
Gene Transcript Profiling in Sea Otters Post-Exxon Valdez Oil Spill: A Tool for Marine Ecosystem Health Assessment

Abstract
:1. Introduction
2. Materials and Methods
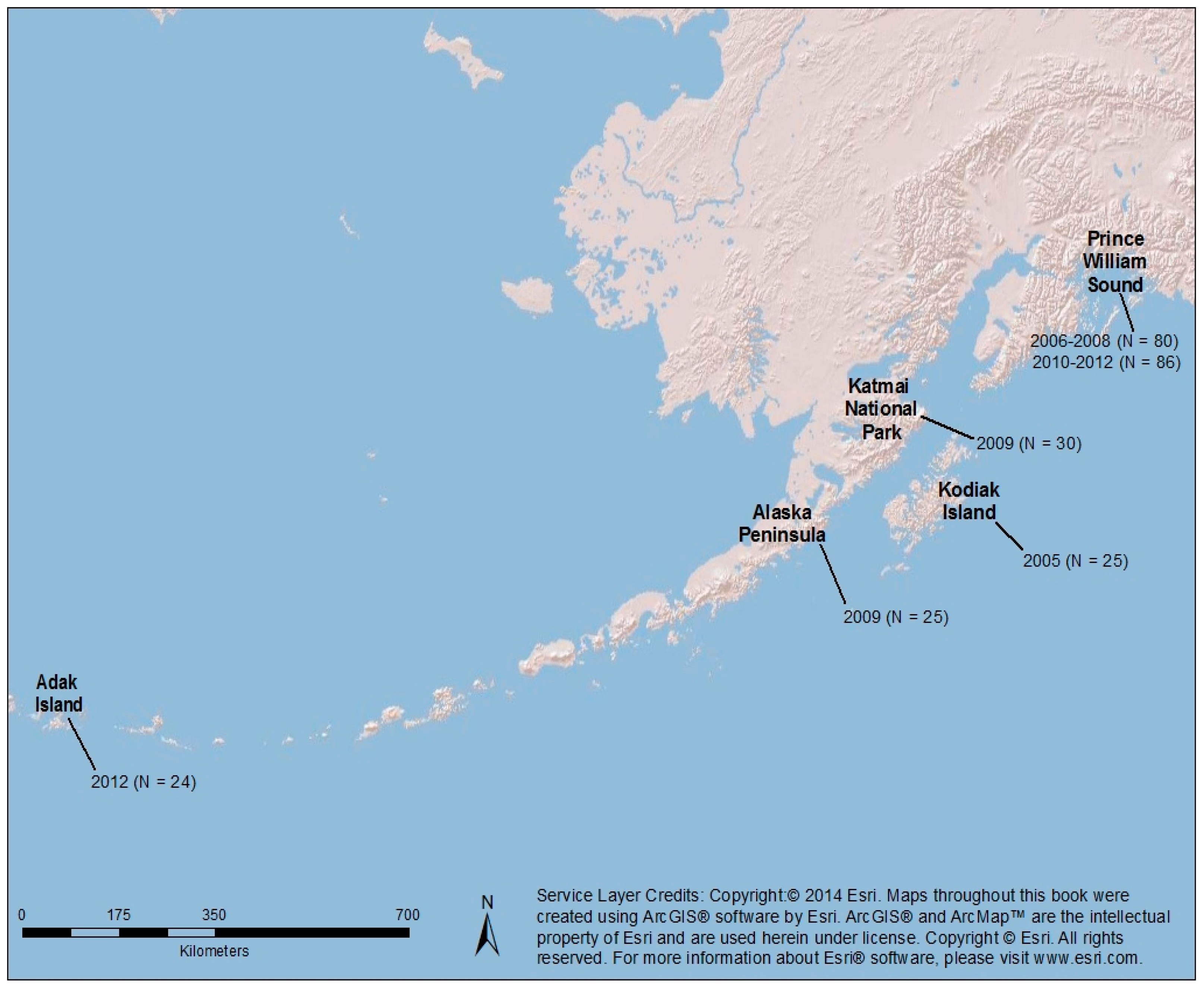
2.1. Sea Otter Samples
2.2. Blood and RNA Processing
2.3. Targeted Genes
2.4. Statistical Analyses
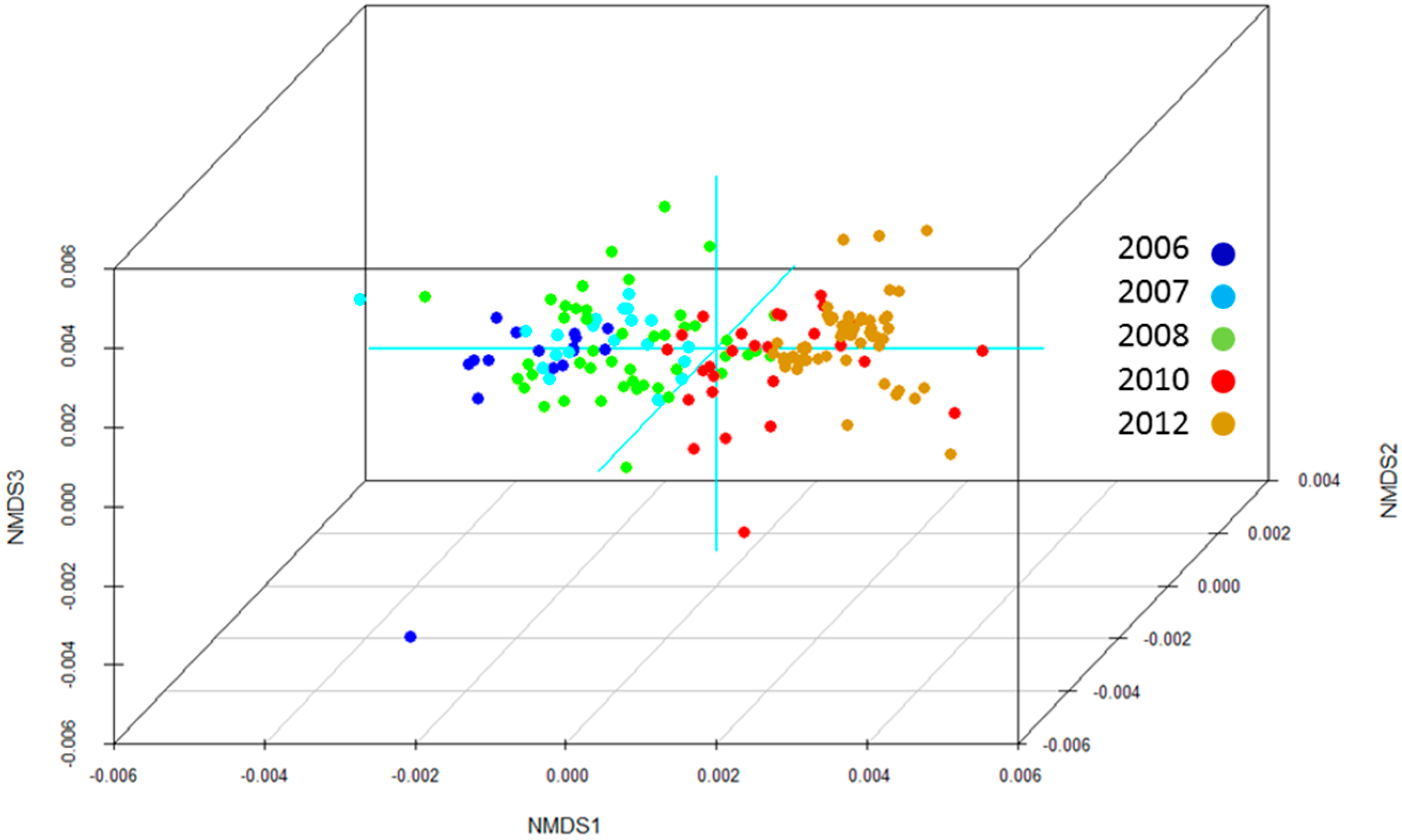
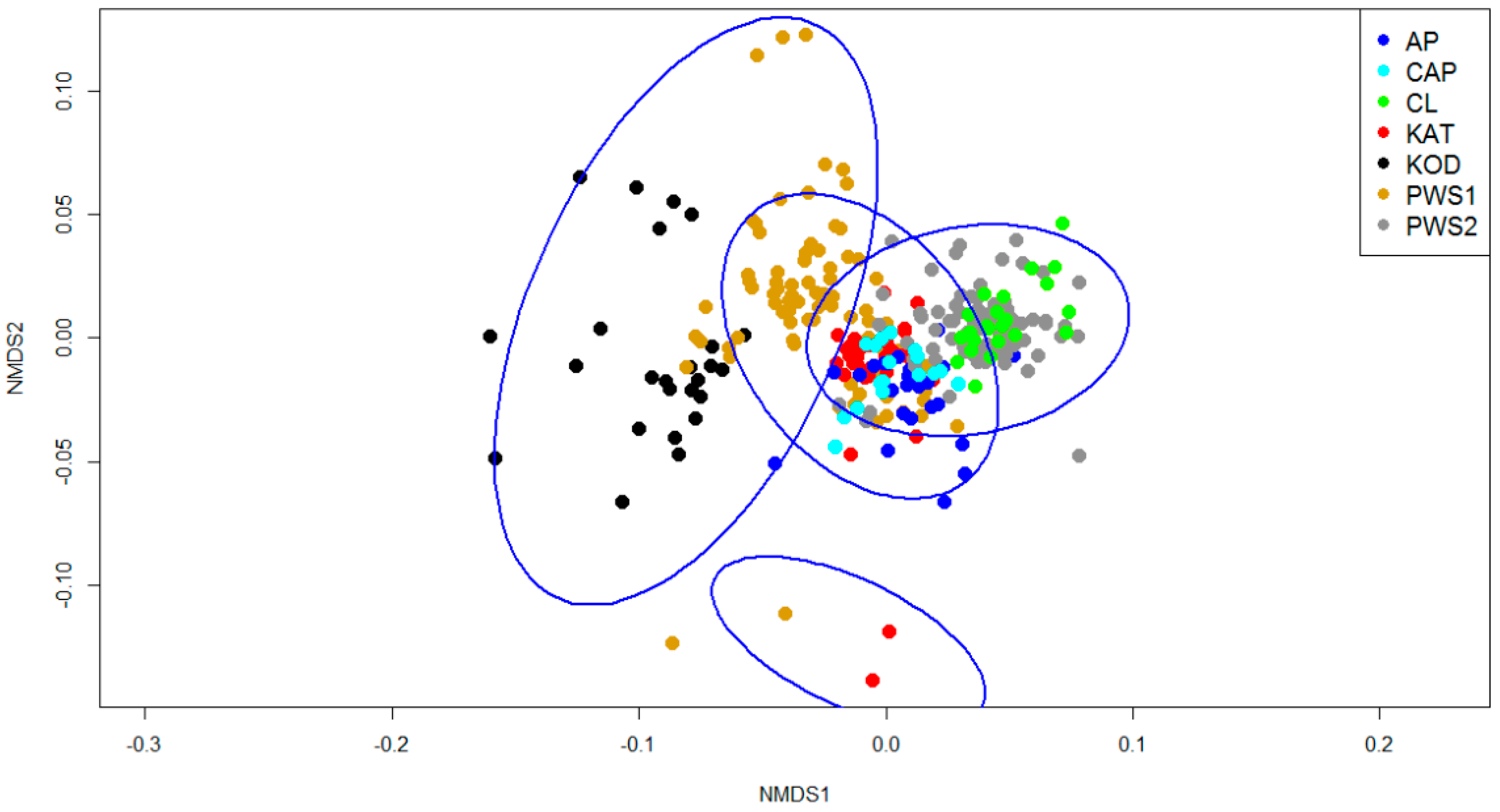
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bodkin, J.L.; Esler, D.; Rice, S.D.; Matkin, C.O.; Ballachey, B.E. The effects of spilled oil on coastal ecosystems: Lessons from the Exxon Valdez spill. In Coastal Conservation; Maslo, B., Lockwood, J.L., Eds.; Cambridge University Press: New York, NY, USA, 2014; pp. 311–346. [Google Scholar]
- Esler, D.; Trust, K.A.; Ballachey, B.E.; Iverson, S.A.; Lewis, T.L.; Rizzolo, D.J.; Mulcahy, D.M.; Miles, A.K.; Woodin, B.R.; Stegeman, J.J.; et al. Cytochrome P4501 A biomarker indication of oil exposure in harlequin ducks up to 20 years after the Exxon Valdez oil spill. Environ. Toxicol. Chem. 2010, 29, 1138–1145. [Google Scholar] [PubMed]
- Esler, D.; Ballachey, B.E.; Trust, K.A.; Iverson, S.A.; Reed, J.A.; Miles, A.K.; Henderson, J.D.; Woodin, B.R.; Statesman, J.J.; McAdie, M.; et al. Cytochrome P4501 A biomarker indication of the timeline of chronic exposure of Barrow’s goldeneyes to residual Exxon Valdez oil. Mar. Pollut. Bull. 2011, 62, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Monson, D.H.; Doak, D.F.; Ballachey, B.E.; Bodkin, J.L. Could residual oil from the Exxon Valdez spill create a long-term population ‘sink’ for sea otters in Alaska? Ecol. Appl. 2011, 21, 2917–2932. [Google Scholar] [CrossRef]
- Bowen, L.; Miles, A.K.; Murray, M.; Haulena, M.; Tuttle, J.; Van Bonn, W.; Adams, L.; Bodkin, J.L.; Ballachey, B.E.; Estes, J.A.; et al. Gene transcription in sea otters (Enhydra. lutris); emerging diagnostics in marine mammal and ecosystem health. Mol. Ecol. Resour. 2012, 12, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Miles, A.K.; Bowen, L.; Ballachey, B.E.; Bodkin, J.L.; Murray, M.; Estes, J.L.; Keister, R.A.; Stott, J.L. Variation in transcript profiles in sea otters (Enhydra lutris) from Prince William Sound, Alaska and clinically normal reference otters. Mar. Ecol. Prog. Ser. 2012, 451, 201–212. [Google Scholar] [CrossRef]
- Bodkin, J.L.; Ballachey, B.E.; Coletti, H.A.; Esslinger, G.G.; Kloecker, K.A.; Rice, S.D.; Reed, J.A.; Monson, D.H. Long-term effects of the Exxon Valdez oil spill—Sea otter foraging in the intertidal as a pathway of exposure to lingering oil. Mar. Ecol. Prog. Ser. 2012, 447, 273–287. [Google Scholar] [CrossRef]
- Ballachey, B.E.; Monson, D.H.; Esslinger, G.G.; Kloecker, K.; Bodkin, J.; Bowen, L.; Miles, A.K. 2013 Update on Sea Otter Studies to Assess Recovery from the 1989 Exxon Valdez Oil Spill, Prince William Sound, Alaska: U.S. Geological Survey Open-File Report 2014–1030, 40p. Available online: http://dx.doi.org/10.3133/ofr20141030 (accessed on 20 May 2016).
- Ballachey, B.E.; Bodkin, J.L.; Esler, D.; Rice, S.D. Lessons from the 1989 Exxon Valdez oil spill: A biological perspective. In Impacts of Oil Spill Disasters on Marine Habitats and Fisheries in North America; Alford, J.B., Peterson, M.S., Green, C.C., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 181–198. [Google Scholar]
- Harwell, M.A.; Gentile, J.H.; Johnson, C.B.; Garshelis, D.L.; Parker, K.R. A Quantitative Ecological Risk Assessment of the Toxicological Risks from Exxon Valdez Subsurface Oil Residues to Sea Otters at Northern Knight Island, Prince William Sound, Alaska. Hum. Ecol. Risk Assess. 2010, 16, 727–761. [Google Scholar] [CrossRef] [PubMed]
- Ballachey, B.E.; Bodkin, J.L.; Monson, D.H. Quantifying long-term risks to sea otters from the 1989 ‘Exxon Valdez’ oil spill: Reply to Harwell and Gentile (2013). Mar. Ecol. Prog. Ser. 2013, 480, 297–301. [Google Scholar] [CrossRef]
- Harwell, M.A.; Gentile, J.H. Assessing risks to sea otters and the Exxon Valdez oil spill: New scenarios, attributable risk, and recovery. Hum. Ecol. Risk Assess. 2014, 20, 889–916. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.H.; Rice, S.D.; Short, J.W.; Esler, D.; Bodkin, J.L.; Ballachey, B.E.; Irons, D.B. Long-term ecosystem response to the Exxon Valdez oil spill. Science 2003, 302, 2082–2086. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.; Dunn, R.T. Concise review: Gene expression applied to toxicology. Toxicol. Sci. 1999, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, K.; Turteltaub, K.; Bankaitis-Davis, D.; Gerren, R.; Siconolfi, L.; Storm, K.; Cheronis, J.; Trollinger, D.; Macejak, D.; Tryon, V.; et al. Limited dynamic range of immune response gene expression observed in healthy blood donors using RT-PCR. J. Mol. Med. 2006, 12, 185–195. [Google Scholar]
- Poynton, H.C.; Vulpe, C.D. Ecotoxicogenomics: Emerging technologies for emerging contaminants. J. Am. Water Resour. Assoc. 2009, 45, 83–96. [Google Scholar] [CrossRef]
- Monson, D.H.; McCormick, C.; Ballachey, B.E. Chemical anesthesia of northern sea otters (Enhydra lutris)—Results of past field studies. J. Zoo Wildl. Med. 2001, 32, 181–189. [Google Scholar] [PubMed]
- Bowen, L.; Schwartz, J.; Aldridge, B.; Riva, F.; Miles, A.K.; Mohr, F.C.; Stott, J.L. Differential gene expression induced by exposure of captive mink to fuel oil—A model for the sea otter. EcoHealth 2007, 4, 298–309. [Google Scholar] [CrossRef]
- Bommer, U.A.; Thiele, B.J. The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol. 2004, 36, 379–385. [Google Scholar] [CrossRef]
- Tuynder, M.; Fiucci, G.; Prieur, S.; Lespagnol, A.; Geant, A.; Beaucourt, S.; Duflaut, D.; Besse, S.; Susini, L.; Cavarelli, J.; et al. Translationally controlled tumor protein is a target of tumor reversion. Proc. Natl. Acad. Sci. USA 2004, 101, 15364–15369. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Geng, Y.; Xu, W.; Wu, Y.; He, F.; Shu, W.; Huang, M.; Du, H.; Li, M. The role of translationally controlled tumor protein in tumor growth and metastasis of colon adenocarcinoma cells. J. Proteome Res. 2010, 9, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Raisuddin, S.; Kwok, K.W.H.; Leung, K.M.Y.; Schlenk, D.; Lee, J. The copepod Tigriopus—A promising marine model organism for ecotoxicology and environmental genomics. Aquat. Toxicol. 2007, 83, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Song, Y.; Qiu, X.; Sun, T.; Ackland, M.L.; Zhang, W. Annetocin and TCTP expressions in the earthworm Eisenia fetida exposed to PAHs in artificial soil. Ecotoxicol. Environ. Saf. 2008, 71, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Goldsby, R.A.; Kindt, T.J.; Osborne, B.A.; Kuby, J. Immunology, 5th ed.; W.H. Freeman and Company: New York, NY, USA, 2003. [Google Scholar]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Jenne, D.E.; Tschopp, J. Molecular structure and functional characterization of a human complement cytolysis inhibitor found in blood and seminal plasma—Identity to sulfated glycoprotein 2, a constituent of rat testis fluid. Proc. Natl. Acad. Sci. USA 1989, 86, 7123–7127. [Google Scholar] [CrossRef] [PubMed]
- Zeytun, A.; McKallip, R.J.; Fisher, M.; Camacho, I.; Nagarkatti, M.; Nagarkatti, P.S. Analysis of 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced gene expression profile in vivo using pathway-specific cDNA arrays. Toxicology 2002, 23, 241–260. [Google Scholar] [CrossRef]
- Oesch-Bartlomowicz, B.; Huelster, A.; Wiss, O.; Antoniou-Lipfert, P.; Dietrich, C.; Arand, M.; Weiss, C.; Bockamp, E.; Oesch, F. Aryl hydrocarbon receptor activation by cAMP vs. dioxin: Divergent signaling pathways. Proc. Natl. Acad. Sci. USA 2005, 102, 9218–9223. [Google Scholar] [CrossRef] [PubMed]
- Quintana, F.J.; Basso, A.S.; Iglesias, A.H.; Korn, T.; Farez, M.F.; Bettelli, E.; Caccamo, M.; Oukka, M.; Weiner, H.L. Control of T(reg) and T(H)17 cell differentiation by the aryl hydrocarbon receptor. Nature 2008, 453, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, M.; Veldhoen, N.; Dangerfield, N.; Jeffries, S.; Helbing, C.C.; Ross, P.S. PCB-related alteration of thyroid hormones and thyroid hormone receptor gene expression in free-ranging harbor seals (Phoca vitulina). Environ. Health Perspect. 2006, 114, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.J.; O’Malley, B.W. Molecular mechanisms of action of steroid/thyroid receptor superfamily members. Annu. Rev. Biochem. 1994, 63, 451–486. [Google Scholar] [CrossRef] [PubMed]
- Iwama, G.K.; Mathilakath, M.V.; Forsyth, R.B.; Ackerman, P.A. Heat shock proteins and physiological stress in fish. Am. Zool. 1999, 39, 901–909. [Google Scholar] [CrossRef]
- Tsan, M.; Gao, B. Cytokine function of heat shock proteins. Am. J. Physiol. Cell Physiol. 2004, 286, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Krumm, B.; Meng, X.; Li, Y.; Xiang, Y.; Deng, J. Structural basis for antagonism of Human interleukin 18 by poxvirus interleukin 18-binding protein. Proc. Natl. Acad. Sci. USA 2008, 105, 20711–20715. [Google Scholar] [CrossRef] [PubMed]
- Beineke, A.; Siebert, U.; Muller, G.; Baumgartner, W. Increased blood interleukin-10 mRNA levels in diseased free-ranging harbor porpoises (Phocoena phocoena). Vet. Immunol. Immunopathol. 2007, 115, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Rigopoulou, E.I.; Abbott, W.G.; Haigh, P.; Naoumov, N.V. Blocking of interleukin-10 receptor-a novel approach to stimulate T-helper cell type 1 responses to hepatitis C virus. Clin. Immunol. 2005, 117, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Bowen, L.; Aldridge, B.; Miles, A.K.; Stott, J.L. Expressed MHC class II genes in sea otters (Enhydra lutris) from geographically disparate populations. Tissue Antigens 2006, 67, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wegner, K.M.; Kalbe, M.; Rauch, G.; Kurtz, J.; Schaschl, H.; Reusch, T.B.H. Genetic variation in MHC class II expression and interactions with MHC sequence polymorphism in three-spined sticklebacks. Mol. Ecol. 2006, 15, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.C.; Stockinger, B. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 2008, 453, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Tumpey, T.M.; Szretter, K.J.; Van Hoeven, N.; Katz, J.M.; Kochs, G.; Haller, O.; Garcia-Sister, A.; Staeheli, P. The Mx1 gene protects mice against the pandemic 1918 and highly lethal human H5 N1 influenza viruses. J. Virol. 2007, 81, 10818–10821. [Google Scholar] [CrossRef] [PubMed]
- Kibenge, M.J.T.; Munir, K.; Kibenge, F.S.B. Constitutive expression of Atlantic salmon Mx1 protein in CHSE-214 cells confers resistance to infectious salmon Anaemia virus. J. Virol. 2005, 2, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; ISBN 3-900051-07-0. R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: http://www.R-project.org/ (accessed on 20 May 2016).
- Monson, D.H.; Doak, B.E.; Johnson, A.; Bodkin, J.L. Long-term impact of the Exxon Valdez oil spill on sea otters, assessed through age-dependent mortality patterns. Proc. Natl. Acad. Sci. USA 2000, 97, 6562–6567. [Google Scholar] [CrossRef] [PubMed]
- U.S. Fish and Wildlife Service. Northern Sea Otter (Enhydra lutris kenyoni) Southwest Alaska Stock Report; U.S. Fish and Wildlife Service: Anchorage, AK, USA, 2014. [Google Scholar]
- Coletti, H.A.; Bodkin, J.L.; Monson, D.H.; Dean, T.A.; Ballachey, B.E. Engaging form and function to detect and infer cause of change in an Alaska marine ecosystem. Ecosphere 2016. submitted. [Google Scholar]
- Saucillo, D.C.; Gerriets, V.A.; Sheng, J.; Rathmell, J.C.; MacIver, N.J. Leptin metabolically licenses T cells for activation to link nutrition and immunity. J. Immunol. 2014, 192, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.B.; Hopkins, W.A.; Mydlarz, L.D.; Rohr, J.R. The effects of anthropogenic global changes on immune functions and disease resistance. Ann. N. Y. Acad. Sci. 2010, 1195, 129–148. [Google Scholar] [CrossRef] [PubMed]
- Bodkin, J.L.; U.S. Geological Survey, Port Townsend, WA, USA. Unpublished work. 2016.
- Estes, J.A.; Tinker, M.T.; Williams, T.M.; Doak, D.F. Killer whale predation on sea otters linking oceanic and nearshore ecosystems. Science 1998, 282, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Menzel, R.; Swain, S.C.; Hoess, S.; Claus, E.; Menzel, S.; Steinberg, C.E.W.; Reifferscheid, G.; Sturzenbaum, S.R. Gene expression profiling to characterize sediment toxicity—A pilot study using Caenorhabditis elegans whole genome microarrays. BMC Genomics 2009, 10, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, C.E.; Sturzenbaum, S.R.; Menzel, R. Genes and environment—Striking the fine balance between sophisticated biomonitoring and true functional environmental genomics. Sci. Total Environ. 2008, 400, 142–161. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kemadjou, J.R.; Zinsmeister, C.; Bauer, M.; Legradi, J.; Muller, F.; Pankratz, M.; Jakel, J.; Strahle, U. Transcriptional profiling reveals barcode-like toxicogenomic responses in the zebrafish embryo. Genome Biol. 2007, 8, R227. [Google Scholar] [CrossRef] [PubMed]
- Poynton, H.C.; Zuzow, R.; Loguinov, A.V.; Perkins, E.J.; Vulpe, C.D. Gene expression profiling in Daphnia magna, Part II: Validation of a copper specific gene expression signature with effluent from two copper mines in California. Environ. Sci. Technol. 2008, 42, 6257–6263. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Ma, Q.; Whitlock, J.P., Jr. Down-regulation of major histocompatibility complex Q1 b gene expression by 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 1997, 272, 29614–29619. [Google Scholar] [CrossRef] [PubMed]
- Ballachey, B.E.; Bodkin, J.L. Challenges to Sea Otter Recovery and Conservation. In Sea Otter Conservation; Larson, S.E., Bodkin, J.L., VanBlaricom, G.R., Eds.; Elsevier: London, UK, 2015; pp. 63–88. [Google Scholar]
- Bowen, L.; Miles, A.K.; Ballachey, B.E.; Esler, D. Long-term Monitoring Program—Evaluating Chronic Exposure of Harlequin Ducks and Sea Otters to Lingering Exxon Valdez Oil in Western Prince William Sound. In Exxon Valdez Oil Spill Trustee Council Restoration Project Final Report (Project 14120114-Q); U.S. Geological Survey, Alaska Science Center: Anchorage, AK, USA, 2014. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Function |
|---|---|
| HDC | The HDCMB21P gene codes for a translationally-controlled tumor protein (TCTP) implicated in cell growth, cell cycle progression, malignant transformation, tumor progression and in the protection of cells against various stress conditions and apoptosis [19,20,21]. Environmental triggers may be responsible for population-based up-regulation of HDC. HDC transcription is known to increase with exposure to carcinogenic compounds, such as polycyclic aromatic hydrocarbons [18,22,23]. |
| COX2 | Cyclooxygenase-2 catalyzes the production of prostaglandins that are responsible for promoting inflammation [24]. Cox2 is responsible for the conversion of arachidonic acid to prostaglandin H2, a lipoprotein critical to the promotion of inflammation [25]. Upregulation of Cox2 is indicative of cellular or tissue damage and an associated inflammatory response. |
| CYT | The complement cytolysis inhibitor protects against cell death [26]. Upregulation of CYT is indicative of cell or tissue death. |
| AHR | The aryl hydrocarbon receptor responds to classes of environmental toxicants, including polycyclic aromatic hydrocarbons, polyhalogenated hydrocarbons, dibenzofurans and dioxin [27]. Depending on the ligand, AHR signaling can modulate T-regulatory (TREG) (immune-suppressive) or T-helper type 17 (TH17) (pro-inflammatory) immunologic activity [28,29]. |
| THRβ | The thyroid hormone receptor beta can be used as a mechanistically-based means of characterizing the thyroid-toxic potential of complex contaminant mixtures [30]. Thus, increases in THRβ transcription may indicate exposure to organic compounds, including PCBs, and associated potential health effects, such as developmental abnormalities and neurotoxicity [30,31]. |
| HSP 70 | The heat shock protein 70 is produced in response to thermal or other stress, including hyperthermia, oxygen radicals, heavy metals and ethanol [32,33]. |
| IL-18 | Interleukin-18 is a pro-inflammatory cytokine [24]. IL-18 plays an important role in inflammation and host defense against microbes [34]. |
| IL-10 | Interleukin-10 is an anti-inflammatory cytokine [24]. Levels of IL-10 have been correlated with the relative health of free-ranging harbor porpoises, e.g., increased amounts of IL-10 correlated with chronic disease, whereas the cytokine was relatively reduced in apparently fit animals experiencing acute disease [35]. Association of IL-10 transcription with chronic disease has also been documented in humans [36]. |
| DRB | A component of the major histocompatibility complex, the DRB class II gene is responsible for the binding and presentation of processed antigen to TH lymphocytes, thereby facilitating the initiation of an immune response [24,37]. Upregulation of MHC genes has been positively correlated with parasite load [37], whereas downregulation of MHC has been associated with contaminant exposure [38,39]. |
| Mx1 | The Mx1 gene responds to viral infection [40]. Vertebrates have an early strong innate immune response against viral infection, characterized by the induction and secretion of cytokines that mediate an antiviral state, leading to the upregulation of the MX-1 gene [41]. |
| Group | Gene | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HDC | COX2 | CYT | AHR | THRβ | HSP70 | IL18 | IL10 | DRB | MX1 | |
| CAP | 5.90 ad (5.02–6.94) | 6.78 abcd (6.02–7.64) | 2.41 (1.91–3.04) | 11.01 abe (10.56–11.48) | 13.30 a (12.49–14.56) | 9.62 ab (8.74–10.59) | 1.65 acde (1.05–2.60) | 13.70 a (13.01–14.44) | −0.33 (−0.86–0.21) | 10.99 ab (9.95–12.15) |
| AP | 6.26 a (5.47–7.16) | 6.60 abcd (5.92–7.36) | 1.90 (1.45–2.52) | 10.67 ab (9.94–11.46) | 13.32 a (12.19–14.56) | 8.58 abd (7.97–9.23) | 1.68 acde (1.30–2.17) | 13.21 a (12.21–14.30) | −0.91 a (−1.57–−0.26) | 12.61 a (11.42–13.93) |
| KAT | 4.54 ab (4.06–5.08) | 7.68 bd (7.10–8.30) | 1.96 (1.54–2.50) | 10.36 ab (9.79–10.96) | 12.53 a (11.86–13.23) | 8.26 abd (7.56–9.04) | 2.78 ce (2.15–3.59) | 13.45 a (12.62–14.33) | −0.56 a (−1.09–−0.03) | 12.56 a (11.99–13.16) |
| KOD | −1.84 e (−2.33–−1.35) | 5.44 c (4.79–6.16) | 2.59 (2.04–3.28) | 8.80 d (8.09–9.57) | 9.50 c (8.62–10.46) | 5.48 d (4.86–6.18) | 5.19 b (4.61–5.85) | 9.03 c (8.26–9.87) | 1.29 (0.78–2.13) | 8.26 b (7.64–8.92) |
| CL | 10.30 c (10.06–10.54) | 9.45 e (8.91–10.02) | 1.53 (1.28–1.83) | 12.78 c (12.38–13.18) | 16.85 b (15.83–17.93) | 14.07 c (13.17–15.02) | 2.49 acde (2.06–3.01) | 22.09 b (20.85–23.40) | 0.43 b (0.22–0.84) | 16.89 c (15.36–18.57) |
| PWS1 | 4.01 bde (2.89–5.57) | 7.98 ad (7.60–8.38) | 1.88 (1.60–2.21) | 10.17 abd (9.75–10.62) | 11.35 ac (10.74–11.99) | 9.76 ab (9.17–10.39) | 1.60 d (1.24–2.07) | 13.34 a (12.77–13.94) | 1.08 b (0.80–1.44) | 10.41 b (10.05–10.78) |
| PWS2 | 8.94 c (8.46–9.46) | 9.30 e (8.96–9.65) | 1.62 (1.45–1.80) | 12.07 c (11.79–12.36) | 16.09 b (15.54–16.66) | 13.62 c (13.10–14.01) | 2.38 e (2.21–2.56) | 20.28 b (19.44–21.16) | −0.071 (−0.25–0.10) | 14.95 c (14.50–15.41) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowen, L.; Miles, A.K.; Ballachey, B.; Waters, S.; Bodkin, J. Gene Transcript Profiling in Sea Otters Post-Exxon Valdez Oil Spill: A Tool for Marine Ecosystem Health Assessment. J. Mar. Sci. Eng. 2016, 4, 39. https://doi.org/10.3390/jmse4020039
Bowen L, Miles AK, Ballachey B, Waters S, Bodkin J. Gene Transcript Profiling in Sea Otters Post-Exxon Valdez Oil Spill: A Tool for Marine Ecosystem Health Assessment. Journal of Marine Science and Engineering. 2016; 4(2):39. https://doi.org/10.3390/jmse4020039
Chicago/Turabian StyleBowen, Lizabeth, A. Keith Miles, Brenda Ballachey, Shannon Waters, and James Bodkin. 2016. "Gene Transcript Profiling in Sea Otters Post-Exxon Valdez Oil Spill: A Tool for Marine Ecosystem Health Assessment" Journal of Marine Science and Engineering 4, no. 2: 39. https://doi.org/10.3390/jmse4020039