nitrogen stress [44]. CsPIN1 and CsPIN7 regulated cucumber fruit length by inhibiting their own expression, leading to decreased auxin levels [45]. The unequal distribution of auxin in the ionospheric zone controlled pedicel abscission in tomatoes by downregulating AUX/LAXs and PINs [46]. AUX1/LAX and PIN proteins are involved in plant growth and development, and their impacts can be synergistic or antagonistic.
ABCBs independently play a role in root geotropism, and together with PINs, they control auxin transport. AtABCB19 interacted with AtPIN1 by forming homopolymers and heteropolymers to determine auxin transport [47]. Both clustering and co-expression analyses revealed that CsPIN10 were positively correlated with CsABCB16 and CsABCB18 (Figure 6 and Figure 7), indicating their collaborative participation in auxin transport for fruit curving. However, CsPIN1 and CsPIN3 were negatively correlated with both CsABCB12 and CsABCB16 (Figure 7). CsPIN1 inhibited the outflow of auxin, whereas CsABCB16 and CsABCB18 might promote the export of auxin to rebuild the auxin balance on both sides of the fruit. This result was consistent with a previous study reporting that ABCBs are independently involved in root geotropism, and together with PINs, they regulate auxin transport [48].
The involvement of auxin in plant growth and development is controlled by signal transduction. TFs play a role in binding intracellular and extracellular auxin signals. ARFs control the tissue-specific expression of PIN, whereas downstream WRKY23 is involved in regulating auxin feedback on PIN polarity localization for root growth [49]. PtARF7 regulated the expression of PtPIN1a/b by binding to the AuxRE motif in their promoter, thereby impacting the feedback mechanism that controls the directional movement of auxin in the cambium of poplar [50]. AtMYB88 regulated the spatiotemporal expression of AtPIN3 and AtPIN7 in root gravitropism [51]. In this study, auxin cis-regulatory elements were identified from PINs (Figure 5). MYBs, ARFs, and WRKYs might regulate the differential roles of CsPINs in fruit curving. The lower expression level of OsAUX5 was attributed to the weak activation ability of OsWRKY78 in rice [52]. Decreased expression level of CsLAX2 on the concave side of fruits may be regulated by WRKYs (Figure 7 and Figure 8). Possible interactions between ABCBs and AtABCB15–18 collaboratively contributed to the development of root structure. ZmWRKY64 positively controlled the roles of ZmABCB10/13/32 under cadmium stress [53]. CsABCB16 expression patterns were consistent with those of CsABCB18, suggesting that they have a synergistic impact and are regulated by WRKYs. Further studies are essential for exploring the relationship between WRKYs and ABCBs in cucumber.
4.3. A Model of Fruit Curving in Cucumber
In the apical hook formation in A. thaliana, the polar distribution of PIN3, PIN4, and PIN7 leads to the transfer of auxin from the central pillar to the epidermal cells outside. PINs carry auxin from the outside to the inner side of the hook, whereas AUX1 on the inner side of the hook facilitate the transfer of auxin into the cell [8,54]. The co-expression network and qPCR analyses (Figure 6) revealed that CsLAX2 and CsPIN1 worked together to promote auxin accumulation on the convex side of the curved fruit (Figure 6). The differential expression patterns of CsLAX2 and CsPIN1 contributed to the uneven distribution of auxin on both sides of the fruits (Figure 1 and Figure 6). A hypothetical model indicated that CsLAX2 was responsible for transporting extracellular auxin to the convex side, whereas CsPIN1 inhibited the auxin efflux on the convex side. Consequently, unequal distribution of auxin leads to fruit curving.
5. Conclusions
In summary, 36 auxin transporters were identified in cucumber, and the differences in their structure and function were investigated. Exogenous auxin treatment and qPCR results revealed that the polar translocation of auxin led to unequal distribution of AUX1/LAX, PIN, and ABCB proteins. Fruit curvature is attributed to the uneven distribution of auxin controlled by differential expression of CsLAX2 and CsPIN1. This study enhanced the understanding of the basic characteristics of auxin transporters and their roles in cucumber fruit curving.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture14050657/s1, Table S1: information of LAXs, PINs, and ABCBs from cucumber and Arabidopsis thaliana; Table S2: list of specific RT-qPCR primers in this study; Table S3: information on CsLAX, CsPIN, and CsABCB genes and properties of the deduced proteins in cucumber (Cucumis sativus L.); Table S4: predicted functional domains of CsABCBs; Table S5: co-expression relationship of cucumber LAXs, PINs, and ABCBs.
Author Contributions
Conceptualization, K.L. and S.L.; validation, K.L. and L.Z.; writing—original draft preparation, K.L. and L.Z.; writing—review and editing, K.L., L.F., S.L. and X.Z.; visualization, K.L. and S.L.; project administration, K.L. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China grant number 32202475, China Postdoctoral Science Foundation grant number 2021M701127, Natural Science Foundation of Heilongjiang Province grant number YQ2021C032, basic scientific research projects of provincial colleges and universities in Heilongjiang Province grant number 2022-KYYWF-1039.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All the data supporting the findings of this study are included in this manuscript and Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hohm, T.; Demarsy, E.; Quan, C.; Allenbach Petrolati, L.; Preuten, T.; Vernoux, T.; Bergmann, S.; Fankhauser, C. Plasma membrane H+-ATP ase regulation is required for auxin gradient formation preceding phototropic growth. Mol. Syst. Biol. 2014, 10, 751. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Zhang, Y. Adaptive growth: Shaping auxin-mediated root system architecture. Trends Plant Sci. 2020, 25, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, D.; Qiu, Y.; Xiao, Z.; Ji, Y.; Li, W.; Xia, Y.; Wang, Y.; Guo, H. Growth asymmetry precedes differential auxin response during apical hook initiation in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Dindas, J.; Scherzer, S.; Roelfsema, M.R.G.; von Meyer, K.; Müller, H.M.; Al-Rasheid, K.A.S.; Palme, K.; Dietrich, P.; Becker, D.; Bennett, M.J.; et al. AUX1-mediated root hair auxin influx governs SCFTIR1/AFB-type Ca2+ signaling. Nat. Commun. 2018, 9, 1174. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wang, D.; Zhang, C.; Ye, M.; Kong, N.; Ma, H.; Chen, Q. Comprehensive Analysis and Expression Profiling of PIN, AUX/LAX, and ABCB Auxin Transporter Gene Families in Solanum tuberosum under Phytohormone Stimuli and Abiotic Stresses. Biology 2021, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- An, F.; Zhang, X.; Zhu, Z.; Ji, Y.; He, W.; Jiang, Z.; Li, M.; Guo, H. Coordinated regulation of apical hook development by gibberellins and ethylene in etiolated Arabidopsis seedlings. Cell Res. 2012, 22, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Maokai, Y.; Cheng, H.; Guo, M.; Liu, Y.; Wang, L.; Chao, S.; Zhang, M.; Lai, L.; Qin, Y. Characterization of auxin transporter AUX, PIN and PILS gene families in pineapple and evaluation of expression profiles during reproductive development and under abiotic stresses. PeerJ 2021, 9, e11410. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Petrášek, J.; Žádníková, P.; Hoyerová, K.; Pešek, B.; Raz, V.; Swarup, R.; Bennett, M.; Zažímalová, E.; Benková, E. The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings. Development 2010, 137, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Tidy, A.; Abu Bakar, N.; Carrier, D.; Kerr, I.D.; Hodgman, C.; Bennett, M.J.; Swarup, R. Mechanistic insight into the role of AUXIN RESISTANCE4 in trafficking of AUXIN1 and LIKE AUX1-2. Plant Physiol. 2024, 194, 422–433. [Google Scholar] [CrossRef]
- Liu, H.; Luo, Q.; Tan, C.; Song, J.; Zhang, T.; Men, S. Biosynthesis- and transport-mediated dynamic auxin distribution during seed development controls seed size in Arabidopsis. Plant J. 2023, 113, 1259–1277. [Google Scholar] [CrossRef]
- Zhang, Y.; Berman, A.; Shani, E. Plant Hormone Transport and Localization: Signaling Molecules on the Move. Annu. Rev. Plant Biol. 2023, 74, 453–479. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.; Brockington, S.F.; Rothfels, C.; Graham, S.W.; Stevenson, D.; Kutchan, T.; Rolf, M.; Thomas, P.; Wong, G.K.-S.; Leyser, O.; et al. Paralogous Radiations of PIN Proteins with Multiple Origins of Noncanonical PIN Structure. Mol. Biol. Evol. 2014, 31, 2042–2060. [Google Scholar] [CrossRef] [PubMed]
- Hammes, U.Z.; Murphy, A.S.; Schwechheimer, C. Auxin transporters—A biochemical view. Cold Spring Harbor Perspect. Biol. 2022, 14, a039875. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Rodriguez, L.; Li, L.; Zhang, X.; Friml, J. Functional innovations of PIN auxin transporters mark crucial evolutionary transitions during rise of flowering plants. Sci. Adv. 2020, 6, eabc8895. [Google Scholar] [CrossRef] [PubMed]
- Žádníková, P.; Petrášek, J.; Marhavý, P.; Raz, V.; Vandenbussche, F.; Ding, Z.; Schwarzerová, K.; Morita, M.T.; Tasaka, M.; Hejátko, J. Role of PIN-mediated auxin efflux in apical hook development of Arabidopsis thaliana. Development 2010, 137, 607–617. [Google Scholar] [CrossRef]
- Wang, H.; Ouyang, Q.; Yang, C.; Zhang, Z.; Hou, D.; Liu, H.; Xu, H. Mutation of OsPIN1b by CRISPR/Cas9 Reveals a Role for Auxin Transport in Modulating Rice Architecture and Root Gravitropism. Int. J. Mol. Sci. 2022, 23, 8965. [Google Scholar] [CrossRef] [PubMed]
- Ung, K.L.; Winkler, M.; Schulz, L.; Kolb, M.; Janacek, D.P.; Dedic, E.; Stokes, D.L.; Hammes, U.Z.; Pedersen, B.P. Structures and mechanism of the plant PIN-FORMED auxin transporter. Nature 2022, 609, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xia, J.; Hong, J.; Zhang, C.; Wei, H.; Ying, W.; Sun, C.; Sun, L.; Mao, Y.; Gao, Y.; et al. Structural insights into auxin recognition and efflux by Arabidopsis PIN1. Nature 2022, 609, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Geisler, M.; Aryal, B.; Di Donato, M.; Hao, P. A critical view on ABC transporters and their interacting partners in auxin transport. Plant Cell Physiol. 2017, 58, 1601–1614. [Google Scholar] [CrossRef]
- Que, F.; Zhu, Y.; Liu, Q.; Wei, Q.; Ramakrishnan, M. Genome-Wide Identification, Expansion, Evolution, and Expression Analysis Reveals ABCB Genes Important for Secondary Cell Wall Development in Moso Bamboo (Phyllostachys edulis). Agronomy 2023, 13, 1828. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, Q.; Wang, X.; Mei, J.; Sharma, A.; Tripathi, D.K.; Yuan, H.; Zheng, B. Genome-wide identification and expression profiles of ABCB gene family in Chinese hickory (Carya cathayensis Sarg.) during grafting. Plant Physiol. Biochem. 2021, 168, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Kubeš, M.; Yang, H.; Richter, G.L.; Cheng, Y.; Młodzińska, E.; Wang, X.; Blakeslee, J.J.; Carraro, N.; Petrášek, J.; Zažímalová, E. The Arabidopsis concentration-dependent influx/efflux transporter ABCB4 regulates cellular auxin levels in the root epidermis. Plant J. 2012, 69, 640–654. [Google Scholar] [CrossRef]
- Cho, M.; Lee, S.H.; Cho, H.-T. P-Glycoprotein4 displays auxin efflux transporter–like action in Arabidopsis root hair cells and tobacco cells. Plant Cell 2007, 19, 3930–3943. [Google Scholar] [CrossRef]
- Wu, G.; Cameron, J.N.; Ljung, K.; Spalding, E.P. A role for ABCB19-mediated polar auxin transport in seedling photomorphogenesis mediated by cryptochrome 1 and phytochrome B. Plant J. Cell Mol. Biol. 2010, 62, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Mazzella, M.A.; Casal, J.J.; Muschietti, J.P.; Fox, A.R. Hormonal networks involved in apical hook development in darkness and their response to light. Front. Plant Sci. 2014, 5, 52. [Google Scholar] [CrossRef]
- Jenness, M.K.; Tayengwa, R.; Bate, G.A.; Tapken, W.; Zhang, Y.; Pang, C.; Murphy, A.S. Loss of multiple ABCB auxin transporters recapitulates the major twisted dwarf 1 phenotypes in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 840260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Nasser, V.; Pisanty, O.; Omary, M.; Wulff, N.; Di Donato, M.; Tal, I.; Hauser, F.; Hao, P.; Roth, O. A transportome-scale amiRNA-based screen identifies redundant roles of Arabidopsis ABCB6 and ABCB20 in auxin transport. Nat. Commun. 2018, 9, 4204. [Google Scholar] [CrossRef]
- Chen, J.; Hu, Y.; Hao, P.; Tsering, T.; Xia, J.; Zhang, Y.; Roth, O.; Njo, M.F.; Sterck, L.; Hu, Y. ABCB-mediated shootward auxin transport feeds into the root clock. EMBO Rep. 2023, 24, e56271. [Google Scholar] [CrossRef]
- Li, S.; Wang, C.; Zhou, X.; Liu, D.; Liu, C.; Luan, J.; Qin, Z.; Xin, M. The curvature of cucumber fruits is associated with spatial variation in auxin accumulation and expression of a YUCCA biosynthesis gene. Hortic. Res. 2020, 7, 35. [Google Scholar] [CrossRef]
- Jahn, L.; Hofmann, U.; Ludwig-Müller, J. Indole-3-Acetic Acid Is Synthesized by the Endophyte Cyanodermella asteris via a Tryptophan-Dependent and -Independent Way and Mediates the Interaction with a Non-Host Plant. Int. J. Mol. Sci. 2021, 22, 2651. [Google Scholar] [CrossRef]
- Weiler, E.; Jourdan, P.; Conrad, W. Levels of indole-3-acetic acid in intact and decapitated coleoptiles as determined by a specific and highly sensitive solid-phase enzyme immunoassay. Planta 1981, 153, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.-H.; Hu, S. Evolview v2: An online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. BioRxiv 2022. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xin, M.; Zhou, X.; Liu, C.; Li, S.; Liu, D.; Xu, Y.; Qin, Z. The novel ethylene-responsive factor CsERF025 affects the development of fruit bending in cucumber. Plant Mol. Biol. 2017, 95, 519–531. [Google Scholar] [CrossRef]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.-L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef]
- Li, Y.; Yang, S.; Shi, M.; Zhang, S.; Wu, S.; Chen, Y.; Li, W.; Tian, W.-M. HbARF2 and HbARF16. 3 function as negative regulators for the radial trunk growth of rubber tree. Ind. Crops Prod. 2020, 158, 112978. [Google Scholar] [CrossRef]
- Gou, H.; Nai, G.; Lu, S.; Ma, W.; Chen, B.; Mao, J. Genome-wide identification and expression analysis of PIN gene family under phytohormone and abiotic stresses in Vitis vinifera L. Physiol. Mol. Biol. Plants 2022, 28, 1905–1919. [Google Scholar] [CrossRef]
- Villaécija-Aguilar, J.A.; Körösy, C.; Maisch, L.; Hamon-Josse, M.; Petrich, A.; Magosch, S.; Chapman, P.; Bennett, T.; Gutjahr, C. KAI2 promotes Arabidopsis root hair elongation at low external phosphate by controlling local accumulation of AUX1 and PIN2. Curr. Biol. 2022, 32, 228–236. e223. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Giehl, R.F.H.; Hartmann, A.; Estevez, J.M.; Bennett, M.J.; von Wirén, N. A spatially concerted epidermal auxin signaling framework steers the root hair foraging response under low nitrogen. Curr. Biol. 2023, 33, 3926–3941.e3925. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Jiang, L.; Che, G.; Pan, Y.; Li, Y.; Hou, Y.; Zhao, W.; Zhong, Y.; Ding, L.; Yan, S. A functional allele of CsFUL1 regulates fruit length through repressing CsSUP and inhibiting auxin transport in cucumber. Plant Cell 2019, 31, 1289–1307. [Google Scholar] [CrossRef]
- Dong, X.; Ma, C.; Xu, T.; Reid, M.S.; Jiang, C.-Z.; Li, T. Auxin response and transport during induction of pedicel abscission in tomato. Hortic. Res. 2021, 8, 192. [Google Scholar] [CrossRef]
- Teale, W.D.; Pasternak, T.; Dal Bosco, C.; Dovzhenko, A.; Kratzat, K.; Bildl, W.; Schwörer, M.; Falk, T.; Ruperti, B.; V Schaefer, J.; et al. Flavonol-mediated stabilization of PIN efflux complexes regulates polar auxin transport. EMBO J. 2021, 40, e104416. [Google Scholar] [CrossRef]
- Mellor, N.L.; Voß, U.; Ware, A.; Janes, G.; Barrack, D.; Bishopp, A.; Bennett, M.J.; Geisler, M.; Wells, D.M.; Band, L.R. Systems approaches reveal that ABCB and PIN proteins mediate co-dependent auxin efflux. Plant Cell 2022, 34, 2309–2327. [Google Scholar] [CrossRef]
- Hajný, J.; Tan, S.; Friml, J. Auxin canalization: From speculative models toward molecular players. Curr. Opin. Plant Biol. 2022, 65, 102174. [Google Scholar] [CrossRef]
- Hu, J.; Su, H.; Cao, H.; Wei, H.; Fu, X.; Jiang, X.; Song, Q.; He, X.; Xu, C.; Luo, K. AUXIN RESPONSE FACTOR7 integrates gibberellin and auxin signaling via interactions between DELLA and AUX/IAA proteins to regulate cambial activity in poplar. Plant Cell 2022, 34, 2688–2707. [Google Scholar] [CrossRef]
- Wang, H.-Z.; Yang, K.-Z.; Zou, J.-J.; Zhu, L.-L.; Xie, Z.D.; Morita, M.T.; Tasaka, M.; Friml, J.; Grotewold, E.; Beeckman, T. Transcriptional regulation of PIN genes by FOUR LIPS and MYB88 during Arabidopsis root gravitropism. Nat. Commun. 2015, 6, 8822. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, Y.; Sun, Y.; Xie, Z.; Luo, Y.; Long, Q.; Feng, J.; Liu, X.; Wang, B.; He, D. Natural variations of OsAUX5, a target gene of OsWRKY78, control the neutral essential amino acid content in rice grains. Mol. Plant 2023, 16, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Hou, Y.; Sun, Y.; Chen, X.; Wang, G.; Wang, H.; Zhu, B.; Du, X. The maize WRKY transcription factor ZmWRKY64 confers cadmium tolerance in Arabidopsis and maize (Zea mays L.). Plant Cell Rep. 2024, 43, 44. [Google Scholar] [CrossRef]
- Béziat, C.; Barbez, E.; Feraru, M.I.; Lucyshyn, D.; Kleine-Vehn, J. Light triggers PILS-dependent reduction in nuclear auxin signalling for growth transition. Nat. Plants 2017, 3, 17105. [Google Scholar] [CrossRef]
Figure 1.
Effects of exogenous auxin treatment on cucumber fruits. (a) Exogenous auxin application on the straight and curved fruits at 0 DPA. Auxin content in the (b) control, (c) IAA treatment, (d) NPA treatment, and (e) NOA treatment at 4 DPA. S1 and S2 represent both sides of the straight fruits; C1 and C2 represent concave and convex sides of the curved fruits, respectively. Scale bar = 3 cm. Data represent the average of three replicates with standard errors. Different letters indicate significant differences (p < 0.05).
Figure 1.
Effects of exogenous auxin treatment on cucumber fruits. (a) Exogenous auxin application on the straight and curved fruits at 0 DPA. Auxin content in the (b) control, (c) IAA treatment, (d) NPA treatment, and (e) NOA treatment at 4 DPA. S1 and S2 represent both sides of the straight fruits; C1 and C2 represent concave and convex sides of the curved fruits, respectively. Scale bar = 3 cm. Data represent the average of three replicates with standard errors. Different letters indicate significant differences (p < 0.05).
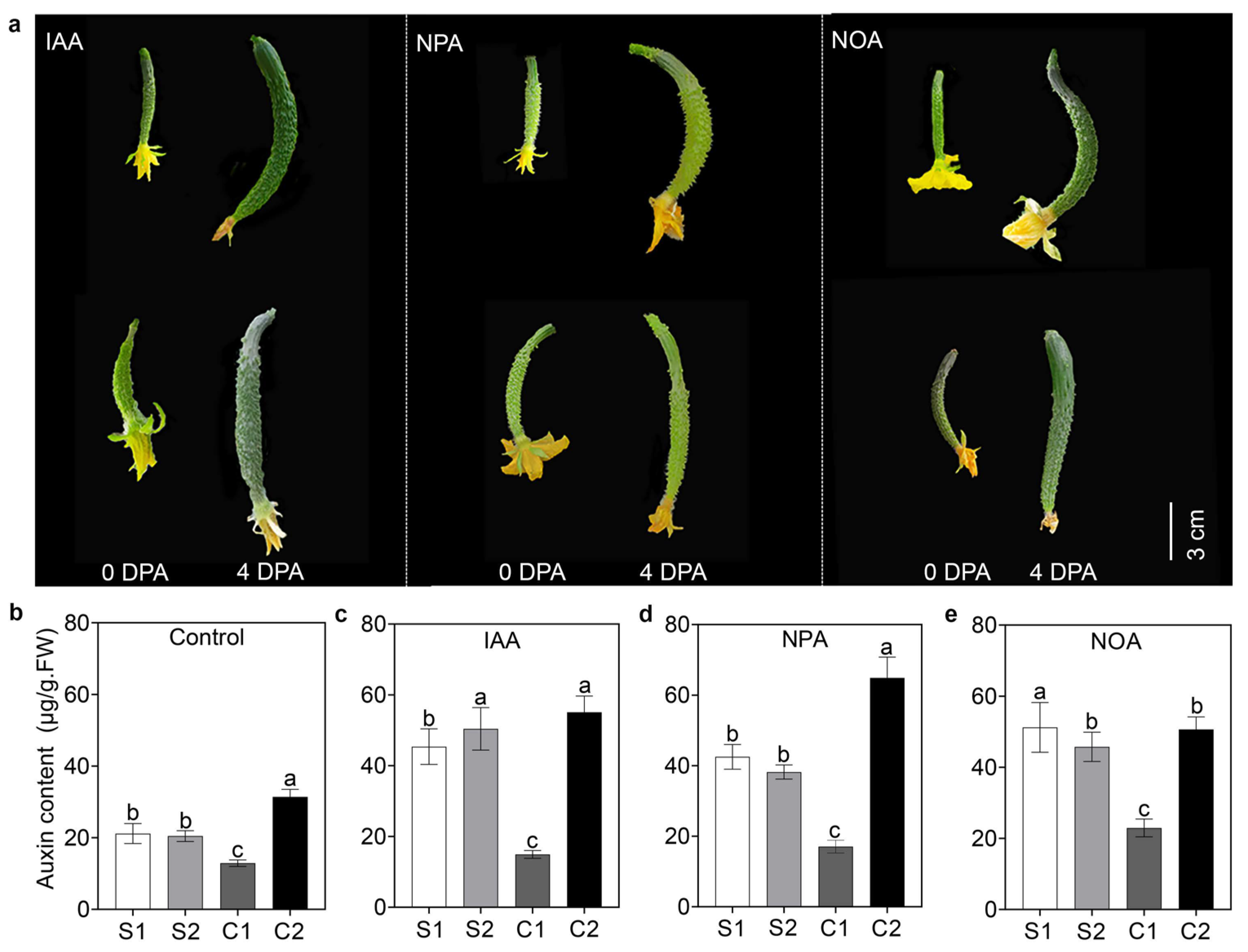
Figure 2.
Chromosome localization and distribution of duplication events of CsLAXs, CsPINs, and CsABCBs.
Figure 2.
Chromosome localization and distribution of duplication events of CsLAXs, CsPINs, and CsABCBs.

Figure 3.
Phylogenetic relationship and synteny analysis of auxin transporters in cucumber and Arabidopsis. Phylogenetic trees of (a) AUX1/LAXs, (b) PINs, and (c) ABCBs. Different subfamilies are marked using different colors. The family members from cucumber and Arabidopsis are marked with red and black squares, respectively. (d) Synteny analysis. Colored lines connecting two chromosomal regions indicate syntenic regions between cucumber (Cs1–7) and Arabidopsis (At1–5) chromosomes.
Figure 3.
Phylogenetic relationship and synteny analysis of auxin transporters in cucumber and Arabidopsis. Phylogenetic trees of (a) AUX1/LAXs, (b) PINs, and (c) ABCBs. Different subfamilies are marked using different colors. The family members from cucumber and Arabidopsis are marked with red and black squares, respectively. (d) Synteny analysis. Colored lines connecting two chromosomal regions indicate syntenic regions between cucumber (Cs1–7) and Arabidopsis (At1–5) chromosomes.

Figure 4.
The motif and gene structure of CsLAXs (a), CsPINs (b), and CsABCBs (c).

Figure 5.
The prediction of cis-regulatory elements in the promoters of auxin transporters.
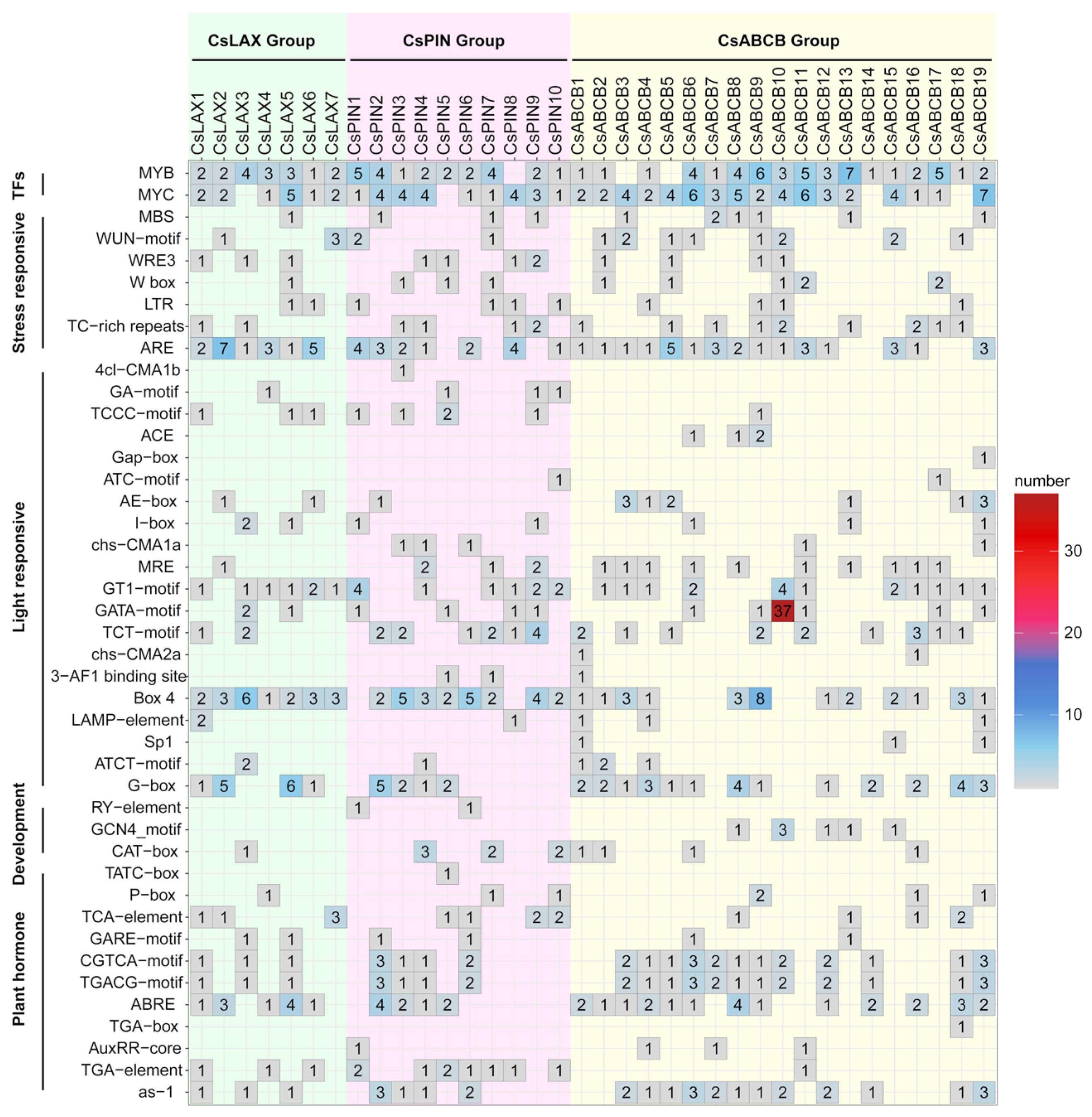
Figure 6.
Expression patterns and clustering analysis of auxin transporters under (a) IAA treatment and (b) NPA treatment. S1 and S2 represent both sides of the straight fruits; C1 and C2 represent the concave and convex sides of curved fruits, respectively.
Figure 6.
Expression patterns and clustering analysis of auxin transporters under (a) IAA treatment and (b) NPA treatment. S1 and S2 represent both sides of the straight fruits; C1 and C2 represent the concave and convex sides of curved fruits, respectively.

Figure 7.
Co-expression network and expression profiles. (a) The co-expression network of LAXs, PINs, and ABCBs on the concave and convex sides of curved cucumber fruits at 2 DPA was established when the Pearson correlation was higher than 0.97 or lower than −0.97. Different colored circles represent different auxin transport gene families. Red and blue lines show that the Pearson correlation was higher than 0.99 and lower than −0.99, respectively. (b) The mRNA levels on the concave and convex sides of curved fruits at 2, 4, 6, 8, and 10 DPA. CsEF1a was used as the reference gene. The data are the means (±SEs) of three independent experiments, with three replicates each (* p < 0.05, and ** p < 0.01).
Figure 7.
Co-expression network and expression profiles. (a) The co-expression network of LAXs, PINs, and ABCBs on the concave and convex sides of curved cucumber fruits at 2 DPA was established when the Pearson correlation was higher than 0.97 or lower than −0.97. Different colored circles represent different auxin transport gene families. Red and blue lines show that the Pearson correlation was higher than 0.99 and lower than −0.99, respectively. (b) The mRNA levels on the concave and convex sides of curved fruits at 2, 4, 6, 8, and 10 DPA. CsEF1a was used as the reference gene. The data are the means (±SEs) of three independent experiments, with three replicates each (* p < 0.05, and ** p < 0.01).
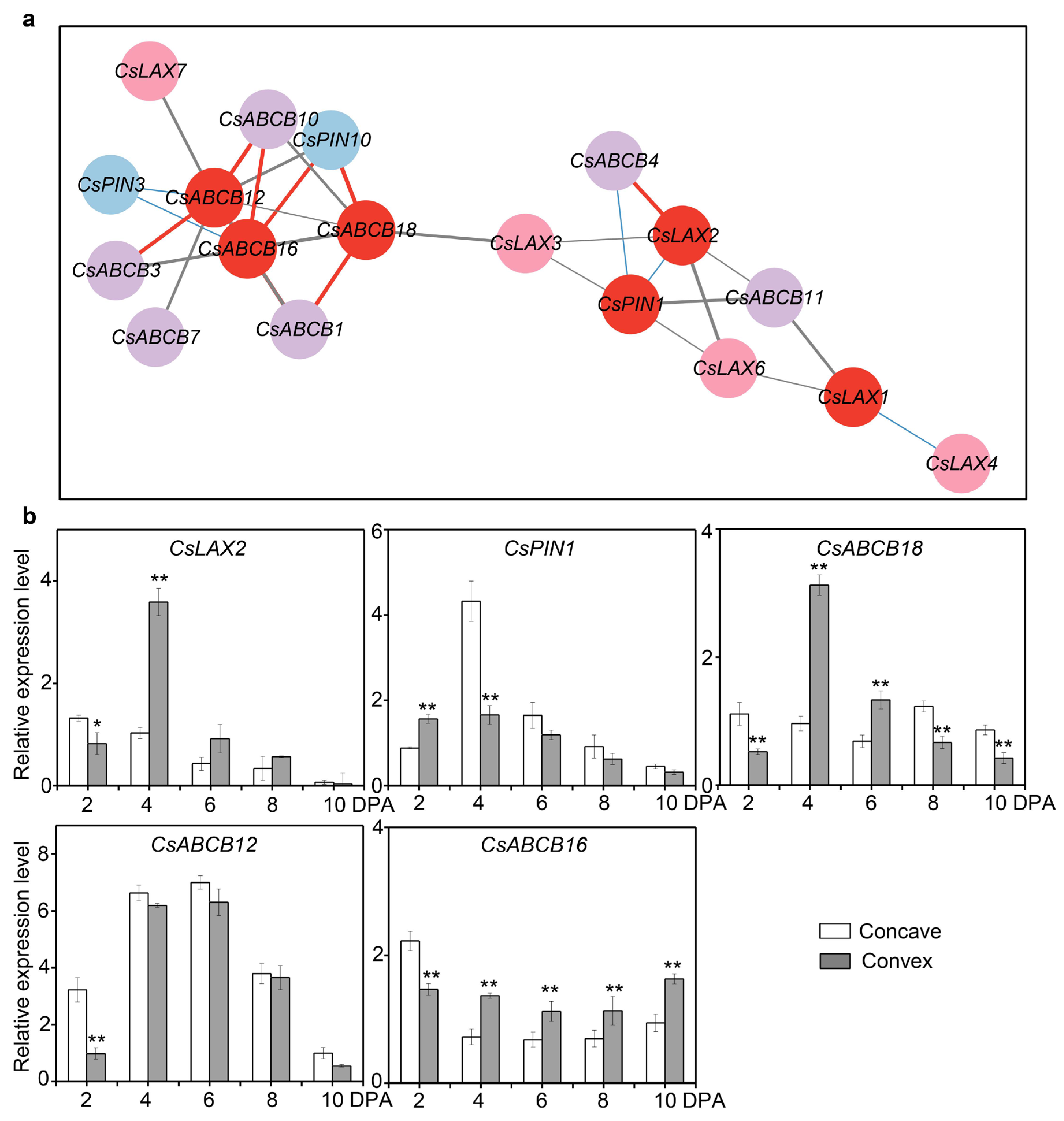
Figure 8.
Co-expression network of TFs and guide genes in CsLAX (a), CsPIN (b) and CsABCB (c) group at 2 DPA. The co-expression relationship was established when the Pearson correlation was higher than 0.97. Different colored circles represent different TFs. Red lines indicate that the Pearson correlation was higher than 0.99.
Figure 8.
Co-expression network of TFs and guide genes in CsLAX (a), CsPIN (b) and CsABCB (c) group at 2 DPA. The co-expression relationship was established when the Pearson correlation was higher than 0.97. Different colored circles represent different TFs. Red lines indicate that the Pearson correlation was higher than 0.99.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lu, K.; Zhang, L.; Fan, L.; Zhou, X.; Li, S. A Genome-Wide Comparative Analysis of AUX1/LAX, PIN, and ABCB Genes Reveals Their Roles in Cucumber Fruit Curving. Agriculture 2024, 14, 657. https://doi.org/10.3390/agriculture14050657
AMA Style
Lu K, Zhang L, Fan L, Zhou X, Li S. A Genome-Wide Comparative Analysis of AUX1/LAX, PIN, and ABCB Genes Reveals Their Roles in Cucumber Fruit Curving. Agriculture. 2024; 14(5):657. https://doi.org/10.3390/agriculture14050657
Chicago/Turabian StyleLu, Ke, La Zhang, Lianxue Fan, Xiuyan Zhou, and Shengnan Li. 2024. "A Genome-Wide Comparative Analysis of AUX1/LAX, PIN, and ABCB Genes Reveals Their Roles in Cucumber Fruit Curving" Agriculture 14, no. 5: 657. https://doi.org/10.3390/agriculture14050657
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.