Fluorescent PSC-Derived Cardiomyocyte Reporter Lines: Generation Approaches and Their Applications in Cardiovascular Medicine


Abstract
:Simple Summary
Abstract
1. Introduction
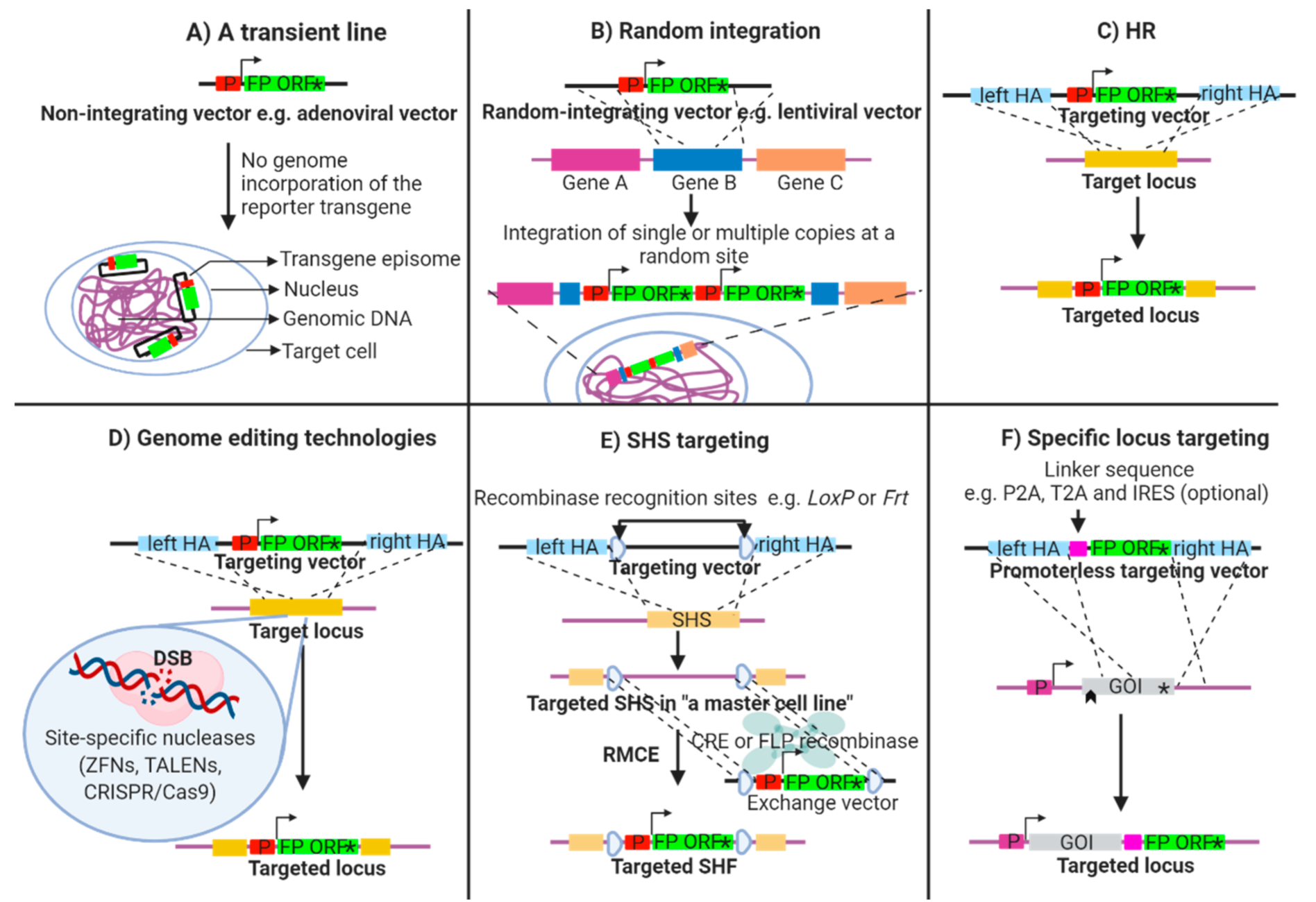
2. Establishment of Transgenic Fluorescent Reporter Lines
2.1. Transient Fluorescent Reporter Lines
2.2. Random Integration of a Fluorescent Reporter Transgene
2.3. Targeted Integration of a Fluorescent Reporter Transgene
2.4. Advanced Genome Engineering Technologies
3. PSC-CM Reporter Lines: An Insight into Cardiac Biology
4. Fluorescence-Guided PSC-CM Purification
4.1. Functional CMs
4.2. Ventricular CM
4.3. Atrial CM
4.4. Nodal CM
5. Applications towards Optical Measurements of Cardiac Functions
5.1. Genetically Encoded Fluorescent Voltage-Sensitive and Ca2+ Indicators
5.2. Optogenetics
6. Emerging Insights into Cardiac Tissue Repair
7. Maturation Approaches
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | 3-dimentional |
| ACTC | α-cardiac actin |
| AP | action potential |
| BAC | bacterial artificial chromosome |
| BRY | T-box transcription factor brachyury |
| CaM | Ca2+-modulating protein |
| cGATA6 | chick GATA6 |
| ChR2 | channelrhodopsin 2 |
| CMs | cardiomyocytes |
| CPCs | cardiac progenitor cells |
| CRISPR/Cas9 | clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR associated protein 9 (Cas9) |
| CT | Ca2+ transient |
| CX43 | connexin 43 |
| DSB | double strand break |
| ESCs | embryonic stem cells |
| FACS | fluorescence-activated cell sorting |
| FHF | first heart field |
| FP | fluorescent protein |
| GECIs | genetically encoded Ca2+ indicator |
| GEVIs | genetically encoded voltage-sensitive indicators |
| GOI | gene of interest |
| HCN4 | hyperpolisation-activated cyclic nucleotide gated K+ channel 4 |
| hESC-CMs | human embryonic stem cell-derived cardiomyocytes |
| hESCs | human embryonic stem cells |
| hiPSC-CMs | human induced pluripotent stem cell-derived cardiomyocytes |
| hiPSCs | human induced pluripotent stem cells |
| hPSC-CMs | human pluripotent stem cell-derived cardiomyocytes |
| HR | homologous recombination |
| iPSCs | induced pluripotent stem cells |
| IRES | internal ribosomal entry sites |
| ISL1 | insulin gene enhancer protein |
| Kdr | kinase insert domain receptor |
| mESCs | mouse embryonic stem cells |
| MESP1 | mesoderm posterior basic helix-loop-helix transcription factor 1 |
| miPSC-CMs | mouse induced pluripotent stem cell-derived cardiomyocytes |
| miPSCs | mouse induced pluripotent stem cells |
| MYH6 | α-myosin heavy chain |
| MYL2 | myosin light chain 2 |
| MYL7 | myosin light chain 7 |
| NCX1 | Na+/Ca2+ antiporter |
| NKX2.5 | NK2 homeobox 5 |
| P2A | porcine teschovirus-1 2A |
| PSCs | pluripotent stem cells |
| RMCE | recombination-mediated cassette exchange |
| SHF | second heart field |
| SHOX2 | short stature homeobox 2 |
| SHS | safe harbour site |
| SLN | sarcolipin |
| T2A | Thosea asigna virus 2A |
| TALENs | transcription activator-like effector nucleases |
| TBX5 | T-box transcription factor 5 |
| TNNT | cardiac troponin |
| TTN | titin |
| VSFP | voltage sensitive fluorescent protein |
| ZFNs | zinc finger nucleases |
References
- Penney, J.; Ralvenius, W.T.; Tsai, L.H. Modeling Alzheimer’s disease with iPSC-derived brain cells. Mol. Psychiatry 2020, 25, 148–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, J.L.; Duncan, S.A. iPSC-derived hepatocytes as a platform for disease modeling and drug discovery. Front. Med. 2019, 6, 265. [Google Scholar] [CrossRef] [PubMed]
- Prajumwongs, P.; Weeranantanapan, O.; Jaroonwitchawan, T.; Noisa, P. Human Embryonic Stem Cells: A Model for the Study of Neural Development and Neurological Diseases. Stem Cells Int. 2016, 2016, 2958210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Oikonomopoulos, A.; Sayed, N.; Wu, J.C. Modeling human diseases with induced pluripotent stem cells: From 2D to 3D and beyond. Development 2018, 145, dev156166. [Google Scholar] [CrossRef] [Green Version]
- Passier, R.; Orlova, V.; Mummery, C. Complex tissue and disease modeling using hiPSCs. Cell Stem Cell 2016, 18, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Le Bras, A. A resource for selecting animal models of heart disease. Lab Anim. 2019, 48, 332. [Google Scholar] [CrossRef]
- Camacho, P.; Fan, H.; Liu, Z.; He, J.Q. Large Mammalian Animal Models of Heart Disease. J. Cardiovasc. Dev. Dis. 2016, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Perlman, R.L. Mouse models of human diseaseAn evolutionary perspective. Evol. Med. Public Health 2016, 2016, 170–176. [Google Scholar]
- Barre-Sinoussi, F.; Montagutelli, X. Animal models are essential to biological research: Issues and perspectives. Future Sci. OA 2015, 1. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.A. Human embryonic stem cell research: Ethical and legal issues. Nat. Rev. Genet. 2001, 2, 74–78. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnuki, M.; Takahashi, K. Present and future challenges of induced pluripotent stem cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140367. [Google Scholar] [CrossRef] [PubMed]
- Bilic, J.; Belmonte, J.C.I. Concise review: Induced pluripotent stem cells versus embryonic stem cells: Close enough or yet too far apart? Stem Cells 2012, 30, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Bilousova, G.; Roop, D.R. Induced pluripotent stem cells in dermatology: Potentials, advances, and limitations. Cold Spring Harb. Perspect. Med. 2014, 4, a015164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, M.; Hayashizaki, Y.; Murakawa, Y. Genomic instability of iPSCs: Challenges towards their clinical applications. Stem Cell Rev. Rep. 2017, 13, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.; Nguyen, W.; Nguyenton, B.; Ratchada, P.; Page, G.; Miller, P.E.; Ghetti, A.; Abi-Gerges, N. Adult Human Primary Cardiomyocyte-Based Model for the Simultaneous Prediction of Drug-Induced Inotropic and Pro-arrhythmia Risk. Front. Physiol. 2017, 8, 1073. [Google Scholar] [CrossRef] [Green Version]
- Abi-Gerges, N.; Indersmitten, T.; Truong, K.; Nguyen, W.; Ratchada, P.; Nguyen, N.; Page, G.; Miller, P.E.; Ghetti, A. Multiparametric Mechanistic Profiling of Inotropic Drugs in Adult Human Primary Cardiomyocytes. Sci. Rep. 2020, 10, 7692. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Guo, H.; Wu, J.C. Applications of genetically engineered human pluripotent stem cell reporters in cardiac stem cell biology. Curr. Opin. Biotechnol. 2018, 52, 66–73. [Google Scholar] [CrossRef]
- Nayerossadat, N.; Maedeh, T.; Ali, P.A. Viral and nonviral delivery systems for gene delivery. Adv. Biomed. Res. 2012, 1, 27. [Google Scholar] [CrossRef]
- Den Hartogh, S.C.; Passier, R.J.S.C. Concise review: Fluorescent reporters in human pluripotent stem cells: Contributions to cardiac differentiation and their applications in cardiac disease and toxicity. Stem Cells 2016, 34, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.R.; Maguire, S.; Davis, L.A.; Alexander, M.; Yang, F.; Chandran, S.; Ffrench-Constant, C.; Pedersen, R.A. Robust, persistent transgene expression in human embryonic stem cells is achieved with AAVS1-targeted integration. Stem Cells 2008, 26, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.; Bellen, H.J.; Gehring, W.J. Position effects on eukaryotic gene expression. Annu. Rev. Cell Biol. 1990, 6, 679–714. [Google Scholar] [CrossRef]
- Klatt, D.; Cheng, E.; Hoffmann, D.; Santilli, G.; Thrasher, A.J.; Brendel, C.; Schambach, A. Differential Transgene Silencing of Myeloid-Specific Promoters in the AAVS1 Safe Harbor Locus of Induced Pluripotent Stem Cell-Derived Myeloid Cells. Hum. Gene Ther. 2020, 31, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Calero-Nieto, F.J.; Bert, A.G.; Cockerill, P.N. Transcription-dependent silencing of inducible convergent transgenes in transgenic mice. Epigenetics Chromatin 2010, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenqvist, N.; Jakobsson, J.; Lundberg, C. Inhibition of chromatin condensation prevents transgene silencing in a neural progenitor cell line transplanted to the rat brain. Cell Transpl. 2005, 14, 129–138. [Google Scholar] [CrossRef]
- Sung, P.; Klein, H. Mechanism of homologous recombination: Mediators and helicases take on regulatory functions. Nat. Rev. Mol. Cell Biol. 2006, 7, 739–750. [Google Scholar] [CrossRef]
- Leavitt, A.D.; Hamlett, I. Homologous recombination in human embryonic stem cells: A tool for advancing cell therapy and understanding and treating human disease. Clin. Transl. Sci. 2011, 4, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef]
- Rouet, P.; Smih, F.; Jasin, M. Introduction of double-strand breaks into the genome of mouse cells by expression of a rare-cutting endonuclease. Mol. Cell. Biol. 1994, 14, 8096–8106. [Google Scholar] [CrossRef] [Green Version]
- Oceguera-Yanez, F.; Kim, S.-I.; Matsumoto, T.; Tan, G.W.; Xiang, L.; Hatani, T.; Kondo, T.; Ikeya, M.; Yoshida, Y.; Inoue, H. Engineering the AAVS1 locus for consistent and scalable transgene expression in human iPSCs and their differentiated derivatives. Methods 2016, 101, 43–55. [Google Scholar] [CrossRef]
- Irion, S.; Luche, H.; Gadue, P.; Fehling, H.J.; Kennedy, M.; Keller, G. Identification and targeting of the ROSA26 locus in human embryonic stem cells. Nat. Biotechnol. 2007, 25, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Papapetrou, E.P.; Schambach, A.J.M.T. Gene insertion into genomic safe harbors for human gene therapy. Mol. Ther. 2016, 24, 678–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turan, S.; Galla, M.; Ernst, E.; Qiao, J.; Voelkel, C.; Schiedlmeier, B.; Zehe, C.; Bode, J. Recombinase-mediated cassette exchange (RMCE): Traditional concepts and current challenges. J. Mol. Biol. 2011, 407, 193–221. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Sierra, G.; Sivapatham, R.; Swistowski, A.; Rao, M.S.; Zeng, X. A platform for rapid generation of single and multiplexed reporters in human iPSC lines. Sci. Rep. 2015, 5, 9205. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Chen, O.; Wall, J.B.J.; Zheng, M.; Zhou, Y.; Wang, L.; Vaseghi, H.R.; Qian, L.; Liu, J. Systematic comparison of 2A peptides for cloning multi-genes in a polycistronic vector. Sci. Rep. 2017, 7, 2193. [Google Scholar] [CrossRef]
- Komar, A.A.; Hatzoglou, M. Cellular IRES-mediated translation: The war of ITAFs in pathophysiological states. Cell Cycle 2011, 10, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Szymczak, A.L.; Vignali, D.A. Development of 2A peptide-based strategies in the design of multicistronic vectors. Expert Opin. Biol. Ther. 2005, 5, 627–638. [Google Scholar] [CrossRef]
- Naito, A.T.; Shiojima, I.; Akazawa, H.; Hidaka, K.; Morisaki, T.; Kikuchi, A.; Komuro, I. Developmental stage-specific biphasic roles of Wnt/beta-catenin signaling in cardiomyogenesis and hematopoiesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19812–19817. [Google Scholar] [CrossRef] [Green Version]
- Kouskoff, V.; Lacaud, G.; Schwantz, S.; Fehling, H.J.; Keller, G. Sequential development of hematopoietic and cardiac mesoderm during embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2005, 102, 13170–13175. [Google Scholar] [CrossRef] [Green Version]
- Fehling, H.J.; Lacaud, G.; Kubo, A.; Kennedy, M.; Robertson, S.; Keller, G.; Kouskoff, V. Tracking mesoderm induction and its specification to the hemangioblast during embryonic stem cell differentiation. Development 2003, 130, 4217–4227. [Google Scholar] [CrossRef] [Green Version]
- Den Hartogh, S.C.; Schreurs, C.; Monshouwer-Kloots, J.J.; Davis, R.P.; Elliott, D.A.; Mummery, C.L.; Passier, R. Dual reporter mesp1mcherry/w-nkx2-5egfp/w hescs enable studying early human cardiac differentiation. Stem Cells 2015, 33, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Chiapparo, G.; Lin, X.; Lescroart, F.; Chabab, S.; Paulissen, C.; Pitisci, L.; Bondue, A.; Blanpain, C. Mesp1 controls the speed, polarity, and directionality of cardiovascular progenitor migration. J. Cell Biol. 2016, 213, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Z.; Termglinchan, V.; Shao, N.Y.; Itzhaki, I.; Liu, C.; Ma, N.; Tian, L.; Wang, V.Y.; Chang, A.C.Y.; Guo, H.; et al. A Human iPSC Double-Reporter System Enables Purification of Cardiac Lineage Subpopulations with Distinct Function and Drug Response Profiles. Cell Stem Cell 2019, 24, 802–811.e5. [Google Scholar] [CrossRef] [PubMed]
- Meilhac, S.M.; Buckingham, M.E. The deployment of cell lineages that form the mammalian heart. Nat. Rev. Cardiol. 2018, 15, 705–724. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.; Jiang, X.; Martin-Puig, S.; Caron, L.; Zhu, S.; Shao, Y.; Roberts, D.J.; Huang, P.L.; Domian, I.J.; Chien, K.R. Human ISL1 heart progenitors generate diverse multipotent cardiovascular cell lineages. Nature 2009, 460, 113–117. [Google Scholar] [CrossRef]
- El-Nachef, D.; Shi, K.; Beussman, K.M.; Martinez, R.; Regier, M.C.; Everett, G.W.; Murry, C.E.; Stevens, K.R.; Young, J.E.; Sniadecki, N.J. A Rainbow Reporter Tracks Single Cells and Reveals Heterogeneous Cellular Dynamics among Pluripotent Stem Cells and Their Differentiated Derivatives. Stem Cell Rep. 2020, 15, 226–241. [Google Scholar] [CrossRef]
- Elliott, D.A.; Braam, S.R.; Koutsis, K.; Ng, E.S.; Jenny, R.; Lagerqvist, E.L.; Biben, C.; Hatzistavrou, T.; Hirst, C.E.; Yu, Q.C.; et al. NKX2-5(eGFP/w) hESCs for isolation of human cardiac progenitors and cardiomyocytes. Nat. Methods 2011, 8, 1037–1040. [Google Scholar] [CrossRef]
- Moon, S.H.; Ban, K.; Kim, C.; Kim, S.S.; Byun, J.; Song, M.K.; Park, I.H.; Yu, S.P.; Yoon, Y.S. Development of a novel two-dimensional directed differentiation system for generation of cardiomyocytes from human pluripotent stem cells. Int. J. Cardiol. 2013, 168, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Ritner, C.; Wong, S.S.; King, F.W.; Mihardja, S.S.; Liszewski, W.; Erle, D.J.; Lee, R.J.; Bernstein, H.S. An engineered cardiac reporter cell line identifies human embryonic stem cell-derived myocardial precursors. PLoS ONE 2011, 6, e16004. [Google Scholar] [CrossRef] [Green Version]
- Tsai, S.-Y.; Ghazizadeh, Z.; Wang, H.-J.; Amin, S.; Ortega, F.A.; Badieyan, Z.S.; Hsu, Z.-T.; Gordillo, M.; Kumar, R.; Christini, D.J. A human embryonic stem cell reporter line for monitoring chemical-induced cardiotoxicity. Cardiovasc. Res. 2020, 116, 658–670. [Google Scholar] [CrossRef]
- Katarzyna, F.; Natalia, R.; Agnieszka, Z.; Agnieszka, M.; Katarzyna, T.; Izabela, S.; Karolina, N.-B.; Magdalena, N.; Kolanowski, T.J.; Wojciech, Ł. Tissue-specific promoter-based reporter system for monitoring cell differentiation from iPSCs to cardiomyocytes. Sci. Rep. 2020, 10, 1895. [Google Scholar]
- Sharma, A.; Toepfer, C.N.; Schmid, M.; Garfinkel, A.C.; Seidman, C.E. Differentiation and Contractile Analysis of GFP-Sarcomere Reporter hiPSC-Cardiomyocytes. Curr. Protoc. Hum. Genet. 2018, 96, 21.12.1–21.12.12. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, D.A.; Hidalgo, A.; Yang, S.-K.; Zhang, X.; Hudson, J.; Mazzone, S.B.; Chen, C.; Cooper-White, J.J.; Wolvetang, E.J. Isolation of contractile cardiomyocytes from human pluripotent stem-cell-derived cardiomyogenic cultures using a human NCX1-EGFP reporter. Stem Cells Dev. 2015, 24, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.-D.; Jiang, P.; Rushing, S.; Liu, J.; Chiamvimonvat, N.; Li, R.A. Na+/Ca2+ exchanger is a determinant of excitation–contraction coupling in human embryonic stem cell–derived ventricular cardiomyocytes. Stem Cells Dev. 2010, 19, 773–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, I.; Itzhaki, I.; Caspi, O.; Arbel, G.; Tzukerman, M.; Gepstein, A.; Habib, M.; Yankelson, L.; Kehat, I.; Gepstein, L. Identification and selection of cardiomyocytes during human embryonic stem cell differentiation. FASEB J. 2007, 21, 2551–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.; Fleischmann, B.K.; Selbert, S.; Ji, G.J.; Endl, E.; Middeler, G.; Muller, O.J.; Schlenke, P.; Frese, S.; Wobus, A.M.; et al. Selection of ventricular-like cardiomyocytes from ES cells in vitro. FASEB J. 2000, 14, 2540–2548. [Google Scholar] [CrossRef] [Green Version]
- Bizy, A.; Guerrero-Serna, G.; Hu, B.; Ponce-Balbuena, D.; Willis, B.C.; Zarzoso, M.; Ramirez, R.J.; Sener, M.F.; Mundada, L.V.; Klos, M.; et al. Myosin light chain 2-based selection of human iPSC-derived early ventricular cardiac myocytes. Stem Cell Res. 2013, 11, 1335–1347. [Google Scholar] [CrossRef]
- Chen, Z.; Xian, W.; Bellin, M.; Dorn, T.; Tian, Q.; Goedel, A.; Dreizehnter, L.; Schneider, C.M.; Ward-van Oostwaard, D.; Ng, J.K.M.; et al. Subtype-specific promoter-driven action potential imaging for precise disease modelling and drug testing in hiPSC-derived cardiomyocytes. Eur. Heart J. 2016, 38, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Josowitz, R.; Lu, J.; Falce, C.; D’Souza, S.L.; Wu, M.; Cohen, N.; Dubois, N.C.; Zhao, Y.; Sobie, E.A.; Fishman, G.I.; et al. Identification and purification of human induced pluripotent stem cell-derived atrial-like cardiomyocytes based on sarcolipin expression. PLoS ONE 2014, 9, e101316. [Google Scholar] [CrossRef] [Green Version]
- Schwach, V.; Verkerk, A.O.; Mol, M.; Monshouwer-Kloots, J.J.; Devalla, H.D.; Orlova, V.V.; Anastassiadis, K.; Mummery, C.L.; Davis, R.P.; Passier, R. A COUP-TFII Human Embryonic Stem Cell Reporter Line to Identify and Select Atrial Cardiomyocytes. Stem Cell Rep. 2017, 9, 1765–1779. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.Z.; Xie, Y.; Moyes, K.W.; Gold, J.D.; Askari, B.; Laflamme, M.A. Neuregulin/ErbB signaling regulates cardiac subtype specification in differentiating human embryonic stem cells. Circ. Res. 2010, 107, 776–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinnawi, R.; Huber, I.; Maizels, L.; Shaheen, N.; Gepstein, A.; Arbel, G.; Tijsen, A.J.; Gepstein, L. Monitoring Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes with Genetically Encoded Calcium and Voltage Fluorescent Reporters. Stem Cell Rep. 2015, 5, 582–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempsey, G.T.; Chaudhary, K.W.; Atwater, N.; Nguyen, C.; Brown, B.S.; McNeish, J.D.; Cohen, A.E.; Kralj, J.M. Cardiotoxicity screening with simultaneous optogenetic pacing, voltage imaging and calcium imaging. J. Pharmacol. Toxicol. Methods 2016, 81, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Lapp, H.; Bruegmann, T.; Malan, D.; Friedrichs, S.; Kilgus, C.; Heidsieck, A.; Sasse, P. Frequency-dependent drug screening using optogenetic stimulation of human iPSC-derived cardiomyocytes. Sci. Rep. 2017, 7, 9629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Liu, H.; Klein, M.; Ostrominski, J.; Hong, S.G.; Yada, R.C.; Chen, G.; Navarengom, K.; Schwartzbeck, R.; San, H.; et al. Efficient differentiation of cardiomyocytes and generation of calcium-sensor reporter lines from nonhuman primate iPSCs. Sci. Rep. 2018, 8, 5907. [Google Scholar] [CrossRef]
- Liu, Z.; Tang, Y.; Lü, S.; Zhou, J.; Du, Z.; Duan, C.; Li, Z.; Wang, C. The tumourigenicity of iPS cells and their differentiated derivates. J. Cell Mol. Med. 2013, 17, 782–791. [Google Scholar] [CrossRef]
- Mummery, C.L.; Zhang, J.; Ng, E.S.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes: A methods overview. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef]
- Christoforou, N.; Liau, B.; Chakraborty, S.; Chellapan, M.; Bursac, N.; Leong, K.W. Induced pluripotent stem cell-derived cardiac progenitors differentiate to cardiomyocytes and form biosynthetic tissues. PLoS ONE 2013, 8, e65963. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tao, R.; Campbell, K.F.; Carvalho, J.L.; Ruiz, E.C.; Kim, G.C.; Schmuck, E.G.; Raval, A.N.; da Rocha, A.M.; Herron, T.J. Functional cardiac fibroblasts derived from human pluripotent stem cells via second heart field progenitors. Nat. Commun. 2019, 10, 2238. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Police, S.; Rao, N.; Carpenter, M.K. Characterization and enrichment of cardiomyocytes derived from human embryonic stem cells. Circ. Res. 2002, 91, 501–508. [Google Scholar] [CrossRef]
- Sharma, A.; Li, G.; Rajarajan, K.; Hamaguchi, R.; Burridge, P.W.; Wu, S.M. Derivation of highly purified cardiomyocytes from human induced pluripotent stem cells using small molecule-modulated differentiation and subsequent glucose starvation. JoVE 2015, e52628. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Linders, A.; Yamak, A.; Correia, C.; Kijlstra, J.D.; Garakani, A.; Xiao, L.; Milan, D.J.; van der Meer, P.; Serra, M.; et al. Metabolic Maturation of Human Pluripotent Stem Cell-Derived Cardiomyocytes by Inhibition of HIF1alpha and LDHA. Circ. Res. 2018, 123, 1066–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, F.; Chen, H.; Yamashita, H.; Tohyama, S.; Satoh, Y.S.; Yuasa, S.; Li, W.; Yamakawa, H.; Tanaka, T.; Onitsuka, T.; et al. Nongenetic method for purifying stem cell-derived cardiomyocytes. Nat. Methods 2010, 7, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.F.; Danoviz, M.E.; Wiczer, B.; Laflamme, M.A.; Tian, R. Mitochondrial Maturation in Human Pluripotent Stem Cell Derived Cardiomyocytes. Stem Cells Int. 2017, 2017, 5153625. [Google Scholar] [CrossRef]
- Klug, M.G.; Soonpaa, M.H.; Koh, G.Y.; Field, L.J. Genetically selected cardiomyocytes from differentiating embronic stem cells form stable intracardiac grafts. J. Clin. Investig. 1996, 98, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Kita-Matsuo, H.; Barcova, M.; Prigozhina, N.; Salomonis, N.; Wei, K.; Jacot, J.G.; Nelson, B.; Spiering, S.; Haverslag, R.; Kim, C.; et al. Lentiviral vectors and protocols for creation of stable hESC lines for fluorescent tracking and drug resistance selection of cardiomyocytes. PLoS ONE 2009, 4, e5046. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zweigerdt, R.; Xu, X.; Zweigerdt, R.; Soo, S.; Ngoh, Z.; Tham, S.; Wang, S.; Graichen, R.; Davidson, B. Highly enriched cardiomyocytes from human embryonic stem cells. Cytotherapy 2008, 10, 376–389. [Google Scholar] [CrossRef]
- Anderson, D.; Self, T.; Mellor, I.R.; Goh, G.; Hill, S.J.; Denning, C. Transgenic enrichment of cardiomyocytes from human embryonic stem cells. Mol. Ther. 2007, 15, 2027–2036. [Google Scholar] [CrossRef]
- Jung, J.J.; Husse, B.; Rimmbach, C.; Krebs, S.; Stieber, J.; Steinhoff, G.; Dendorfer, A.; Franz, W.-M.; David, R. Programming and isolation of highly pure physiologically and pharmacologically functional sinus-nodal bodies from pluripotent stem cells. Stem Cell Rep. 2014, 2, 592–605. [Google Scholar] [CrossRef] [Green Version]
- Hannes, T.; Wolff, M.; Doss, M.X.; Pfannkuche, K.; Haustein, M.; Müller-Ehmsen, J.; Sachinidis, A.; Hescheler, J.; Khalil, M.; Halbach, M. Electrophysiological characteristics of embryonic stem cell-derived cardiomyocytes are cell line-dependent. Cell. Physiol. Biochem. 2015, 35, 305–314. [Google Scholar] [CrossRef]
- Belus, A.; White, E. Effects of antibiotics on the contractility and Ca2+ transients of rat cardiac myocytes. Eur. J. Pharmacol. 2001, 412, 121–126. [Google Scholar] [CrossRef]
- Anderson, D.J.; Kaplan, D.I.; Bell, K.M.; Koutsis, K.; Haynes, J.M.; Mills, R.J.; Phelan, D.G.; Qian, E.L.; Leitoguinho, A.R.; Arasaratnam, D.; et al. NKX2-5 regulates human cardiomyogenesis via a HEY2 dependent transcriptional network. Nat. Commun. 2018, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, H.; Bartunkova, S.; Schinke, M.; Tanaka, M.; Izumo, S. Cardiac and extracardiac expression of Csx/Nkx2.5 homeodomain protein. Circ. Res. 1998, 82, 936–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaBarge, W.; Mattappally, S.; Kannappan, R.; Fast, V.G.; Pretorius, D.; Berry, J.L.; Zhang, J. Maturation of three-dimensional, hiPSC-derived cardiomyocyte spheroids utilizing cyclic, uniaxial stretch and electrical stimulation. PLoS ONE 2019, 14, e0219442. [Google Scholar]
- Guo, Y.; Pu, W.T. Cardiomyocyte maturation: New phase in development. Circ. Res. 2020, 126, 1086–1106. [Google Scholar] [CrossRef]
- Cui, M.; Wang, Z.; Bassel-Duby, R.; Olson, E.N. Genetic and epigenetic regulation of cardiomyocytes in development, regeneration and disease. Development 2018, 145, dev171983. [Google Scholar] [CrossRef] [Green Version]
- Toepfer, C.N.; Garfinkel, A.C.; Venturini, G.; Wakimoto, H.; Repetti, G.; Alamo, L.; Sharma, A.; Agarwal, R.; Ewoldt, J.F.; Cloonan, P.; et al. Myosin Sequestration Regulates Sarcomere Function, Cardiomyocyte Energetics, and Metabolism, Informing the Pathogenesis of Hypertrophic Cardiomyopathy. Circulation 2020, 141, 828–842. [Google Scholar] [CrossRef]
- Kane, C.; Terracciano, C.M. Concise Review: Criteria for Chamber-Specific Categorization of Human Cardiac Myocytes Derived from Pluripotent Stem Cells. Stem Cells 2017, 35, 1881–1897. [Google Scholar] [CrossRef] [Green Version]
- Paik, D.T.; Chandy, M.; Wu, J.C. Patient and Disease–Specific Induced Pluripotent Stem Cells for Discovery of Personalized Cardiovascular Drugs and Therapeutics. Pharmacol. Rev. 2020, 72, 320–342. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, F.; Lyon, R.C.; Chen, J. Functions of myosin light chain-2 (MYL2) in cardiac muscle and disease. Gene 2015, 569, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Piccini, I.; Rao, J.; Seebohm, G.; Greber, B. Human pluripotent stem cell-derived cardiomyocytes: Genome-wide expression profiling of long-term in vitro maturation in comparison to human heart tissue. Genom. Data 2015, 4, 69–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, D.; Lamers, W.H.; Moorman, A.F. Patterns of expression in the developing myocardium: Towards a morphologically integrated transcriptional model. Cardiovasc. Res. 1998, 38, 25–53. [Google Scholar] [CrossRef] [Green Version]
- Minamisawa, S.; Wang, Y.; Chen, J.; Ishikawa, Y.; Chien, K.R.; Matsuoka, R. Atrial chamber-specific expression of sarcolipin is regulated during development and hypertrophic remodeling. J. Biol. Chem. 2003, 278, 9570–9575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devalla, H.D.; Schwach, V.; Ford, J.W.; Milnes, J.T.; El-Haou, S.; Jackson, C.; Gkatzis, K.; Elliott, D.A.; de Sousa Lopes, S.M.C.; Mummery, C.L.; et al. Atrial-like cardiomyocytes from human pluripotent stem cells are a robust preclinical model for assessing atrial-selective pharmacology. EMBO Mol. Med. 2015, 7, 394–410. [Google Scholar] [CrossRef] [PubMed]
- Yechikov, S.; Copaciu, R.; Gluck, J.M.; Deng, W.; Chiamvimonvat, N.; Chan, J.W.; Lieu, D.K. Same-Single-Cell Analysis of Pacemaker-Specific Markers in Human Induced Pluripotent Stem Cell-Derived Cardiomyocyte Subtypes Classified by Electrophysiology. Stem Cells 2016, 34, 2670–2680. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.L.; Edwards, A.V.; Juraszek, A.L.; Phelps, A.; Wessels, A.; Burch, J.B. A GATA-6 gene heart-region-specific enhancer provides a novel means to mark and probe a discrete component of the mouse cardiac conduction system. Mech. Dev. 2001, 108, 105–119. [Google Scholar] [CrossRef]
- Sun, C.; Yu, D.; Ye, W.; Liu, C.; Gu, S.; Sinsheimer, N.R.; Song, Z.; Li, X.; Chen, C.; Song, Y. The short stature homeobox 2 (Shox2)-bone morphogenetic protein (BMP) pathway regulates dorsal mesenchymal protrusion development and its temporary function as a pacemaker during cardiogenesis. J. Biol. Chem. 2015, 290, 2007–2023. [Google Scholar] [CrossRef] [Green Version]
- Ionta, V.; Liang, W.; Kim, E.H.; Rafie, R.; Giacomello, A.; Marbán, E.; Cho, H.C. SHOX2 overexpression favors differentiation of embryonic stem cells into cardiac pacemaker cells, improving biological pacing ability. Stem Cell Rep. 2015, 4, 129–142. [Google Scholar] [CrossRef] [Green Version]
- Polonchuk, L. Toward a new gold standard for early safety: Automated temperature-controlled hERG test on the PatchLiner®. Front. Pharmacol. 2012, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Herron, T.J.; Lee, P.; Jalife, J. Optical imaging of voltage and calcium in cardiac cells & tissues. Circ. Res. 2012, 110, 609–623. [Google Scholar]
- Smith, N.A.; Kress, B.T.; Lu, Y.; Chandler-Militello, D.; Benraiss, A.; Nedergaard, M. Fluorescent Ca(2+) indicators directly inhibit the Na,K-ATPase and disrupt cellular functions. Sci. Signal. 2018, 11, eaal2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grienberger, C.; Konnerth, A. Imaging calcium in neurons. Neuron 2012, 73, 862–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankaranarayanan, S.; De Angelis, D.; Rothman, J.E.; Ryan, T.A. The use of pHluorins for optical measurements of presynaptic activity. Biophys. J. 2000, 79, 2199–2208. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Han, Z.; Platisa, J.; Wooltorton, J.R.; Cohen, L.B.; Pieribone, V.A. Single action potentials and subthreshold electrical events imaged in neurons with a fluorescent protein voltage probe. Neuron 2012, 75, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Hitoshi, N.; Ken-ichi, Y.; Jun-ichi, M. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef]
- Goldfracht, I.; Efraim, Y.; Shinnawi, R.; Kovalev, E.; Huber, I.; Gepstein, A.; Arbel, G.; Shaheen, N.; Tiburcy, M.; Zimmermann, W.H.; et al. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 2019, 92, 145–159. [Google Scholar] [CrossRef]
- Jares-Erijman, E.A.; Jovin, T.M. FRET imaging. Nat. Biotechnol. 2003, 21, 1387–1395. [Google Scholar] [CrossRef]
- Akerboom, J.; Rivera, J.D.; Guilbe, M.M.; Malave, E.C.; Hernandez, H.H.; Tian, L.; Hires, S.A.; Marvin, J.S.; Looger, L.L.; Schreiter, E.R. Crystal structures of the GCaMP calcium sensor reveal the mechanism of fluorescence signal change and aid rational design. J. Biol. Chem. 2009, 284, 6455–6464. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Sawano, A.; Park, E.S.; Miyawaki, A. Circularly permuted green fluorescent proteins engineered to sense Ca2+. Proc. Natl. Acad. Sci. USA 2001, 98, 3197–3202. [Google Scholar] [CrossRef] [Green Version]
- Deisseroth, K. Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 2015, 18, 1213–1225. [Google Scholar] [CrossRef] [Green Version]
- Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 2003, 100, 13940–13945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, F.; Setozaki, S.; Kowalski, W.J.; Dwenger, M.; Yuan, F.; Tinney, J.P.; Nakane, T.; Masumoto, H.; Keller, B.B. Progress in the Generation of Multiple Lineage Human-iPSC-Derived 3D-Engineered Cardiac Tissues for Cardiac Repair. In Molecular Mechanism of Congenital Heart Disease and Pulmonary Hypertension; Springer: Singapore, 2020; pp. 353–361. [Google Scholar]
- Schwartz, P.J.; Priori, S.G.; Spazzolini, C.; Moss, A.J.; Vincent, G.M.; Napolitano, C.; Denjoy, I.; Guicheney, P.; Breithardt, G.; Keating, M.T.; et al. Genotype-phenotype correlation in the long-QT syndrome: Gene-specific triggers for life-threatening arrhythmias. Circulation 2001, 103, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buja, L.M. Cardiac repair and the putative role of stem cells. J. Mol. Cell. Cardiol. 2019, 128, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Tiercy, J.-M. How to select the best available related or unrelated donor of hematopoietic stem cells? Haematologica 2016, 101, 680–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rikhtegar, R.; Pezeshkian, M.; Dolati, S.; Safaie, N.; Rad, A.A.; Mahdipour, M.; Nouri, M.; Jodati, A.R.; Yousefi, M. Stem cells as therapy for heart disease: iPSCs, ESCs, CSCs, and skeletal myoblasts. Biomed. Pharmacother. 2019, 109, 304–313. [Google Scholar] [CrossRef]
- Mingliang, R.; Bo, Z.; Zhengguo, W. Stem cells for cardiac repair: Status, mechanisms, and new strategies. Stem Cells Int. 2011, 2011, 310928. [Google Scholar] [CrossRef] [Green Version]
- Lalit, P.A.; Hei, D.J.; Raval, A.N.; Kamp, T.J. Induced pluripotent stem cells for post-myocardial infarction repair: Remarkable opportunities and challenges. Circ. Res. 2014, 114, 1328–1345. [Google Scholar] [CrossRef]
- van Laake, L.W.; Qian, L.; Cheng, P.; Huang, Y.; Hsiao, E.C.; Conklin, B.R.; Srivastava, D. Reporter-based isolation of induced pluripotent stem cell- and embryonic stem cell-derived cardiac progenitors reveals limited gene expression variance. Circ. Res. 2010, 107, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Sottas, V.; Wahl, C.M.; Trache, M.C.; Bartolf-Kopp, M.; Cambridge, S.; Hecker, M.; Ullrich, N.D. Improving electrical properties of iPSC-cardiomyocytes by enhancing Cx43 expression. J. Mol. Cell. Cardiol. 2018, 120, 31–41. [Google Scholar] [CrossRef]
- Denning, C.; Borgdorff, V.; Crutchley, J.; Firth, K.S.; George, V.; Kalra, S.; Kondrashov, A.; Hoang, M.D.; Mosqueira, D.; Patel, A. Cardiomyocytes from human pluripotent stem cells: From laboratory curiosity to industrial biomedical platform. Biochim. Biophys. Acta 2016, 1863, 1728–1748. [Google Scholar] [CrossRef]
- Zhou, H.; Dickson, M.E.; Kim, M.S.; Bassel-Duby, R.; Olson, E.N. Akt1/protein kinase B enhances transcriptional reprogramming of fibroblasts to functional cardiomyocytes. Proc. Natl. Acad. Sci. USA 2015, 112, 11864–11869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Rodriguez, M.; Pabon, L.; Fischer, K.A.; Reinecke, H.; Regnier, M.; Sniadecki, N.J.; Ruohola-Baker, H.; Murry, C.E. Tri-iodo-l-thyronine promotes the maturation of human cardiomyocytes-derived from induced pluripotent stem cells. J. Mol. Cell. Cardiol. 2014, 72, 296–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radisic, M.; Park, H.; Shing, H.; Consi, T.; Schoen, F.J.; Langer, R.; Freed, L.E.; Vunjak-Novakovic, G. Functional assembly of engineered myocardium by electrical stimulation of cardiac myocytes cultured on scaffolds. Proc. Natl. Acad. Sci. USA 2004, 101, 18129–18134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwenger, M.; Kowalski, W.J.; Ye, F.; Yuan, F.; Tinney, J.P.; Setozaki, S.; Nakane, T.; Masumoto, H.; Campbell, P.; Guido, W. Chronic optical pacing conditioning of h-iPSC engineered cardiac tissues. J. Tissue Eng. 2019, 10, 2041731419841748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering adolescence: Maturation of human pluripotent stem cell–derived cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huyer, L.D.; Montgomery, M.; Zhao, Y.; Xiao, Y.; Conant, G.; Korolj, A.; Radisic, M. Biomaterial based cardiac tissue engineering and its applications. Biomed. Mater. 2015, 10, 034004. [Google Scholar] [CrossRef]
- Mannhardt, I.; Breckwoldt, K.; Letuffe-Brenière, D.; Schaaf, S.; Schulz, H.; Neuber, C.; Benzin, A.; Werner, T.; Eder, A.; Schulze, T. Human engineered heart tissue: Analysis of contractile force. Stem Cell Rep. 2016, 7, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Giacomelli, E.; Bellin, M.; Sala, L.; Van Meer, B.J.; Tertoolen, L.G.; Orlova, V.V.; Mummery, C.L. Three-dimensional cardiac microtissues composed of cardiomyocytes and endothelial cells co-differentiated from human pluripotent stem cells. Development 2017, 144, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Goldfracht, I.; Protze, S.; Shiti, A.; Setter, N.; Gruber, A.; Shaheen, N.; Nartiss, Y.; Keller, G.; Gepstein, L. Generating ring-shaped engineered heart tissues from ventricular and atrial human pluripotent stem cell-derived cardiomyocytes. Nat. Commun. 2020, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Nunes, S.S.; Miklas, J.W.; Liu, J.; Aschar-Sobbi, R.; Xiao, Y.; Zhang, B.; Jiang, J.; Massé, S.; Gagliardi, M.; Hsieh, A. Biowire: A platform for maturation of human pluripotent stem cell–derived cardiomyocytes. Nat. Methods 2013, 10, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Maia-Joca, R.P.M.; Ong, C.S.; Wilson, I.; DiSilvestre, D.; Tomaselli, G.F.; Reich, D.H. Enhancement of human iPSC-derived cardiomyocyte maturation by chemical conditioning in a 3D environment. J. Mol. Cell. Cardiol. 2020, 138, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, E.; Meraviglia, V.; Campostrini, G.; Cochrane, A.; Cao, X.; van Helden, R.W.; Garcia, A.K.; Mircea, M.; Kostidis, S.; Davis, R.P. Human-iPSC-Derived Cardiac Stromal Cells Enhance Maturation in 3D Cardiac Microtissues and Reveal Non-cardiomyocyte Contributions to Heart Disease. Cell Stem Cell 2020, 26, 862–879.e11. [Google Scholar] [CrossRef] [PubMed]
- Bargehr, J.; Ong, L.P.; Colzani, M.; Davaapil, H.; Hofsteen, P.; Bhandari, S.; Gambardella, L.; Le Novère, N.; Iyer, D.; Sampaziotis, F. Epicardial cells derived from human embryonic stem cells augment cardiomyocyte-driven heart regeneration. Nat. Biotechnol. 2019, 37, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Lemme, M.; Braren, I.; Prondzynski, M.; Aksehirlioglu, B.; Ulmer, B.M.; Schulze, M.L.; Ismaili, D.; Meyer, C.; Hansen, A.; Christ, T. Chronic intermittent tachypacing by an optogenetic approach induces arrhythmia vulnerability in human engineered heart tissue. Cardiovasc. Res. 2020, 116, 1487–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazelbaker, D.Z.; Beccard, A.; Mazzucato, P.; Angelini, G.; Messana, A.; Lam, D.; Eggan, K.; Barrett, L.E. A multiplexed gRNA piggyBac transposon system facilitates efficient induction of CRISPRi and CRISPRa in human pluripotent stem cells. Sci. Rep. 2020, 10, 635. [Google Scholar] [CrossRef] [Green Version]
- Truong, V.A.; Hsu, M.N.; Kieu Nguyen, N.T.; Lin, M.W.; Shen, C.C.; Lin, C.Y.; Hu, Y.C. CRISPRai for simultaneous gene activation and inhibition to promote stem cell chondrogenesis and calvarial bone regeneration. Nucleic Acids Res. 2019, 47, e74. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Method | Advantage | Disadvantage |
|---|---|---|
| (1) Transient fluorescent reporter lines |
|
|
| (2) Random integration of a fluorescent reporter transgene |
|
|
| (3) Targeted integration of a fluorescent reporter transgene |
|
|
| (3.1) Homologous recombination |
|
|
| (3.2) Advanced genome engineering technologies |
|
|
| (3.2.1) Safe habour site targeting |
|
|
| (3.2.2) Specific locus of interest targeting |
|
|
| Reporter Construct | Type of Promoter Used | Labelled Cell Population | Utility | Host PSC System | Transgenesis Method | Ref |
|---|---|---|---|---|---|---|
| Bry-GFP | Endogenous Bry | Mesoderm | Tracking mesodermal cell differentiation towards cardiogenic and haematopoietic lineages | mESC | HR by electroporation of the BAC-derived targeting vector into mESCs | [40] |
| MESP1-mCherry/NKX2.5-eGFP | Endogenous MESP1 and NKX2.5 | Cardiac mesoderm (MESP1) and CPCs (NKX2.5) | Tracking of transition of cardiac mesoderm toward CPCs and subsequent CM differentiation | hESC | HR by electroporation of BAC-derived targeting vector into hESCs | [41] |
| NKX2.5-IRES- TagRed/TBX5-IRES-Clover | Endogenous NXK2.5 and TBX5 | CPC subsets | Identification and characterisation of presumptive FHF, SHF, pro-pericardial and pre-endothelial progenitors | hiPSC | CRISPR/Cas9 editing in hiPSC at the NKX2.5 and TBX5 loci using plasmid-based lipofection | [45] |
| ISL1-cre dsRed | Endogenous ISL1 | ISL1+ CPC | Characterisation of ISL1+ CPC derivation | hESC | HR by electroporation of BAC-derived ISL1-cre vector into hESCs followed by transfection of floxed dsRed vector | [44] |
| NKX2.5-GFP | Endogenous NKX2.5 | NKX2.5+ CPC/CMs | Isolation of NKX2.5+ CPC and CMs | hESC | HR by electroporation of BAC-derived targeting vector | [47] |
| MYH6-GFP | Ectopic MYH6 | Beating CMs | Purification of beating CMs and development of PSC-CM differentiation protocol | hESCs and hiPSCs | Lentiviral transduction/ random integration of the transgene into differentiating hPSCs | [48] |
| MYH6-eGFP | Ectopic MYH6 | CM progenitor /beating CMs | Identification of early CM progenitor and purification of CMs | hESC | Lentiviral transduction/random integration of the transgene into hESCs | [49] |
| MYH6-mCherry | Endogenous MYH6 | Beating CMs | Purification of beating CMs for cardiotoxicity evaluation | hESC | CRISPR/Cas9 editing in hiPSC at the MYH6 locus using plasmid-based nucleofection | [50] |
| ACTC-mCherry-WPRE-EF1-neo¥ | Ectopic ACTC | Beating CMs | Purification of CMs | hiPSC | Lentiviral transduction/random integration of the transgene into hiPSCs | [51] |
| TTN-GFP | Endogenous TTN | Beating CMs | Purification of beating CMs and, potentially, study of sarcomere functions | hiPSC | CRISPR/Cas9 editing in hiPSC at the TTN locus | [52] |
| NCX1-eGFP-WPRE | Ectopic NCX1 | Beating CMs | Purification of beating CMs | hiPSC and hESC | Lentiviral transduction/random integration of the transgene into hPSCs | [53] |
| MYL2-dsRed | Ectopic MYL2 | Ventricular-like CMs | Purification of ventricular-like CMs | hESC | Lentiviral transduction/ random integration of the transgene into hESCs | [54] |
| MYL2-eGFP | Ectopic MYL2 | Ventricular-like CMs | Purification of ventricular-like CMs | hESC | Lentriviral transduction/ random integration of the transgene into hESCs | [55] |
| Myl2-eGFP | Ectopic Myl2 promoter + human CMVehc | Ventricular-like CMs | Purification of ventricular-like CMs for cell transplantation study | mES | Random integration of the transgene into mESCs via electroporation delivery | [56] |
| MYL2-GFPφ | Ectopic MYL2 | Ventricular-like CMs | Purification of ventricular-like CMs | hiPSC | Adenoviral transduction of the transgene into hiPSCs | [57] |
| MYL2-VSFP | Ectopic MYL2 promoter-enhancer | Ventricular-like CMs | Purification of ventricular-like CMs and study of optical AP | hiPSC | Lentiviral transduction/random integration of the transgene into hiPSC | [58] |
| MYL7-GFPφ | Ectopic MYL7 | Atrial-like CMs | Identification of atrial-like CMs | hiPSC | Adenoviral transduction/random integration of the transgene into hiPSC | [57] |
| SLN-tdTomato | Endogenous SLN | Atrial-like CMs | Purification of atrial-like CMs | hiPSC | HR by electroporation of recombineered BAC DNA into hiPSCs | [59] |
| SLN-VSFP | Ectopic SLN | Atrial-like CMs | Purification of atrial-like CMs and study of optical AP | hiPSC | Lentiviral transduction/random integration of the transgene into hiPSC | [58] |
| NKX2.5-GFP/NR2F2-mCherry | Endogenous NR2F2 and NKX2.5 | Atrial-like CMs | Identification and purification of atrial-like cells and functional study of NR2F2 | hESC | CRISPR/Cas9 targeting (NR2F2-mCherry) into NKX2.5-GFP hESC reporter line [47] | [60] |
| cGATA6-eGFP | Ectopic cGATA6 promoter-enhancer | Nodal-like cells | Identification and purification of nodal-like cells and development of PSC-derived nodal cell differentiation protocol | hESC | Lentriviral transduction/ random integration of the transgene into differentiated hESC-CMs | [61] |
| SHOX2-VSFP | Ectopic SHOX2 | Nodal-like cells | Purification of nodal-like CMs and study of optical AP | hiPSC | Lentiviral transduction/random integration of the transgene into hiPSC | [58] |
| CAG-Arclight | Synthetic CAG | Ubiquitous | Overexpression of Arclight GEVI for optical AP recording in normal and pathological condition in engineered heart tissue | hiPSC | Lentiviral transduction/random integration of the transgene into healthy individual derived hiPSCs and long QT2 patient-derived hiPSCs | [62] |
| TNNT2-GCaMP5G | Ectopic TNNT2 | Beating CMs | Overexpression of GCAPMP5 GECI for optical CT recording in normal and pathological condition in engineered heart tissue | hiPSC | Lentiviral transduction/random integration of the transgene into healthy individual derived hiPSCs and catecholaminergic polymorphic ventricular tachycardia type 2 patient-derived hiPSCs | [62] |
| CaViar (CMV-Arch(D95N)-linker-GCaMP5G-WPRE) | Ectopic CMV promoter-enhancer | Ubiquitous | Overexpression of Arch(D95N) GEVI and GCaPMP5 GECI for simultaneous recordings of AP and CT | hiPSC | Lentiviral transduction/random integration of the transgene into hiPSC | [62] |
| CaViar (CMV-QuasAr2-linker-GCaMP6f-WPRE) OR CaMKIIa-Cheriff-eGFP | Ectopic CMV promoter-enhancer OR ectopic Ca2+/calmodulin-dependent protein kinase II (CaMKIIa) promoter | Ubiquitous | Overexpression of QuasAr2 GEVI and GCaMP5f GECI for simultaneous optical AP and CT recordings in combination with overexpression of optogenetic Cheriff for synchronising CM contraction | hiPSC | Lentiviral transduction/random integration of the transgene into differentiated hiPSC-CMs | [63] |
| CAG-hChR2-mCherryφ | Synthetic CAG | Ubiquitous | Transient overexpression of the optogenetic ChR2 for frequency-dependent drug screening purpose | hiPSC | Adenoviral transduction of the transgene vector into differentiated hiPSC-CMs | [64] |
| CAG-GCaMP3 OR CAG-GCaMP6s OR GAG-G-GECO1.0 | Synthetic CAG | Ubiquitous | Overexpression of a different GECI version for optimising optical CT imaging in rhesus iPSC-CMs | Rhesus iPSC | CRISPR/Cas9 editing in rhiPSC at the AAVS1 locus via plasmid-based chemical transfection | [65] |
| CMV-tri reporter | Ectopic CMV | Ubiquitous | Overexpression of luciferase (for bioluminescent cell tracking after transplantation), mRFP (purification of transduced cells) and thymidine kinase (following dividing cells and tumouriginicity) | miPSC | Lentiviral transduction/random integration of the transgene into miPSC | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sontayananon, N.; Redwood, C.; Davies, B.; Gehmlich, K. Fluorescent PSC-Derived Cardiomyocyte Reporter Lines: Generation Approaches and Their Applications in Cardiovascular Medicine. Biology 2020, 9, 402. https://doi.org/10.3390/biology9110402
Sontayananon N, Redwood C, Davies B, Gehmlich K. Fluorescent PSC-Derived Cardiomyocyte Reporter Lines: Generation Approaches and Their Applications in Cardiovascular Medicine. Biology. 2020; 9(11):402. https://doi.org/10.3390/biology9110402
Chicago/Turabian StyleSontayananon, Naeramit, Charles Redwood, Benjamin Davies, and Katja Gehmlich. 2020. "Fluorescent PSC-Derived Cardiomyocyte Reporter Lines: Generation Approaches and Their Applications in Cardiovascular Medicine" Biology 9, no. 11: 402. https://doi.org/10.3390/biology9110402