
Heterogeneity of Intramuscular, Intermuscular, and Subcutaneous Fat in Laiwu Pigs: Insights from Targeted Lipidomics and Transcriptomics
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Samples Preparation
2.2. Hematoxylin–Eosin (H&E) Staining
2.3. Immunofluorescence Staining
2.4. Lipidomics Analyses
2.5. Unsupervised Multivariate Data Analyses
2.6. Quantitative Real-Time PCR(qPCR)
2.7. RNA Sequencing (RNA-Seq) Analysis and Differentially Expressed Gene Screening
2.8. Bioinformatic Analysis
2.9. Statistical Analysis
3. Results
3.1. The Size of IMF Adipocytes is Smaller than BF and IMAT
- To explore the lipid and molecular characteristics of the IMF of Laiwu pigs, we collected the middle layer of BF, IMAT of the hind leg, and IMF-rich LDM from 300-day-old Laiwu pigs. We performed hematoxylin-eosin (H&E) staining on the BF, IMAT, and LDM samples to assess the adipocyte morphology and found the size of IMF adipocytes within the LDM was smaller than the BF and IMAT adipocytes (Figure 1A). Quantitative analysis revealed that BF adipocytes are primarily distributed over areas ranging from 5000 to 15,000 μm², with an average area of 11,265 μm². IMAT adipocytes mainly span areas between 4000 and 11,000 μm², with an average area of 6205 μm². Meanwhile, IMF adipocytes predominantly occupy areas from 2000 to 5000 μm², with an average area of 3356 μm² (Figure 1B,C). These findings imply a comparatively slower development of IMF relative to BF and IMAT.
3.2. Alterations in Lipid Class Composition across IMF, IMAT, and BF
- To explore the differences in the overall lipid composition and distribution in different adipose tissues, we carried out lipidomic analysis through liquid chromatography-tandem mass spectrometry (LC-MS/MS). This analysis identified over 467 lipid species distributed among 29 lipid classes in the BF, IMAT, and IMF (Figure 2A). These lipid classes included triacylglycerols (TAGs), alkyl-phosphatidylethanolamines (PE-Os), phosphatidylcholines (PCs), phosphatidylethanolamines (PEs), galactosylceramides (GalCers), phosphatidyl-inositols (PIs), and others. The abbreviations of these quantified lipid classes are shown in Figure S1A. Multivariate analysis of the data from the 467 lipid molecules across the three tissue types revealed a close relationship between the BF and IMAT groups, whereas the IMF group exhibited a distinct profile (Figure S1B–D). Utilizing the PLS-DA model, 70 critical variables were identified that differentiated the IMF group from the BF and IMAT groups, based on VIP values >1.0 and p-values < 0.05. Predominantly, these variables included CLs, PE-Os, PC-Os, AcCas, and GalCers, which were found to be elevated in IMF and demonstrated high positive correlations amongst themselves (Figure 2B and Figure S2B). A total of 152 and 93 lipid species in the IMF group were identified as significantly changed compared to the BF and IMAT groups based on p-values < 0.05, respectively. The downregulated lipid species within the IMF group included FFAs, DAGs, and TAGs (Figure 2C and Figure S2B). Furthermore, the IMF exhibited a notable reduction in glycerolipid levels alongside a significant elevation in the concentrations of fatty acyls, sphingolipids, glycerophospholipids, and sulfatides (Figure 2D). Of these, the FFA and DAG contents were significantly lower, and AcCa, CL, PS, PC-O, PE-O, S1P, SM, GM3, and GalCer contents were significantly higher in the IMF (Figure 2E), while other lipid subclasses were unchanged among the three adipose tissues (Figure S2C–F). These findings further underscored substantial alterations in the composition and content of lipids in IMF compared to BF and IMAT.
3.3. IMF Exhibited a Reduction in TAG Species Characterized by Longer Acyl Chain Carbon Numbers and More Double Bonds
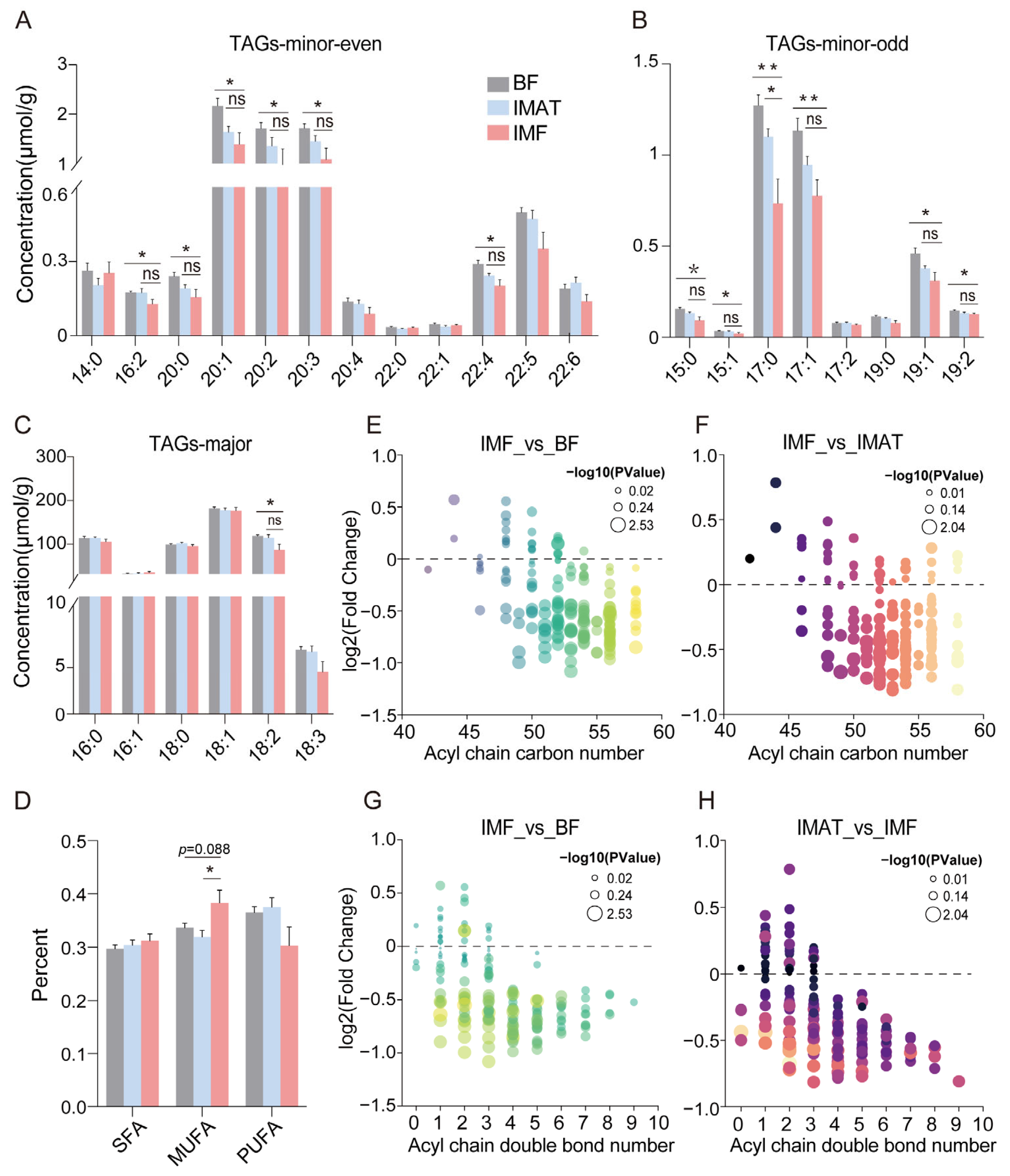
- TAGs and glycerophospholipids are the two most abundant lipid types in adipose tissues, with their associated individual fatty acyl chains mirroring the composition of the predominant fatty acids. The results showed the content of most fatty acyl chains associated with TAG was significantly decreased in the IMF group compared to the BF group, with only the C17:0 fatty acyl chain showing a significant decrease compared to the IMAT group (Figure 3A,B). Notably, the content of the LA (C18:2) fatty acyl chain associated with TAGs was decreased in the IMF group relative to BF (p < 0.05) and IMAT groups (p = 0.09) (Figure 3C); LA is an essential fatty acid with documented health benefits [19]. Moreover, the percentage of MUFA chains associated with TAGs was decreased in the IMF relative to BF (p < 0.05) and IMAT (p = 0.088), while the SFA and PUFA chains showed no significant differences among the three adipose tissue groups (Figure 3D). Furthermore, the IMF exhibited a marked decrease in TAG content as acyl chain carbon numbers increased (Figure 3E,F) or as the number of double bonds rose (Figure 3G,H). These observations suggest that the fatty acid profile as reflected by TAGs undergoes significant modifications in IMF in contrast to BF and IMAT.
3.4. IMF Exhibited Significantly Higher Levels of Glycerophospholipid Species Characterized by Longer Acyl Chain Carbon Numbers and Increased Double Bonds
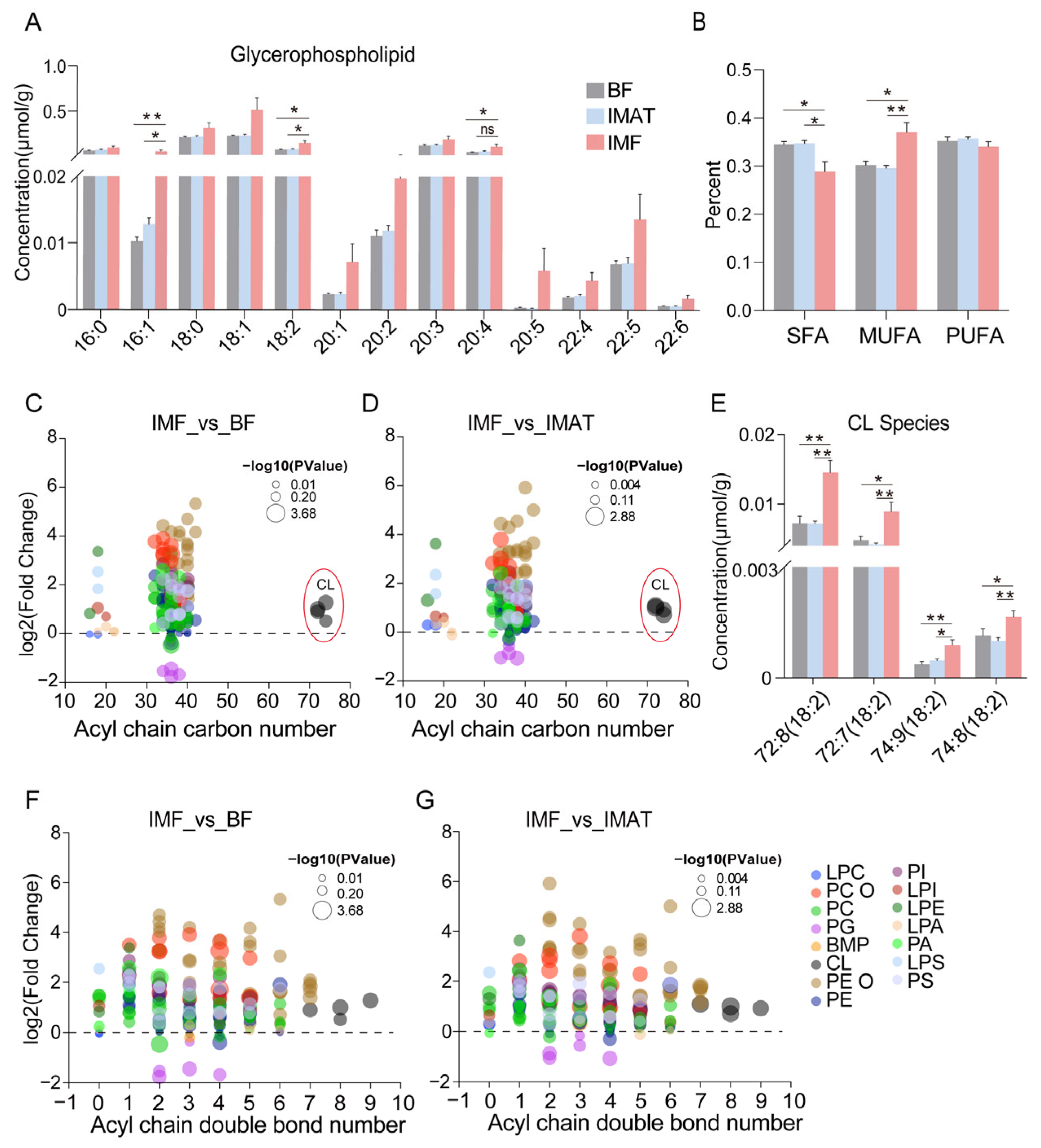
- We next analyzed the individual fatty acyl chain composition associated with glycerophospholipids. The concentration of C16:1 and C18:2 fatty acyl chains in the glycerophospholipids pool was notably increased in IMF compared to BF and IMAT (Figure 4A). We analyzed the total contents of SFA, MUFA, and PUFA chains associated with glycerophospholipids. The SFA percentage was significantly lower, and the MUFA percentage was significantly higher in IMF; the PUFA percentage was unaffected (Figure 4B). Moreover, the IMF displayed elevated levels of glycerophospholipid species with lower (below 30 carbons, mostly associated with LPCs, LPEs, and LPSs), intermediate (above 30 and below 50 carbons, particularly associated with PC-O and PE-O), and higher (above 70 carbons, associated with CLs) acyl chain carbon numbers (Figure 4C,D). Particularly, all four significantly increased CLs contained C18:2 fatty acyl chains (Figure 4E); cardiolipins (CLs) play a crucial role in maintaining mitochondrial membrane integrity and function [20]. Additionally, the IMF displayed significantly higher levels of most glycerophospholipids species with greater acyl chain double bonds compared to BF and IMAT groups (Figure 4F,G). These results suggest a dramatic change in the composition and fatty acyl chains within glycerophospholipids in IMF.
3.5. IMF Exhibited a Significant Increase in Fatty Acyl Species, Characterized by Longer Acyl Chain Carbon Numbers and More Double Bonds
- The above-described results showed a significant increase in the total content of fatty acyls in the IMF. Fatty acyls contain FFAs and AcCas, which are fatty acid metabolites that play important roles in many cellular energy metabolism pathways and are important diagnostic markers for mitochondrial-related metabolic syndrome [21]. We analyzed the composition of fatty acyl chains associated with fatty acyls and found most FFA species significantly decreased, while AcCas species markedly increased in the IMF and compared to BF and IMAT (Figure 5A,B). Particularly, in line with the alteration of C18:2 associated with TAGs and glycerophospholipids, FFA (C18:2) and AcCa (C18:2) exhibited a significant change in the IMF (Figure 5A,B). Moreover, IMF showed a greatly lower percentage of SFA chains and a notably higher percentage of MUFA chains associated with fatty acyls, with an unaffected percentage of PUFA chains relative to BF and IMAT (Figure 5C). Additionally, FFA species showed a significant decrease in IMF, but AcCa species exhibited a significant increase conversely within the IMF with varying acyl chain carbon numbers (Figure 5D,E) and acyl chain double bonds (Figure 5F,G). These results suggest the composition and acyl chains of fatty acyl species considerably change in IMF.
3.6. IMF Isolated from the LDM Constitutes a Unique Adipose Tissue, Exhibiting Distinct Molecular Characteristics Compared to BF and IMAT
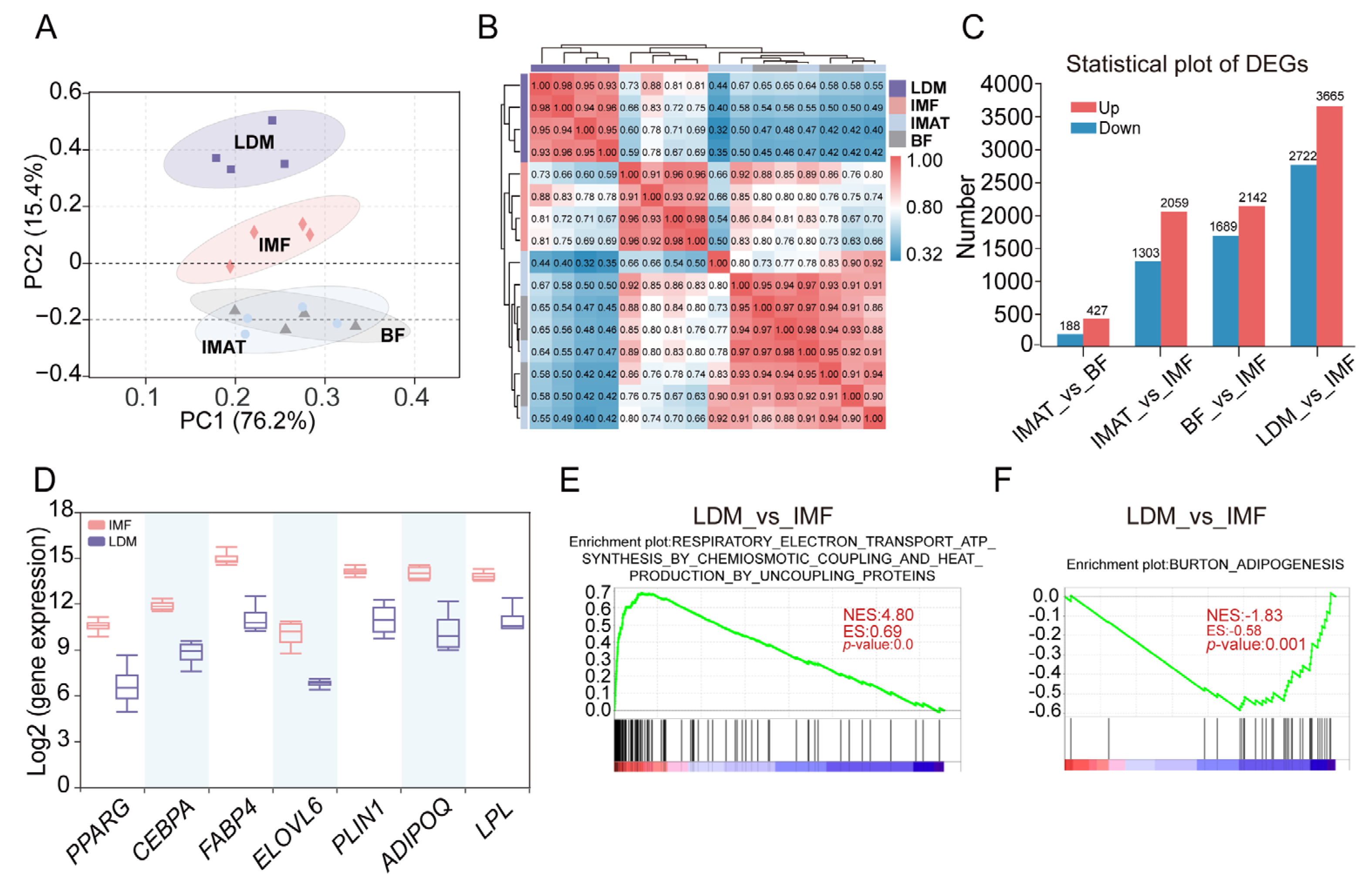
- We conducted RNA-Seq analysis on IMF, BF, and IMAT samples obtained from Laiwu pigs to investigate transcriptional changes and lipid metabolic pathways in adipose tissues. The IMF samples were manually dissected from the LDM under a stereomicroscope, and RNA extraction was successfully performed. To validate the adipose tissue specificity of the IMF samples, we included RNA-Seq data from LDM samples for comparison. PCA based on FPKM values of commonly expressed genes in all four tissues showed that BF and IMAT samples closely clustered together, while IMF samples occupied an intermediate position between BF (or IMAT) and LDM samples on the second axis of the PCA plot, indicating gene expression patterns in IMF were different from BF, IMAT, and LDM (Figure 6A). Moreover, Pearson’s correlation coefficients demonstrated significantly greater similarity between pairwise BF (or IMAT) and IMF samples compared to pairwise IMF and LDM samples (Figure 6B). This observation was supported by the identification of a greater number of differentially expressed genes between IMF and LDM than between IMF and BF (or IMAT) (Figure 6C). We identified genes associated with adipogenesis (PPARG, CEBPA), lipogenesis (FABP4, ELOVL6, PLIN1), and adipocytokines (ADIPOQ, LPL) that exhibited higher expression in IMF compared with LDM (Figure 6D). Furthermore, GSEA showed that RESPIRATORY_ELECTRON_TRANSPORT_ATP_SYNTHESIS_BY_CHEMIOSMOTIC_COPLING_AND_HEAT_PRODUCTION_BY_UNCOUPLING_PROTEINS (NES = 4.80, p = 0.0) was greatly upregulated in LDM, but ADIPOGENESIS (NES= −1.83, p = 0.0001) was greatly upregulated in IMF (Figure 6E,F). These findings reveal that IMF samples, isolated from the LDM, represent largely adipose tissue and possess distinct molecular characteristics from BF and IMAT.
3.7. Transcriptomic Gene Enrichment Analysis Indicates a Reduction in Lipogenesis and an Enhanced Innervation in IMF
- To better characterize the molecular signature of the IMF, we performed a differential gene expression analysis on IMF, BF, and IMAT. We identified 1054 BF and 992 IMF genes that exhibited a significantly higher expression than the corresponding LDM genes (fold change > 2 and p-value < 0.05) (Figure 7A). Moreover, 755 IMAT and 861 IMF genes exhibited a significantly higher expression than the corresponding LDM genes, respectively (Figure S4A). GO and KEGG pathway enrichment analysis with genes differentially expressed in BF, IMAT, and IMF. The 1054 genes highly expressed in BF and 755 genes highly expressed in IMAT were commonly involved in the PPAR signaling pathway, glycerolipid metabolism, unsaturated fatty acid biosynthesis, fatty acid elongation, and lipid storage (Figure 7B and Figure S4B), which was consistent with the significant higher glycerolipid content and unsaturated and long-chain fatty acids associated with TAGs. In comparison, the IMF genes were associated with axon guidance, the cAMP signaling pathway, cell adhesion molecules, glutamatergic synapse, dopaminergic synapse, postsynaptic density, insulin secretion, calcium signaling, and ATP synthesis coupled electron transport (Figure 7C and Figure S4C). Moreover, gene set enrichment analysis (GSEA) analysis showed the ADIPOGENESIS pathway was significantly downregulated (Figure 7D and Figure S4D), while the NEURONAL_SYSTEM pathway was notably upregulated in IMF (Figure 7E and Figure S4E). The heatmap showed significant differences in gene expression between IMF and BF (or IMAT) in the PPAR signaling pathway, glycerolipid metabolism, unsaturated fatty acid biosynthesis, axon guidance, and cAMP signaling pathway (Figure 7F,G and Figure S4F,G).
- qPCR validation showed the genes related to glycerolipid metabolism and lipogenesis significantly decreased in IMF compared to BF (Figure 7H). The genes related to axon guidance and the cAMP signaling pathway were increased in the IMF, including BMP7 (p = 0.077) and PPP1R12A (p < 0.01) (Figure 7I). In particular, the gene UGT8, which encodes the rate-limiting enzyme for GalCer biosynthesis, was significantly increased in IMF compared with BF (Figure 7I). Furthermore, immunostaining analysis found tyrosine hydroxylase (TH), the marker of sympathetic innervation, was more densely distributed in IMF adipocytes (Figure 7J). Taken together, these results suggest that IMF has a lower capacity for fat deposition and receives stronger innervation.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drewnowski, A. Perspective: The Place of Pork Meat in Sustainable Healthy Diets. Adv. Nutr. 2024, 15, 100213. [Google Scholar] [CrossRef] [PubMed]
- Warriss, P.D.; Brown, S.N.; Franklin, J.G.; Kestin, S.C. The thickness and quality of backfat in various pig breeds and their relationship to intramuscular fat and the setting of joints from the carcasses. Meat Sci. 1990, 28, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Font-i-Furnols, M.; Tous, N.; Esteve-Garcia, E.; Gispert, M. Do all the consumers accept marbling in the same way? The relationship between eating and visual acceptability of pork with different intramuscular fat content. Meat Sci. 2012, 91, 448–453. [Google Scholar] [CrossRef]
- Lonergan, S.M.; Huff-Lonergan, E.; Rowe, L.J.; Kuhlers, D.L.; Jungst, S.B. Selection for lean growth efficiency in Duroc pigs influences pork quality. J. Anim. Sci. 2001, 79, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Newcom, D.W.; Baas, T.J.; Schwab, C.R.; Stalder, K.J. Genetic and phenotypic relationships between individual subcutaneous backfat layers and percentage of longissimus intramuscular fat in Duroc swine1. J. Anim. Sci. 2005, 83, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Poklukar, K.; Čandek-Potokar, M.; Batorek Lukač, N.; Tomažin, U.; Škrlep, M. Lipid Deposition and Metabolism in Local and Modern Pig Breeds: A Review. Animals 2020, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.J.; Megens, H.J.; Martinez Barrio, A.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, Ö.; Jern, P.; Jørgensen, C.B.; et al. Strong signatures of selection in the domestic pig genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529–19536. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, E.; Wing, A.; Holtrup, B.; Sebo, Z.; Kaplan, J.L.; Saavedra-Peña, R.; Church, C.D.; Colman, L.; Berry, R.; Rodeheffer, M.S. The Adipose Tissue Microenvironment Regulates Depot-Specific Adipogenesis in Obesity. Cell Metab. 2016, 24, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Leiria, L.O.; Tseng, Y.H. Lipidomics of brown and white adipose tissue: Implications for energy metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158788. [Google Scholar] [CrossRef]
- Lam, S.M.; Tian, H.; Shui, G. Lipidomics, en route to accurate quantitation. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 752–761. [Google Scholar] [CrossRef]
- Ogawa, M.; Lester, R.; Akima, H.; Gorgey, A.S. Quantification of intermuscular and intramuscular adipose tissue using magnetic resonance imaging after neurodegenerative disorders. Neural Regen. Res. 2017, 12, 2100–2105. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhang, Y.; Wu, J.; Qiao, M.; Xu, Z.; Peng, X.; Mei, S. Proteomic and lipidomic analyses reveal saturated fatty acids, phosphatidylinositol, phosphatidylserine, and associated proteins contributing to intramuscular fat deposition. J. Proteom. 2021, 241, 104235. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Fang, G.-f.; Wang, S.-d.; Wang, H.; Zeng, Y.-q. Longissimus lumborum muscle transcriptome analysis of Laiwu and Yorkshire pigs differing in intramuscular fat content. Genes Genom. 2017, 39, 759–766. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, R.; Yang, M.; Niu, N.; Wu, J.; Shu, Z.; Zhang, P.; Shi, L.; Zhao, F.; Wang, L.; et al. Metabolomics and lipidomics profiles related to intramuscular fat content and flavor precursors between Laiwu and Yorkshire pigs. Food Chem. 2023, 404, 134699. [Google Scholar] [CrossRef]
- Lam, S.M.; Li, J.; Sun, H.; Mao, W.; Lu, Z.; Zhao, Q.; Han, C.; Gong, X.; Jiang, B.; Chua, G.H.; et al. Quantitative Lipidomics and Spatial MS-Imaging Uncovered Neurological and Systemic Lipid Metabolic Pathways Underlying Troglomorphic Adaptations in Cave-Dwelling Fish. Mol. Biol. Evol. 2022, 39, msac050. [Google Scholar] [CrossRef]
- Pan, J.; Tao, C.; Cao, C.; Zheng, Q.; Lam, S.M.; Shui, G.; Liu, X.; Li, K.; Zhao, J.; Wang, Y. Adipose lipidomics and RNA-Seq analysis revealed the enhanced mitochondrial function in UCP1 knock-in pigs. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2019, 1864, 1375–1383. [Google Scholar] [CrossRef]
- Lin, J.; Cao, C.; Tao, C.; Ye, R.; Dong, M.; Zheng, Q.; Wang, C.; Jiang, X.; Qin, G.; Yan, C.; et al. Cold adaptation in pigs depends on UCP3 in beige adipocytes. J. Mol. Cell Biol. 2017, 9, 364–375. [Google Scholar] [CrossRef]
- Hamilton, J.S.; Klett, E.L. Linoleic acid and the regulation of glucose homeostasis: A review of the evidence. Prostaglandins Leukot. Essent. Fat. Acids 2021, 175, 102366. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; De Benedictis, V.; Ruggiero, F.M.; Petrosillo, G. Functional role of cardiolipin in mitochondrial bioenergetics. Biochim. Biophys. Acta 2014, 1837, 408–417. [Google Scholar] [CrossRef]
- Dambrova, M.; Makrecka-Kuka, M.; Kuka, J.; Vilskersts, R.; Nordberg, D.; Attwood, M.M.; Smesny, S.; Sen, Z.D.; Guo, A.C.; Oler, E.; et al. Acylcarnitines: Nomenclature, Biomarkers, Therapeutic Potential, Drug Targets, and Clinical Trials. Pharmacol. Rev. 2022, 74, 506–551. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.; Chen, H.; Zhang, X.; Liu, X.; He, Z.; Cong, P.; Chen, Y.; Mo, D. Comparative Transcriptome Analysis Reveals a More Complicated Adipogenic Process in Intramuscular Stem Cells than That of Subcutaneous Vascular Stem Cells. J. Agric. Food Chem. 2019, 67, 4700–4708. [Google Scholar] [CrossRef] [PubMed]
- Gardan, D.; Gondret, F.; Louveau, I. Lipid metabolism and secretory function of porcine intramuscular adipocytes compared with subcutaneous and perirenal adipocytes. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E372–E380. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, K.; Evans, T.; Hla, T. Sphingosine 1-phosphate signalling. Development 2014, 141, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.S.B.; Færgeman, N.J. Sphingolipids: Membrane microdomains in brain development, function and neurological diseases. Open Biol. 2017, 7, 170069. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.; Dulovic-Mahlow, M.; Mandik, F.; Frese, L.; Kanana, Y.; Haissatou Diaw, S.; Depperschmidt, J.; Böhm, C.; Rohr, J.; Lohnau, T.; et al. Ceramide accumulation induces mitophagy and impairs β-oxidation in PINK1 deficiency. Proc. Natl. Acad. Sci. USA 2021, 118, e2025347118. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Suzuki, T. Role of sulfatide in normal and pathological cells and tissues. J. Lipid Res. 2012, 53, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. In Sphingolipids as Signaling and Regulatory Molecules; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2010; Volume 688, pp. 1–23. [Google Scholar] [CrossRef]
- Eckhardt, M. The role and metabolism of sulfatide in the nervous system. Mol. Neurobiol. 2008, 37, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Lam, S.M.; Shui, G. Emerging roles and therapeutic potentials of sphingolipids in pathophysiology: Emphasis on fatty acyl heterogeneity. J. Genet. Genom. 2023, 51, 268–278. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Matsuzaki, E.; Higashi, K.; Takahashi-Yanaga, F.; Takano, A.; Hirata, M.; Nishimura, F. Sphingosine-1-phosphate inhibits differentiation of C3H10T1/2 cells into adipocyte. Mol. Cell Biochem. 2015, 401, 39–47. [Google Scholar] [CrossRef]
- Moon, M.H.; Jeong, J.K.; Lee, Y.J.; Seol, J.W.; Park, S.Y. Sphingosine-1-phosphate inhibits the adipogenic differentiation of 3T3-L1 preadipocytes. Int. J. Mol. Med. 2014, 34, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Garcia-Carbonell, R.; Yamachika, S.; Zhao, P.; Dhar, D.; Loomba, R.; Kaufman, R.J.; Saltiel, A.R.; Karin, M. ER Stress Drives Lipogenesis and Steatohepatitis via Caspase-2 Activation of S1P. Cell 2018, 175, 133–145.e115. [Google Scholar] [CrossRef] [PubMed]
- Brites, P.; Waterham, H.R.; Wanders, R.J. Functions and biosynthesis of plasmalogens in health and disease. Biochim. Biophys. Acta 2004, 1636, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Indiveri, C.; Iacobazzi, V.; Tonazzi, A.; Giangregorio, N.; Infantino, V.; Convertini, P.; Console, L.; Palmieri, F. The mitochondrial carnitine/acylcarnitine carrier: Function, structure and physiopathology. Mol. Asp. Med. 2011, 32, 223–233. [Google Scholar] [CrossRef] [PubMed]
- El-Hafidi, M.; Correa, F.; Zazueta, C. Mitochondrial dysfunction in metabolic and cardiovascular diseases associated with cardiolipin remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165744. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; He, A.; Tan, M.; Johnson, J.M.; Dean, J.M.; Pietka, T.A.; Chen, Y.; Zhang, X.; Hsu, F.F.; Razani, B.; et al. Peroxisome-derived lipids regulate adipose thermogenesis by mediating cold-induced mitochondrial fission. J. Clin. Investig. 2019, 129, 694–711. [Google Scholar] [CrossRef]
- Boyman, L.; Karbowski, M.; Lederer, W.J. Regulation of Mitochondrial ATP Production: Ca(2+) Signaling and Quality Control. Trends Mol. Med. 2020, 26, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, L.; Chen, W.; Li, J.; Shan, T. CRTC3 Regulates the Lipid Metabolism and Adipogenic Differentiation of Porcine Intramuscular and Subcutaneous Adipocytes by Activating the Calcium Pathway. J. Agric. Food Chem. 2021, 69, 7243–7255. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, A.; Tejerina, S.; Raes, M.; Keijer, J.; Arnould, T. Mitochondrial (dys)function in adipocyte (de)differentiation and systemic metabolic alterations. Am. J. Pathol. 2009, 175, 927–939. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, L.; Zhang, J.; Liu, X.; Zhang, Y.; Cai, L.; Zhang, W.; Cui, L.; Yang, J.; Ji, J.; et al. A large-scale comparison of meat quality and intramuscular fatty acid composition among three Chinese indigenous pig breeds. Meat Sci. 2020, 168, 108182. [Google Scholar] [CrossRef]
- Kim, J.A.; Cho, E.S.; Jeong, Y.D.; Choi, Y.H.; Kim, Y.S.; Choi, J.W.; Kim, J.S.; Jang, A.; Hong, J.K.; Sa, S.J. The effects of breed and gender on meat quality of Duroc, Pietrain, and their crossbred. J. Anim. Sci. Technol. 2020, 62, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Glazier, B.J.; Hinkel, B.C.; Cao, J.; Liu, L.; Liang, C.; Shi, H. Neuroendocrine Regulation of Energy Metabolism Involving Different Types of Adipose Tissues. Int. J. Mol. Sci. 2019, 20, 2707. [Google Scholar] [CrossRef] [PubMed]
- Ravnskjaer, K.; Madiraju, A.; Montminy, M. Role of the cAMP pathway in glucose and lipid metabolism. In Metabolic Control; Springer: Cham, Switzerland, 2016; pp. 29–49. [Google Scholar]
- Zechner, R.; Madeo, F.; Kratky, D. Cytosolic lipolysis and lipophagy: Two sides of the same coin. Nat. Rev. Mol. Cell Biol. 2017, 18, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Y.; Wu, Z.; Xiong, X.; Zhang, J.; Ma, J.; Xiao, S.; Huang, L.; Yang, B. Subcutaneous and intramuscular fat transcriptomes show large differences in network organization and associations with adipose traits in pigs. Sci. China Life Sci. 2021, 64, 1732–1746. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Wu, T.; Lam, S.M.; Shui, G.; Yang, S.; Wang, Y.; Tao, C. Heterogeneity of Intramuscular, Intermuscular, and Subcutaneous Fat in Laiwu Pigs: Insights from Targeted Lipidomics and Transcriptomics. Agriculture 2024, 14, 658. https://doi.org/10.3390/agriculture14050658
Xu J, Wu T, Lam SM, Shui G, Yang S, Wang Y, Tao C. Heterogeneity of Intramuscular, Intermuscular, and Subcutaneous Fat in Laiwu Pigs: Insights from Targeted Lipidomics and Transcriptomics. Agriculture. 2024; 14(5):658. https://doi.org/10.3390/agriculture14050658
Chicago/Turabian StyleXu, Jian, Tianwen Wu, Sin Man Lam, Guanghou Shui, Shulin Yang, Yanfang Wang, and Cong Tao. 2024. "Heterogeneity of Intramuscular, Intermuscular, and Subcutaneous Fat in Laiwu Pigs: Insights from Targeted Lipidomics and Transcriptomics" Agriculture 14, no. 5: 658. https://doi.org/10.3390/agriculture14050658