
A Closer Look at Schlemm’s Canal Cell Physiology: Implications for Biomimetics


Abstract
:1. Introduction


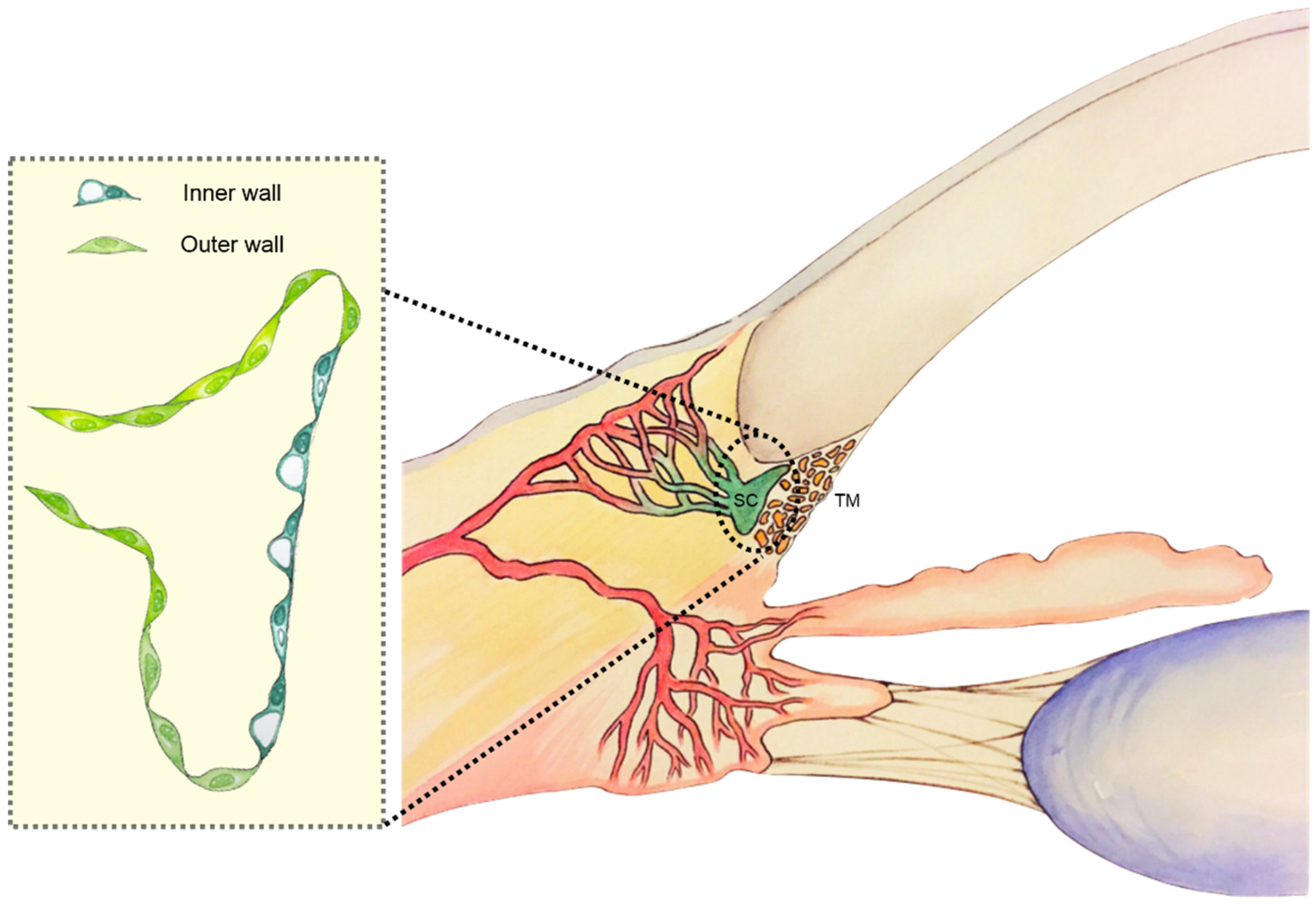{kind=link}
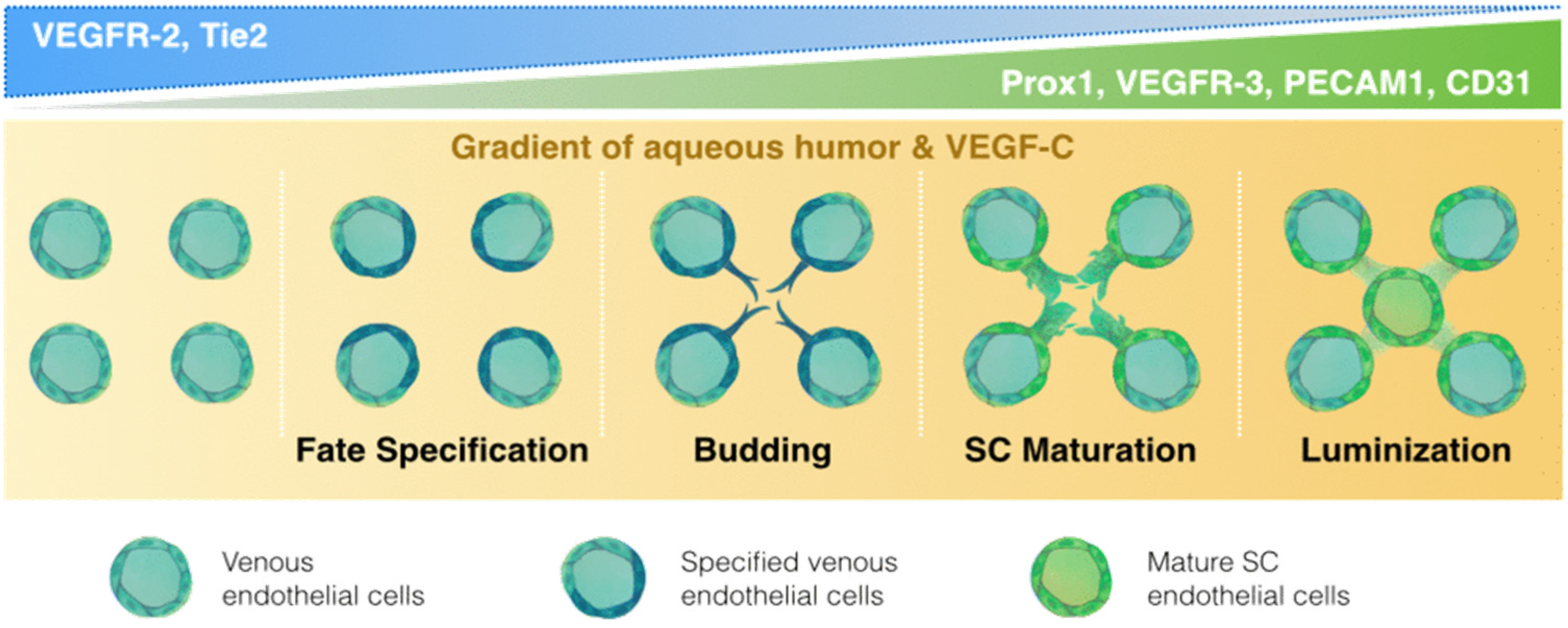{kind=link}
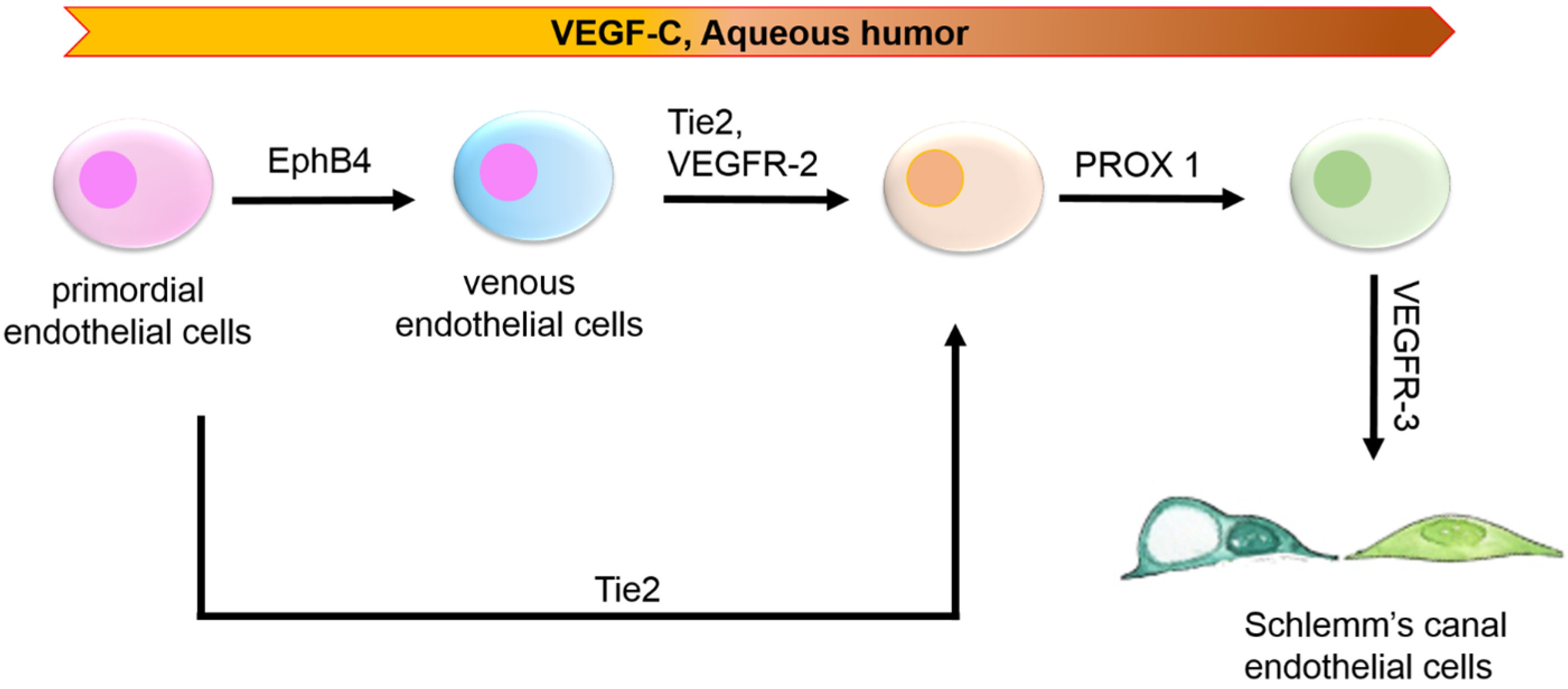{kind=link}
| Property | Inner Wall | Outer Wall |
|---|---|---|
| Morphology | Cobblestone appearance [15] | Smooth and flat [5], continuous basement membrane [17] |
| Discontinuous basement membrane [15,16] | ||
| Cell-specific marker | Zipper-like VE-cadherin [18] | Desmin |
| Reactivity to Factor VIII-related antigen [19] | ||
| Subcellular structure | Giant vacuoles [20], paracellular pores [21] | Weibel-Palade bodies [17] |
| Function | Aqueous humor filtration | Unknown |
| IOP homeostasis [5,22,23] |
2. Embryological Origin: Of Lymph or Blood?

| Lineage | Development | Progenitors | Budding | Lumenization/Sac Formation | Separation from Venous Vasculature |
|---|---|---|---|---|---|
| Lymphatic | Embryonic | PROX1 [32], Sox18 [33], COUP-FII [34], VE-cadherin [35] | PDPN [36], VEFGR3 [37], CCBE1 [38], NRP2 [39], RAC1 [40], LYVE-1 [41] | NFATC1 [42], GATA2 [43,44], Calcr1 [45], Ramp2 [45], TIE1 [46] | Syk [47], SLP76 [47], Runx1 [48], PDPN [36], Meis1 [49], Clec2 [50,51], CXADR [52] |
| Schlemm’s Canal | Postnatal | VEGFR-2, TIE 2 [2,28] | PROX1 [2,28,55] | VEFGR-3 [2,28] | PECAM1, VEFGR-3 [2,18,28] |
3. Schlemm’s Canal Anatomy
3.1. Macroarchitecture
3.2. Microarchitecture
4. Characteristics of Human Schlemm’s Canal Cells
| Molecular/Cellular Characteristics | Schlemm’s Canal Endothelium a | Lymphatic Endothelium | Vascular Endothelium |
|---|---|---|---|
| Sox18 | – | + [33] | – |
| VEGFR-2 | + [2,28] | – | + [82] |
| VEGFR-3 | + [2,28] | + [38] | + [83] |
| PROX1 | + [2,28,53] | + [34] | – |
| CCL21 | + [2,28] | – | – |
| Itga9 | + [2] | – | – |
| Collagen IV | + [2] | – | – |
| PECAM1 | + [18] | – | + [84] |
| VE-cadherin | + [18] | + [35] | + [85] |
| Endomucin | + [53] | – | – |
| Foxc2 | + [2,28] | – | – |
| LYVE-1 | – | + [41] | – |
| Podoplanin | – | + [36] | – |
| vWF | + [17] | – | + [86] |
| Wiebel-Palade bodies | + [17] | – | + [86] |
| Endothelial monolayer | continuous [5] | continuous [29,87] | continuous |
| Basement membrane | discontinuous [5] | discontinuous [29,87] | continuous |
| Basal-to-apical Flow | + [5,88] | + [87,89] | – |
5. Biomechanics
6. Perspective on Schlemm’s Canal Engineering
6.1. Criteria for a 3D in Vitro Model of the Schlemm’s Canal Inner Wall
- (1)
- Express key cell-specific markers, necessary for the endothelial integrity and mechanosensing;
- (2)
- Display both paracellular and transcellular pathways vital to aqueous outflow function of the SC;
- (3)
- Mimic the in vivo cellular micro architecture with respect to morphological features such as a cellular dimensions or surface area of cell-cell interactions within the cultured monolayer, the spatial distribution of subcellular organelles (vacuoles), the complexity of tight junctional strands;
- (4)
- Allow for ease of culture using phenotypically stable cell lines to facilitate high throughput screening. Thus, a well-characterized in vitro 3D model of the HSCIW would provide a system in which to study and understand the physiology, biomechanics, outflow functions, physiological drug responses as well as pathological processes in glaucoma.
6.2. Potential Sources of Human Schlemm’s Canal Inner Wall Endothelial Cells

6.3. Biomaterials for 3D Culture of Human Schlemm’s Canal Inner Wall Cells
6.4. Soluble Factors for Directed Schlemm’s Canal Cell Differentiation
6.5. Dynamic 3D Culture
7. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| Calcr1 | Calcitonin Receptor 1 |
| CCBE1 | Collagen and calcium-binding EGF domain-containing protein 1 |
| CCL21 | Chemokine (C-C motif) ligand 21 |
| Clec2 | C-type lectin-like receptor 2 |
| COUP-FTII | Chicken ovalbumin upstream promoter-transcription factor 2 |
| CXADR | Coxsackie virus and adenovirus receptor |
| Foxc2 | Forkhead box protein C2 |
| GATA2 | GATA binding protein 2 |
| Itga9 | Integrin alpha-9 |
| LYVE-1 | Lymphatic vessel endothelial hyaluronan receptor |
| Meis1 | Meis homeobox 1 |
| Nfatc1 | Nuclear factor of activated T-cells, cytoplasmic 1 |
| NRP2 | Neuropilin 2 |
| PDPN | Podoplanin |
| PECAM1 | Platelet endothelial cell adhesion molecule |
| PROX1 | Prospero homeobox protein 1 |
| RAC1 | Ras-related C3 botulinum toxin substrate 1 |
| Ramp2 | Receptor activity modifying protein 2 |
| Runx1 | Runt-related transcription factor 1 |
| SLP76 | Lymphocyte cytosolic protein 2 |
| Syk | Spleen tyrosine kinase |
| TIE1 | Tunica interna endothelial cell kinase 1 |
| TIE2 | Tunica interna endothelial cell kinase 2 |
| VE-cadherin | Vascular endothelial cadherin |
| VEFGR-2 | Vascular endothelial growth factor 2 |
| VEFGR-3 | Vascular endothelial growth factor 3 |
| vWF | Von Willebrand factor |
References
- Mansouri, K.; Shaarawy, T. Update on Schlemm’s canal based procedures. Middle East Afr. J. Ophthalmol. 2015, 22, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Aspelund, A.; Tammela, T.; Antila, S.; Nurmi, H.; Leppanen, V.M.; Zarkada, G.; Stanczuk, L.; Francois, M.; Makinen, T.; Saharinen, P.; et al. The Schlemm’s canal is a VEGF-C/VEGFR-3-responsive lymphatic-like vessel. J. Clin. Investig. 2014, 124, 3975–3986. [Google Scholar] [CrossRef] [PubMed]
- Ashpole, N.E.; Overby, D.R.; Ethier, C.R.; Stamer, W.D. Shear stress-triggered nitric oxide release from Schlemm’s canal cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 8067–8076. [Google Scholar] [CrossRef] [PubMed]
- Lutjen-Drecoll, E.; Rohen, J.W. [Endothelial studies of the Schlemm’s canal using silver-impregnation technic]. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 1970, 180, 249–266. [Google Scholar] [PubMed]
- Ethier, C.R. The inner wall of Schlemm’s canal. Exp. Eye Res. 2002, 74, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Karl, M.O.; Fleischhauer, J.C.; Stamer, W.D.; Peterson-Yantorno, K.; Mitchell, C.H.; Stone, R.A.; Civan, M.M. Differential P1-purinergic modulation of human Schlemm’s canal inner-wall cells. Am. J. Physiol. Cell Physiol. 2005, 288, C784–C794. [Google Scholar] [CrossRef] [PubMed]
- Vranka, J.A.; Kelley, M.J.; Acott, T.S.; Keller, K.E. Extracellular matrix in the trabecular meshwork: Intraocular pressure regulation and dysregulation in glaucoma. Exp. Eye Res. 2015, 133, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Acott, T.S.; Kelley, M.J. Extracellular matrix in the trabecular meshwork. Exp. Eye Res. 2008, 86, 543–561. [Google Scholar] [CrossRef] [PubMed]
- Maepea, O.; Bill, A. Pressures in the juxtacanalicular tissue and Schlemm’s canal in monkeys. Exp. Eye Res. 1992, 54, 879–883. [Google Scholar] [CrossRef]
- Johnson, M. What controls aqueous humour outflow resistance? Exp. Eye Res. 2006, 82, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.C.; Kamm, R.D. The role of Schlemm’s canal in aqueous outflow from the human eye. Investig. Ophthalmol. Vis. Sci. 1983, 24, 320–325. [Google Scholar]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Braakman, S.T.; Pedrigi, R.M.; Read, A.T.; Smith, J.A.E.; Stamer, W.D.; Ethier, C.R.; Overby, D.R. Biomechanical strain as a trigger for pore formation in Schlemm’s canal endothelial cells. Exp. Eye Res. 2014, 127, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Zhou, E.H.; Tambe, D.; Chen, B.; Lavoie, T.; Dowell, M.; Simeonov, A.; Maloney, D.J.; Marinkovic, A.; Tschumperlin, D.J.; et al. High-throughput screening for modulators of cellular contractile force. Integr. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Tripathi, R.C.; Tripathi, B.J. Morphology of the aqueous outflow pathway. Microsc. Res. Tech. 1996, 33, 336–367. [Google Scholar] [CrossRef]
- Tamm, E.R. The trabecular meshwork outflow pathways: Structural and functional aspects. Exp. Eye Res. 2009, 88, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, T.; Bill, A.; Ichinohasama, R.; Ishida, T. Aspects of the development of Schlemm’s canal. Exp. Eye Res. 1992, 55, 479–488. [Google Scholar] [CrossRef]
- Heimark, R.L.; Kaochar, S.; Stamer, W.D. Human Schlemm’s canal cells express the endothelial adherens proteins, VE-cadherin and PECAM-1. Curr. Eye Res. 2002, 25, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, T.; Bill, A. Platelet aggregation on the endothelium of Schlemm’s canal. Exp. Eye Res. 1994, 59, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, M.A.; Grant, W.M. Pressure-dependent changes in structures of the aqueous outflow system of human and monkey eyes. Am. J. Ophthalmol. 1973, 75, 365–383. [Google Scholar] [CrossRef]
- Ethier, C.R.; Coloma, F.M.; Sit, A.J.; Johnson, M. Two pore types in the inner-wall endothelium of Schlemm’s canal. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2041–2048. [Google Scholar]
- Goel, M.; Picciani, R.G.; Lee, R.K.; Bhattacharya, S.K. Aqueous humor dynamics: A review. Open Ophthalmol. J. 2010, 4, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Braakman, S.T.; Read, A.T.; Chan, D.W.; Ethier, C.R.; Overby, D.R. Colocalization of outflow segmentation and pores along the inner wall of Schlemm’s canal. Exp. Eye Res. 2015, 130, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Smelser, G.K.; Ozanics, V. The development of the trabecular meshwork in primate eyes. Am. J. Ophthalmol. 1971, 71, 366–385. [Google Scholar] [CrossRef]
- Foets, B.; van den Oord, J.; Engelmann, K.; Missotten, L. A comparative immunohistochemical study of human corneotrabecular tissue. Graefes Arch. Clin. Exp. Ophthalmol. 1992, 230, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Wulle, K.G. Electron microscopic observations of the development of Schlemm’s canal in the human eye. Trans. Am. Acad. Ophthalmol. Otolaryngol. 1968, 72, 765–773. [Google Scholar] [PubMed]
- Ramirez, J.M.; Ramirez, A.I.; Salazar, J.J.; Rojas, B.; de Hoz, R.; Trivino, A. Schlemm’s canal and the collector channels at different developmental stages in the human eye. Cells Tissues Organs 2004, 178, 180–185. [Google Scholar] [PubMed]
- Park, D.Y.; Lee, J.; Park, I.; Choi, D.; Lee, S.; Song, S.; Hwang, Y.; Hong, K.Y.; Nakaoka, Y.; Makinen, T.; et al. Lymphatic regulator PROX1 determines Schlemm’s canal integrity and identity. J. Clin. Investig. 2014, 124, 3960–3974. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Oliver, G. Development of the mammalian lymphatic vasculature. J. Clin. Investig. 2014, 124, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Srinivasan, R.S. Endothelial cell plasticity: How to become and remain a lymphatic endothelial cell. Development 2010, 137, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Hagerling, R.; Pollmann, C.; Andreas, M.; Schmidt, C.; Nurmi, H.; Adams, R.H.; Alitalo, K.; Andresen, V.; Schulte-Merker, S.; Kiefer, F. A novel multistep mechanism for initial lymphangiogenesis in mouse embryos based on ultramicroscopy. EMBO J. 2013, 32, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.K.; Detmar, M. PROX1, master regulator of the lymphatic vasculature phenotype. Cell Tissue Res. 2003, 314, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Francois, M.; Caprini, A.; Hosking, B.; Orsenigo, F.; Wilhelm, D.; Browne, C.; Paavonen, K.; Karnezis, T.; Shayan, R.; Downes, M.; et al. Sox18 induces development of the lymphatic vasculature in mice. Nature 2008, 456, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Yoshimatsu, Y.; Morishita, Y.; Miyazono, K.; Watabe, T. Coup-TFII regulates the functions of PROX1 in lymphatic endothelial cells through direct interaction. Genes Cells 2009, 14, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; García-Verdugo, J.M.; Soriano-Navarro, M.; Srinivasan, R.S.; Scallan, J.P.; Singh, M.K.; Epstein, J.A.; Oliver, G. Lymphatic endothelial progenitors bud from the cardinal vein and intersomitic vessels in mammalian embryos. Blood 2012, 120, 2340–2348. [Google Scholar] [CrossRef] [PubMed]
- Breiteneder-Geleff, S.; Soleiman, A.; Kowalski, H.; Horvat, R.; Amann, G.; Kriehuber, E.; Diem, K.; Weninger, W.; Tschachler, E.; Alitalo, K.; et al. Angiosarcomas express mixed endothelial phenotypes of blood and lymphatic capillaries: Podoplanin as a specific marker for lymphatic endothelium. Am. J. Pathol. 1999, 154, 385–394. [Google Scholar] [CrossRef]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the FMS-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc. Natl. Acad. Sci. USA 1995, 92, 3566–3570. [Google Scholar] [CrossRef] [PubMed]
- Le Guen, L.; Karpanen, T.; Schulte, D.; Harris, N.C.; Koltowska, K.; Roukens, G.; Bower, N.I.; van Impel, A.; Stacker, S.A.; Achen, M.G.; et al. CCBE1 regulates VEGFC-mediated induction of VEGFR3 signaling during embryonic lymphangiogenesis. Development 2014, 141, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.J.; Chen, X.; Qin, J.; Hong, Y.K.; Tsai, M.J.; Tsai, S.Y. Direct transcriptional regulation of neuropilin-2 by COUP-TFII modulates multiple steps in murine lymphatic vessel development. J. Clin. Investig. 2010, 120, 1694–1707. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, G.; Jones, D.T.; Nye, E.; Sapienza, K.; Ramjuan, A.R.; Reynolds, L.E.; Robinson, S.D.; Kostourou, V.; Martinez, D.; Aubyn, D.; et al. Regulation of lymphatic-blood vessel separation by endothelial RAC1. Development 2009, 136, 4043–4053. [Google Scholar] [CrossRef] [PubMed]
- Banerji, S.; Ni, J.; Wang, S.-X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.M.; Greenberg, J.M.; Akeson, A.L. NFATC1 regulates lymphatic endothelial development. Mech. Dev. 2009, 126, 350–365. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.-C.; Hosoya, T.; Brandt, W.; Ku, C.-J.; Hosoya-Ohmura, S.; Camper, S.A.; Yamamoto, M.; Engel, J.D. Conditional GATA2 inactivation results in HSC loss and lymphatic mispatterning. J. Clin. Investig. 2012, 122, 3705–3717. [Google Scholar] [CrossRef] [PubMed]
- Kazenwadel, J.; Secker, G.A.; Liu, Y.J.; Rosenfeld, J.A.; Wildin, R.S.; Cuellar-Rodriguez, J.; Hsu, A.P.; Dyack, S.; Fernandez, C.V.; Chong, C.-E.; et al. Loss-of-function germline GATA2 mutations in patients with MDS/AML or monomac syndrome and primary lymphedema reveal a key role for GATA2 in the lymphatic vasculature. Blood 2012, 119, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Fritz-Six, K.L.; Dunworth, W.P.; Li, M.; Caron, K.M. Adrenomedullin signaling is necessary for murine lymphatic vascular development. J. Clin. Investig. 2008, 118, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Tompkins, K.; Batts, L.E.; Puri, M.; Baldwin, S. Abnormal embryonic lymphatic vessel development in TIE1 hypomorphic mice. Development 2010, 137, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Abtahian, F.; Guerriero, A.; Sebzda, E.; Lu, M.M.; Zhou, R.; Mocsai, A.; Myers, E.E.; Huang, B.; Jackson, D.G.; Ferrari, V.A.; et al. Regulation of blood and lymphatic vascular separation by signaling proteins SLP-76 and SYK. Science 2003, 299, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.S.; Dillard, M.E.; Lagutin, O.V.; Lin, F.-J.; Tsai, S.; Tsai, M.-J.; Samokhvalov, I.M.; Oliver, G. Lineage tracing demonstrates the venous origin of the mammalian lymphatic vasculature. Genes Dev. 2007, 21, 2422–2432. [Google Scholar] [CrossRef] [PubMed]
- Carramolino, L.; Fuentes, J.; Garcia-Andres, C.; Azcoitia, V.; Riethmacher, D.; Torres, M. Platelets play an essential role in separating the blood and lymphatic vasculatures during embryonic angiogenesis. Circ. Res. 2010, 106, 1197–1201. [Google Scholar] [CrossRef] [PubMed]
- Uhrin, P.; Zaujec, J.; Breuss, J.M.; Olcaydu, D.; Chrenek, P.; Stockinger, H.; Fuertbauer, E.; Moser, M.; Haiko, P.; Fassler, R.; et al. Novel function for blood platelets and podoplanin in developmental separation of blood and lymphatic circulation. Blood 2010, 115, 3997–4005. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Inoue, K.; Inoue, O.; Ding, G.; Nishimura, S.; Hokamura, K.; Eto, K.; Kashiwagi, H.; Tomiyama, Y.; Yatomi, Y.; Umemura, K.; et al. Essential in vivo roles of the c-type lectin receptor clec-2: Embryonic/neonatal lethality of CLEC-2-deficient mice by blood/lymphatic misconnections and impaired thrombus formation of CLEC-2-deficient platelets. J. Biol. Chem. 2010, 285, 24494–24507. [Google Scholar] [CrossRef] [PubMed]
- Mirza, M.; Pang, M.-F.; Zaini, M.A.; Haiko, P.; Tammela, T.; Alitalo, K.; Philipson, L.; Fuxe, J.; Sollerbrant, K. Essential role of the coxsackie- and adenovirus receptor (CAR) in development of the lymphatic system in mice. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Kizhatil, K.; Ryan, M.; Marchant, J.K.; Henrich, S.; John, S.W.M. Schlemm’s canal is a unique vessel with a combination of blood vascular and lymphatic phenotypes that forms by a novel developmental process. PLoS Biol. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Truong, T.N.; Li, H.; Hong, Y.-K.; Chen, L. Novel characterization and live imaging of Schlemm’s canal expressing PROX-1. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Karpinich, N.O.; Caron, K.M. Schlemm’s canal: More than meets the eye, lymphatics in disguise. J. Clin. Investig. 2014, 124, 3701–3703. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.M. Further studies on facility of flow through the trabecular meshwork. AMA Arch. Ophthalmol. 1958, 60, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.F.; Hoying, J.B.; Witte, M.H.; Daniel Stamer, W. Schlemm’s canal endothelia, lymphatic, or blood vasculature? J. Glaucoma 2007, 16, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Allingham, R.R.; de Kater, A.W.; Ethier, C.R. Schlemm’s canal and primary open angle glaucoma: Correlation between Schlemm’s canal dimensions and outflow facility. Exp. Eye Res. 1996, 62, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Kagemann, L.; Wang, B.; Wollstein, G.; Ishikawa, H.; Nevins, J.E.; Nadler, Z.; Sigal, I.A.; Bilonick, R.A.; Schuman, J.S. IOP elevation reduces Schlemm’s canal cross-sectional area. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1805–1809. [Google Scholar] [CrossRef] [PubMed]
- Kagemann, L.; Wollstein, G.; Ishikawa, H.; Bilonick, R.A.; Brennen, P.M.; Folio, L.S.; Gabriele, M.L.; Schuman, J.S. Identification and assessment of Schlemm’s canal by spectral-domain optical coherence tomography. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4054–4059. [Google Scholar] [CrossRef] [PubMed]
- Kagemann, L.; Nevins, J.E.; Jan, N.-J.; Wollstein, G.; Ishikawa, H.; Kagemann, J.; Sigal, I.A.; Nadler, Z.; Ling, Y.; Schuman, J.S. Characterisation of Schlemm’s canal cross-sectional area. Br. J. Ophthalmol. 2014, 98, ii10–ii14. [Google Scholar] [CrossRef] [PubMed]
- Kagemann, L.; Wollstein, G.; Ishikawa, H.; Sigal, I.A.; Folio, L.S.; Xu, J.; Gong, H.; Schuman, J.S. 3D visualization of aqueous humor outflow structures in-situ in humans. Exp. Eye Res. 2011, 93, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Kagemann, L.; Wollstein, G.; Ishikawa, H.; Nadler, Z.; Sigal, I.A.; Folio, L.S.; Schuman, J.S. Visualization of the conventional outflow pathway in the living human eye. Ophthalmology 2012, 119, 1563–1568. [Google Scholar] [CrossRef] [PubMed]
- Dvorak-Theobald, G. Schlemm’s canal: Its anastomoses and anatomic relations. Trans. Am. Ophthalmol. Soc. 1934, 32, 574–595. [Google Scholar] [PubMed]
- Rosenquist, R.; Epstein, D.; Melamed, S.; Johnson, M.; Grant, W.M. Outflow resistance of enucleated human eyes at two different perfusion pressures and different extents of trabeculotomy. Curr. Eye Res. 1989, 8, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Grierson, I.; Lee, W.R.; Abraham, S.; Howes, R.C. Associations between the cells of the walls of Schlemm’s canal. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 1978, 208, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Hamanaka, T.; Thornell, L.E.; Bill, A. Cytoskeleton and tissue origin in the anterior cynomolgus monkey eye. Jpn. J. Ophthalmol. 1997, 41, 138–149. [Google Scholar] [CrossRef]
- Ethier, C.R.; Read, A.T.; Chan, D. Biomechanics of Schlemm’s canal endothelial cells: Influence on f-actin architecture. Biophys. J. 2004, 87, 2828–2837. [Google Scholar] [CrossRef] [PubMed]
- Grierson, I.; Lee, W.R. Pressure-induced changes in the ultrastructure of the endothelium lining Schlemm’s canal. Am. J. Ophthalmol. 1975, 80, 863–884. [Google Scholar] [CrossRef]
- Bill, A.; Svedbergh, B. Scanning electron microscopic studies of the trabecular meshwork and the canal of Schlemm—An attempt to localize the main resistance to outflow of aqueous humor in man. Acta Ophthalmol. 1972, 50, 295–320. [Google Scholar] [CrossRef]
- Tripathi, R.C. Ultrastructure of Schlemm’s canal in relation to aqueous outflow. Exp. Eye Res. 1968, 7, 335–341. [Google Scholar] [CrossRef]
- Holmberg, A. The fine structure of the inner wall of Schlemms canal. Arch. Ophthalmol. 1959, 62, 956–958. [Google Scholar] [CrossRef]
- Epstein, D.L.; Rohen, J.W. Morphology of the trabecular meshwork and inner-wall endothelium after cationized ferritin perfusion in the monkey eye. Investig. Ophthalmol. Vis. Sci. 1991, 32, 160–171. [Google Scholar]
- Herrnberger, L.; Ebner, K.; Junglas, B.; Tamm, E.R. The role of plasmalemma vesicle-associated protein (PLVAP) in endothelial cells of Schlemm’s canal and ocular capillaries. Exp. Eye Res. 2012, 105, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Inomata, H.; Bill, A.; Smelser, G.K. Aqueous humor pathways through the trabecular meshwork and into Schlemm’s canal in the cynomolgus monkey (Macaca Irus). An electron microscopic study. Am. J. Ophthalmol. 1972, 73, 760–789. [Google Scholar] [CrossRef]
- Overby, D.R.; Stamer, W.D.; Johnson, M. The changing paradigm of outflow resistance generation: Towards synergistic models of the JCT and inner wall endothelium. Exp. Eye Res. 2009, 88, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Folz, S.J.; Laryea, S.N.; Overby, D.R. Multi-scale analysis of segmental outflow patterns in human trabecular meshwork with changing intraocular pressure. J. Ocul. Pharmacol. Ther. 2014, 30, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Hann, C.R.; Fautsch, M.P. Preferential fluid flow in the human trabecular meshwork near collector channels. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.D.; Roberts, B.C.; Howell, D.N.; Epstein, D.L. Isolation, culture, and characterization of endothelial cells from Schlemm’s canal. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1804–1812. [Google Scholar]
- Perkumas, K.M.; Stamer, W.D. Protein markers and differentiation in culture for Schlemm’s canal endothelial cells. Exp. Eye Res. 2012, 96, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Geiger, B.; Epstein, D.L.; Kaufman, P.L. Cytoskeletal involvement in the regulation of aqueous humor outflow. Investig. Ophthalmol. Vis. Sci. 2000, 41, 619–623. [Google Scholar]
- Hristov, M.; Weber, C. Endothelial progenitor cells: Characterization, pathophysiology, and possible clinical relevance. J. Cell Mol. Med. 2004, 8, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Welti, J.; Loges, S.; Dimmeler, S.; Carmeliet, P. Recent molecular discoveries in angiogenesis and antiangiogenic therapies in cancer. J. Clin. Investig. 2013, 123, 3190–3200. [Google Scholar] [CrossRef] [PubMed]
- Garlanda, C.; Dejana, E. Heterogeneity of endothelial cells. Specific markers. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Gavard, J. Endothelial permeability and VE-cadherin: A wacky comradeship. Cell Adh. Migr. 2014, 8, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Lenting, P.J.; Christophe, O.D.; Denis, C.V. von Willebrand factor biosynthesis, secretion, and clearance: connecting the far ends. Blood 2015, 125, 2019–2028. [Google Scholar] [CrossRef] [PubMed]
- Ikomi, F.; Kawai, Y.; Ohhashi, T. Recent advance in lymph dynamic analysis in lymphatics and lymph nodes. Ann. Vasc. Dis. 2012, 5, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.D.; Braakman, S.T.; Zhou, E.H.; Ethier, C.R.; Fredberg, J.J.; Overby, D.R.; Johnson, M. Biomechanics of Schlemm’s canal endothelium and intraocular pressure reduction. Prog. Retin. Eye Res. 2015, 44, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Miteva, D.O.; Rutkowski, J.M.; Dixon, J.B.; Kilarski, W.; Shields, J.D.; Swartz, M.A. Transmural flow modulates cell and fluid transport functions of lymphatic endothelium. Circ. Res. 2010, 106, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Dautriche, C.N.; Szymanski, D.; Kerr, M.; Torrejon, K.Y.; Bergkvist, M.; Xie, Y.; Danias, J.; Stamer, W.D.; Sharfstein, S.T. A biomimetic Schlemm’s canal inner wall: A model to study outflow physiology, glaucoma pathology and high-throughput drug screening. Biomaterials 2015, 65, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Raviola, G.; Raviola, E. Paracellular route of aqueous outflow in the trabecular meshwork and canal of Schlemm. A freeze-fracture study of the endothelial junctions in the sclerocorneal angle of the macaque monkey eye. Investig. Ophthalmol. Vis. Sci. 1981, 21, 52–72. [Google Scholar]
- Swartz, M.A. The physiology of the lymphatic system. Adv. Drug Deliv. Rev. 2001, 50, 3–20. [Google Scholar] [CrossRef]
- Overby, D.R.; Zhou, E.H.; Vargas-Pinto, R.; Pedrigi, R.M.; Fuchshofer, R.; Braakman, S.T.; Gupta, R.; Perkumas, K.M.; Sherwood, J.M.; Vahabikashi, A.; et al. Altered mechanobiology of Schlemm’s canal endothelial cells in glaucoma. Proc. Natl. Acad. Sci. USA 2014, 111, 13876–13881. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Pinto, R.; Lai, J.; Gong, H.; Ethier, C.R.; Johnson, M. Finite element analysis of the pressure-induced deformation of Schlemm’s canal endothelial cells. Biomech. Model. Mechanobiol. 2014, 14, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Zhou, E.H.; Krishnan, R.; Stamer, W.D.; Perkumas, K.M.; Rajendran, K.; Nabhan, J.F.; Lu, Q.; Fredberg, J.J.; Johnson, M. Mechanical responsiveness of the endothelial cell of Schlemm’s canal: Scope, variability and its potential role in controlling aqueous humour outflow. J. R. Soc. Interf. 2012, 9, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Juzkiw, T.; Read, A.T.; Chan, D.W.H.; Glucksberg, M.R.; Ethier, C.R.; Johnson, M. Young’s modulus of elasticity of Schlemm’s canal endothelial cells. Biomech. Model. Mechanobiol. 2010, 9, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Shapiro, A.; Ethier, C.R.; Kamm, R.D. Modulation of outflow resistance by the pores of the inner wall endothelium. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1670–1675. [Google Scholar]
- Johnson, M.; Johnson, D.H.; Kamm, R.D.; DeKater, A.W.; Epstein, D.L. The filtration characteristics of the aqueous outflow system. Exp. Eye Res. 1990, 50, 407–418. [Google Scholar] [CrossRef]
- Allingham, R.R.; de Kater, A.W.; Ethier, C.R.; Anderson, P.J.; Hertzmark, E.; Epstein, D.L. The relationship between pore density and outflow facility in human eyes. Investig. Ophthalmol. Vis. Sci. 1992, 33, 1661–1669. [Google Scholar]
- Alvarado, J.A.; Yeh, R.-F.; Franse-Carman, L.; Marcellino, G.; Brownstein, M.J. Interactions between endothelia of the trabecular meshwork and of Schlemm’s canal: A new insight into the regulation of aqueous outflow in the eye. Trans. Am. Ophthalmol. Soc. 2005, 103, 148–163. [Google Scholar] [PubMed]
- Curcio, C.A. Declining availability of human eye tissues for research. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2747–2749. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Overby, D.R.; Read, A.T.; Stamer, W.D.; Ethier, C.R. A new method for selection of angular aqueous plexus cells from porcine eyes: A model for Schlemm’s canal endothelium. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5744–5750. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D.Z.; Sharif, N.A.; Dismuke, W.M. Endogenous regulation of human Schlemm’s canal cell volume by nitric oxide signaling. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5817–5824. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.-I.; Goldstein, R.A.; Nierras, C.R. The promise of human induced pluripotent stem cells for research and therapy. Nat. Rev. Mol. Cell Biol. 2008, 9, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.L.; Wagers, A.J. No place like home: Anatomy and function of the stem cell niche. Nat. Rev. Mol. Cell Biol. 2008, 9, 11–21. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, B.D.; Ma’ayan, A.; Lemischka, I.R. Systems biology of stem cell fate and cellular reprogramming. Nat. Rev. Mol. Cell Biol. 2009, 10, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Tay, C.Y.; Sathiyanathan, P.; Chu, S.W.; Stanton, L.W.; Wong, T.T. Identification and characterization of mesenchymal stem cells derived from the trabecular meshwork of the human eye. Stem Cells Dev. 2012, 21, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Roh, D.S.; Mann, M.M.; Funderburgh, M.L.; Funderburgh, J.L.; Schuman, J.S. Multipotent stem cells from trabecular meshwork become phagocytic tm cells. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.J.; Zhu, W.; Cook, A.C.; Anfinson, K.R.; Tucker, B.A.; Kuehn, M.H. Induction of trabecular meshwork cells from induced pluripotent stem cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7065–7072. [Google Scholar] [CrossRef]
- Du, Y.; Yun, H.; Yang, E.; Schuman, J.S. Stem cells from trabecular meshwork home to TM tissue in vivo. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in life, disease and medicine. Nature 2005, 438, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Kume, T. Specification of arterial, venous, and lymphatic endothelial cells during embryonic development. Histol. Histopathol. 2010, 25, 637–646. [Google Scholar] [PubMed]
- Swift, M.R.; Weinstein, B.M. Arterial-venous specification during development. Circ. Res. 2009, 104, 576–588. [Google Scholar] [CrossRef] [PubMed]
- You, L.R.; Lin, F.J.; Lee, C.T.; DeMayo, F.J.; Tsai, M.J.; Tsai, S.Y. Suppression of notch signalling by the COUP-TFII transcription factor regulates vein identity. Nature 2005, 435, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.E.; Acott, T.S. The juxtacanalicular region of ocular trabecular meshwork: A tissue with a unique extracellular matrix and specialized function. J. Ocul. Biol. 2013, 1, 10:1–10:7. [Google Scholar]
- Adachi, T.; Osako, Y.; Tanaka, M.; Hojo, M.; Hollister, S.J. Framework for optimal design of porous scaffold microstructure by computational simulation of bone regeneration. Biomaterials 2006, 27, 3964–3972. [Google Scholar] [CrossRef] [PubMed]
- Hollister, S.J. Porous scaffold design for tissue engineering. Nat. Mater. 2005, 4, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Hutmacher, D.W.; Sittinger, M.; Risbud, M.V. Scaffold-based tissue engineering: Rationale for computer-aided design and solid free-form fabrication systems. Trends Biotechnol. 2004, 22, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Ravi, S.; Chaikof, E.L. Biomaterials for vascular tissue engineering. Regen. Med. 2010, 5, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Ravi, S.; Qu, Z.; Chaikof, E.L. Polymeric materials for tissue engineering of arterial substitutes. Vascular 2009, 17, S45–S54. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Memic, A.; Annabi, N.; Hossain, M.; Paul, A.; Dokmeci, M.R.; Dehghani, F.; Khademhosseini, A. Electrospun scaffolds for tissue engineering of vascular grafts. Acta Biomater. 2014, 10, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Woods, I.; Flanagan, T.C. Electrospinning of biomimetic scaffolds for tissue-engineered vascular grafts: Threading the path. Expert Rev. Cardiovasc. Ther. 2014, 12, 815–832. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.-Y.; Lau, K.-T.; Lu, T.-P.; Hui, D. A critical review on polymer-based bio-engineered materials for scaffold development. Compos. B Eng. 2007, 38, 291–300. [Google Scholar] [CrossRef]
- Rhee, D.J.; Haddadin, R.I.; Kang, M.H.; Oh, D.J. Matricellular proteins in the trabecular meshwork. Exp. Eye Res. 2009, 88, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Genasetti, A.; Vigetti, D.; Viola, M.; Karousou, E.; Moretto, P.; Rizzi, M.; Bartolini, B.; Clerici, M.; Pallotti, F.; De Luca, G.; et al. Hyaluronan and human endothelial cell behavior. Connect. Tissue Res. 2008, 49, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.J.; Kielty, C.M.; Walker, M.G.; Canfield, A.E. A novel hyaluronan-based biomaterial (Hyaff-11®) as a scaffold for endothelial cells in tissue engineered vascular grafts. Biomaterials 2004, 25, 5955–5964. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.; Ramamurthi, A. Hyaluronic acid cues for functional endothelialization of vascular constructs. J. Tissue Eng. Regen. Med. 2008, 2, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Boccafoschi, F.; Habermehl, J.; Vesentini, S.; Mantovani, D. Biological performances of collagen-based scaffolds for vascular tissue engineering. Biomaterials 2005, 26, 7410–7417. [Google Scholar] [CrossRef] [PubMed]
- Boland, E.D.; Matthews, J.A.; Pawlowski, K.J.; Simpson, D.G.; Wnek, G.E.; Bowlin, G.L. Electrospinning collagen and elastin: Preliminary vascular tissue engineering. Front. Biosci. 2004, 9, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Berglund, J.D.; Nerem, R.M.; Sambanis, A. Incorporation of intact elastin scaffolds in tissue-engineered collagen-based vascular grafts. Tissue Eng. 2004, 10, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.; Boue, S.; Izpisua Belmonte, J.C. Dedifferentiation, transdifferentiation and reprogramming: Three routes to regeneration. Nat. Rev. Mol. Cell Biol. 2011, 12, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Hannum, C.; Culpepper, J.; Campbell, D.; McClanahan, T.; Zurawski, S.; Bazan, J.F.; Kastelein, R.; Hudak, S.; Wagner, J.; Mattson, J.; et al. Ligand for FLT3/FLK2 receptor tyrosine kinase regulates growth of haematopoietic stem cells and is encoded by variant rnas. Nature 1994, 368, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Leslie, P.; Zhang, Y.; Dong, J. Stem cells in a three-dimensional scaffold environment. Springer Plus 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Weiner, H.; Schwarzbauer, J.E.; Engler, A.J. Defined extracellular matrix components are necessary for definitive endoderm induction. Stem Cells 2013, 31, 2084–2094. [Google Scholar] [CrossRef] [PubMed]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Liersch, R.; Nay, F.; Lu, L.; Detmar, M. Induction of lymphatic endothelial cell differentiation in embryoid bodies. Blood 2006, 107, 1214–1216. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.P.; Acott, T.S. Involvement of the ERK-MAP kinase pathway in TNFalpha regulation of trabecular matrix metalloproteinases and TIMPS. Investig. Ophthalmol. Vis. Sci. 2003, 44, 164–169. [Google Scholar] [CrossRef]
- Bradley, J.M.; Anderssohn, A.M.; Colvis, C.M.; Parshley, D.E.; Zhu, X.H.; Ruddat, M.S.; Samples, J.R.; Acott, T.S. Mediation of laser trabeculoplasty-induced matrix metalloproteinase expression by IL-1beta and TNFalpha. Investig. Ophthalmol. Vis. Sci. 2000, 41, 422–430. [Google Scholar]
- Chang, J.Y.; Stamer, W.D.; Bertrand, J.; Read, A.T.; Marando, C.M.; Ethier, C.R.; Overby, D.R. The role of nitric oxide in murine conventional outflow physiology. Am. J. Physiol. Cell Physiol. 2015, 309, C205–C214. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.F.; Fleissner, F.; Sun, J.; Cooke, J.P. Role of nitric oxide signaling in endothelial differentiation of embryonic stem cells. Stem Cells Dev. 2010, 19, 1617–1625. [Google Scholar] [CrossRef] [PubMed]
- Bishop, C.; Kim, J.; Green, J. Biomolecule delivery to engineer the cellular microenvironment for regenerative medicine. Ann. Biomed. Eng. 2014, 42, 1557–1572. [Google Scholar] [CrossRef] [PubMed]
- Quake, S.R.; Scherer, A. From micro- to nanofabrication with soft materials. Science 2000, 290, 1536–1540. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension: How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Shin, H. Matrices and scaffolds for delivery of bioactive molecules in bone and cartilage tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 339–359. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Park, T.G. Surface engineered and drug releasing pre-fabricated scaffolds for tissue engineering. Adv. Drug Deliv. Rev. 2007, 59, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Pedrigi, R.M.; Simon, D.; Reed, A.; Stamer, W.D.; Overby, D.R. A model of giant vacuole dynamics in human Schlemm’s canal endothelial cells. Exp. Eye Res. 2011, 92, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.G.; Zhou, W.; O’Brien, E.T.; Roberts, B.C.; Stamer, W.D. Effect of hydrostatic pressure gradients and Na2EDTA on permeability of human Schlemm’s canal cell monolayers. Curr. Eye Res. 2004, 28, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.D.; Roberts, B.C.; Epstein, D.L. Hydraulic pressure stimulates adenosine 3′,5′-cyclic monophosphate accumulation in endothelial cells from Schlemm’s canal. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1983–1988. [Google Scholar]
- Schindler, M.; Nur, E.K.A.; Ahmed, I.; Kamal, J.; Liu, H.Y.; Amor, N.; Ponery, A.S.; Crockett, D.P.; Grafe, T.H.; Chung, H.Y.; et al. Living in three dimensions: 3D nanostructured environments for cell culture and regenerative medicine. Cell Biochem. Biophys. 2006, 45, 215–227. [Google Scholar] [CrossRef]
- Albelda, S.M.; Sampson, P.M.; Haselton, F.R.; McNiff, J.M.; Mueller, S.N.; Williams, S.K.; Fishman, A.P.; Levine, E.M. Permeability characteristics of cultured endothelial cell monolayers. J. Appl. Physiol. 1988, 64, 308–322. [Google Scholar] [PubMed]
- Santaguida, S.; Janigro, D.; Hossain, M.; Oby, E.; Rapp, E.; Cucullo, L. Side by side comparison between dynamic versus static models of blood–brain barrier in vitro: A permeability study. Brain Res. 2006, 1109, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Torrejon, K.Y.; Pu, D.; Bergkvist, M.; Danias, J.; Sharfstein, S.T.; Xie, Y. Recreating a human trabecular meshwork outflow system on microfabricated porous structures. Biotechnol. Bioeng. 2013, 110, 3205–3218. [Google Scholar] [CrossRef] [PubMed]
- Dautriche, C.N.; Xie, Y.; Sharfstein, S.T. Walking through trabecular meshwork biology: Toward engineering design of outflow physiology. Biotechnol. Adv. 2014, 32, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Helm, C.L.; Zisch, A.; Swartz, M.A. Engineered blood and lymphatic capillaries in 3-D VEGF-fibrin-collagen matrices with interstitial flow. Biotechnol. Bioeng. 2007, 96, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.P.; Helm, C.L.; Swartz, M.A. Interstitial flow differentially stimulates blood and lymphatic endothelial cell morphogenesis in vitro. Microvasc. Res. 2004, 68, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Boardman, K.C.; Swartz, M.A. Interstitial flow as a guide for lymphangiogenesis. Circ. Res. 2003, 92, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Niebruegge, S.; Bauwens, C.L.; Peerani, R.; Thavandiran, N.; Masse, S.; Sevaptisidis, E.; Nanthakumar, K.; Woodhouse, K.; Husain, M.; Kumacheva, E.; et al. Generation of human embryonic stem cell-derived mesoderm and cardiac cells using size-specified aggregates in an oxygen-controlled bioreactor. Biotechnol. Bioeng. 2009, 102, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Sargent, C.Y.; Berguig, G.Y.; McDevitt, T.C. Cardiomyogenic differentiation of embryoid bodies is promoted by rotary orbital suspension culture. Tissue Eng. A 2009, 15, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Parikh, A.; Tzanakakis, E.S. Cardiac cell generation from encapsulated embryonic stem cells in static and scalable culture systems. Cell Transplant. 2010, 19, 1397–1412. [Google Scholar] [CrossRef] [PubMed]
- Illi, B.; Scopece, A.; Nanni, S.; Farsetti, A.; Morgante, L.; Biglioli, P.; Capogrossi, M.C.; Gaetano, C. Epigenetic histone modification and cardiovascular lineage programming in mouse embryonic stem cells exposed to laminar shear stress. Circ. Res. 2005, 96, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Hazeltine, L.B.; Selekman, J.A.; Palecek, S.P. Engineering the human pluripotent stem cell microenvironment to direct cell fate. Biotechnol. Adv. 2013, 31, 1002–1091. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dautriche, C.N.; Tian, Y.; Xie, Y.; Sharfstein, S.T. A Closer Look at Schlemm’s Canal Cell Physiology: Implications for Biomimetics. J. Funct. Biomater. 2015, 6, 963-985. https://doi.org/10.3390/jfb6030963
Dautriche CN, Tian Y, Xie Y, Sharfstein ST. A Closer Look at Schlemm’s Canal Cell Physiology: Implications for Biomimetics. Journal of Functional Biomaterials. 2015; 6(3):963-985. https://doi.org/10.3390/jfb6030963
Chicago/Turabian StyleDautriche, Cula N., Yangzi Tian, Yubing Xie, and Susan T. Sharfstein. 2015. "A Closer Look at Schlemm’s Canal Cell Physiology: Implications for Biomimetics" Journal of Functional Biomaterials 6, no. 3: 963-985. https://doi.org/10.3390/jfb6030963