Identification of Metabolite and Lipid Profiles in a Segregating Peach Population Associated with Mealiness in Prunus persica (L.) Batsch

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
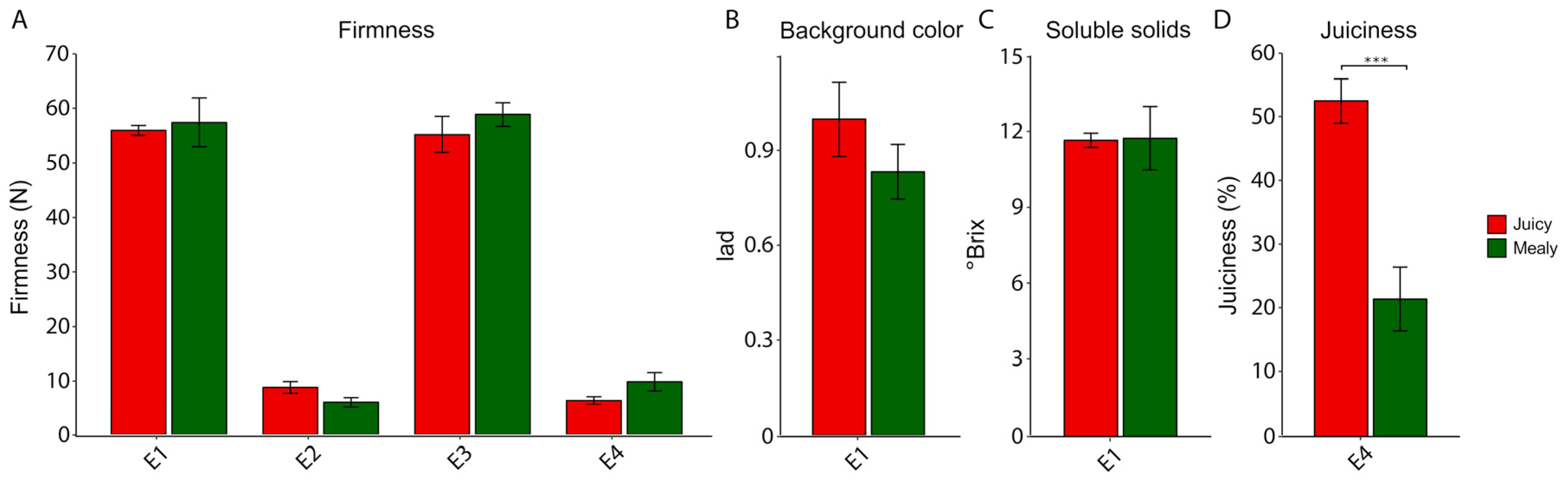
2.1. Phenotypic Parameters in Response to Chilling Injury
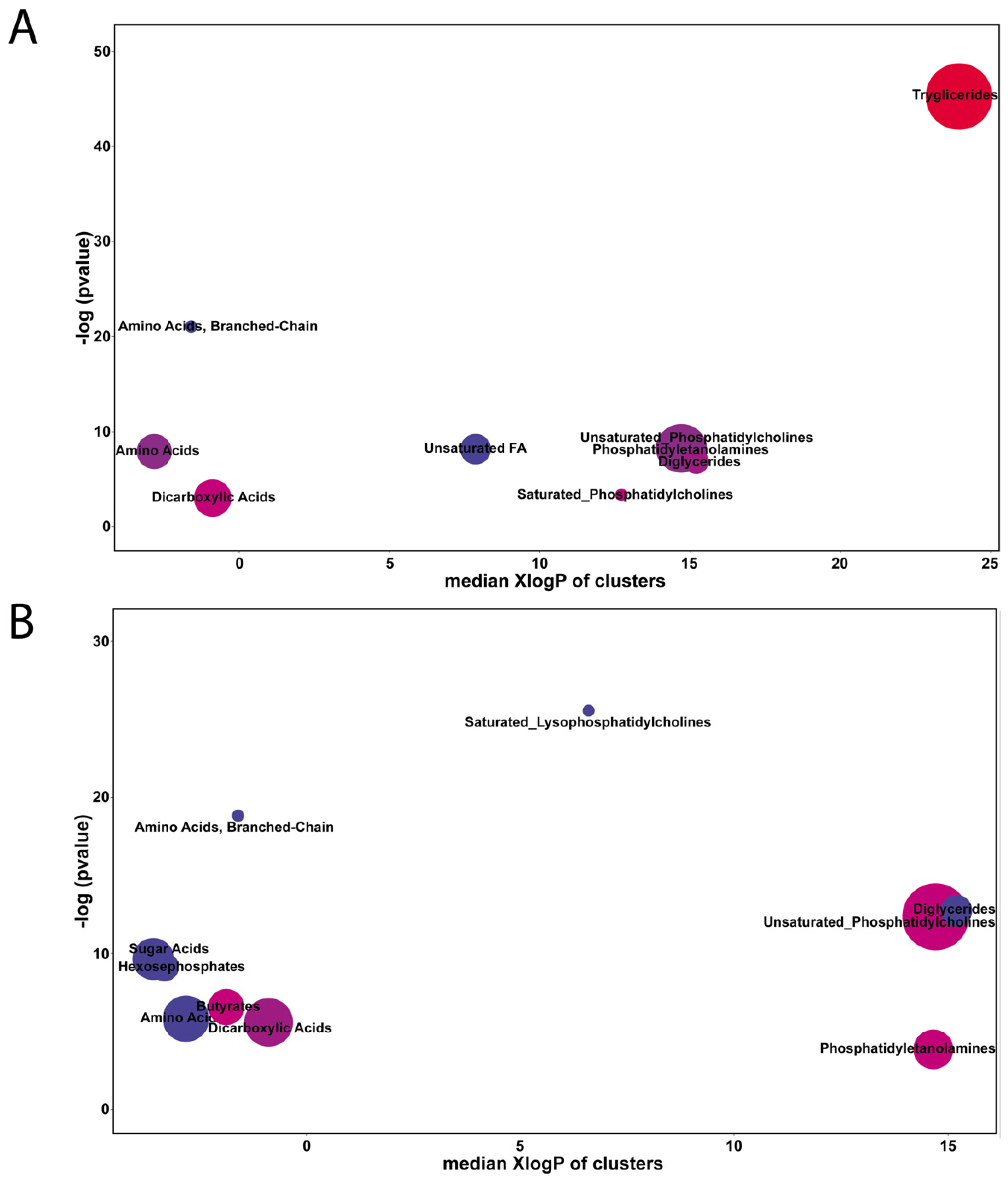
2.2. Comparison of Differentially Accumulated Metabolites and Lipids
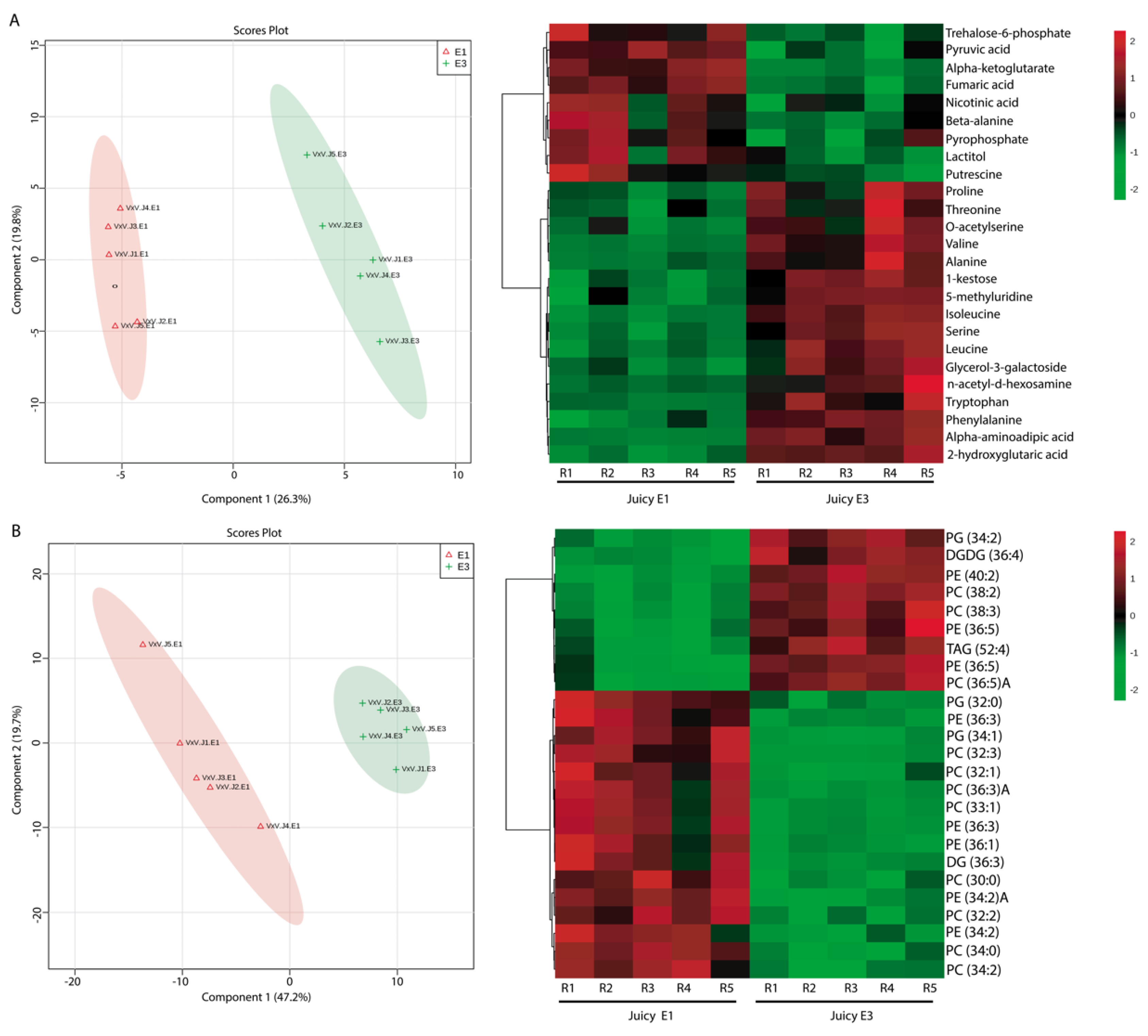
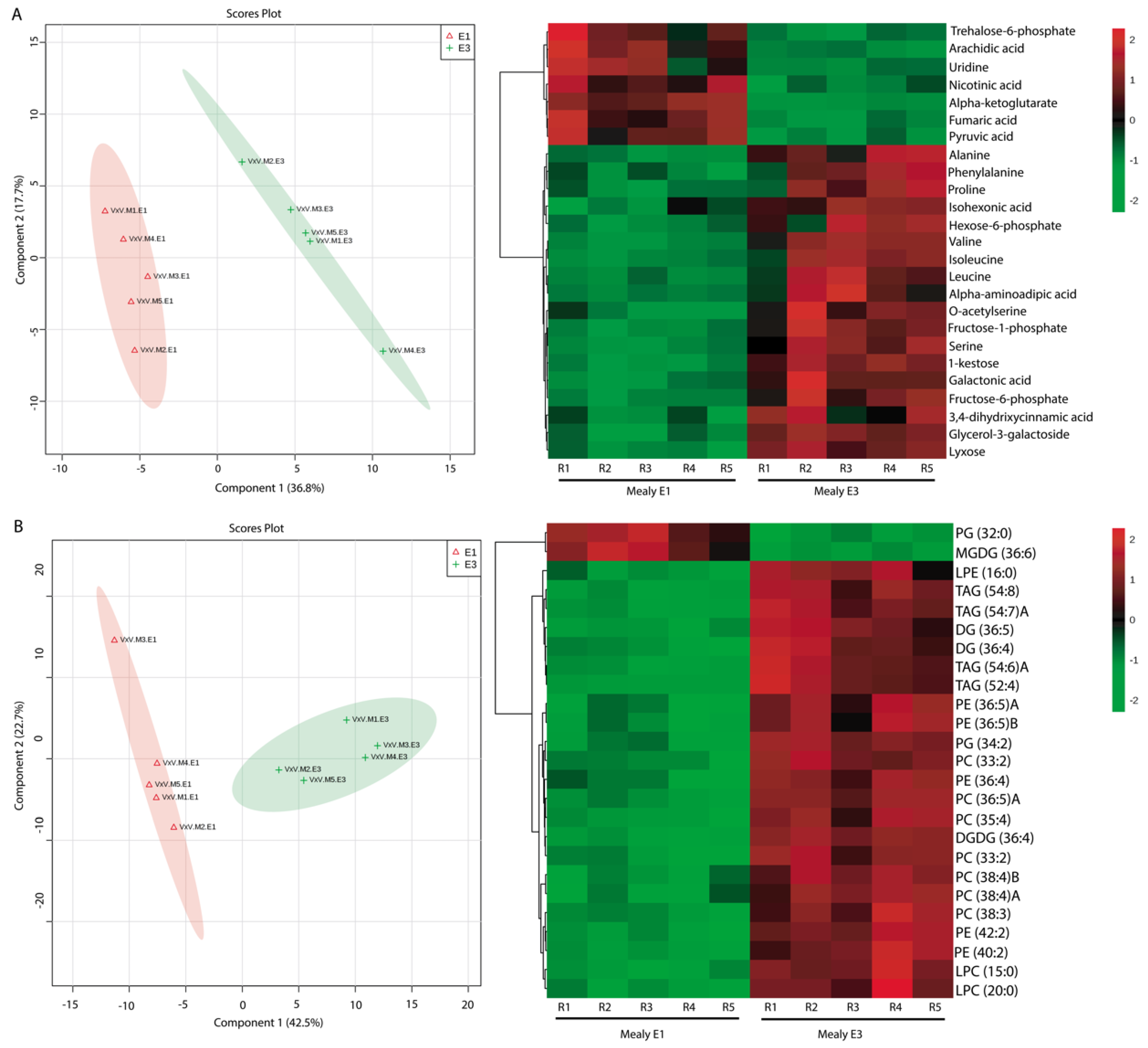
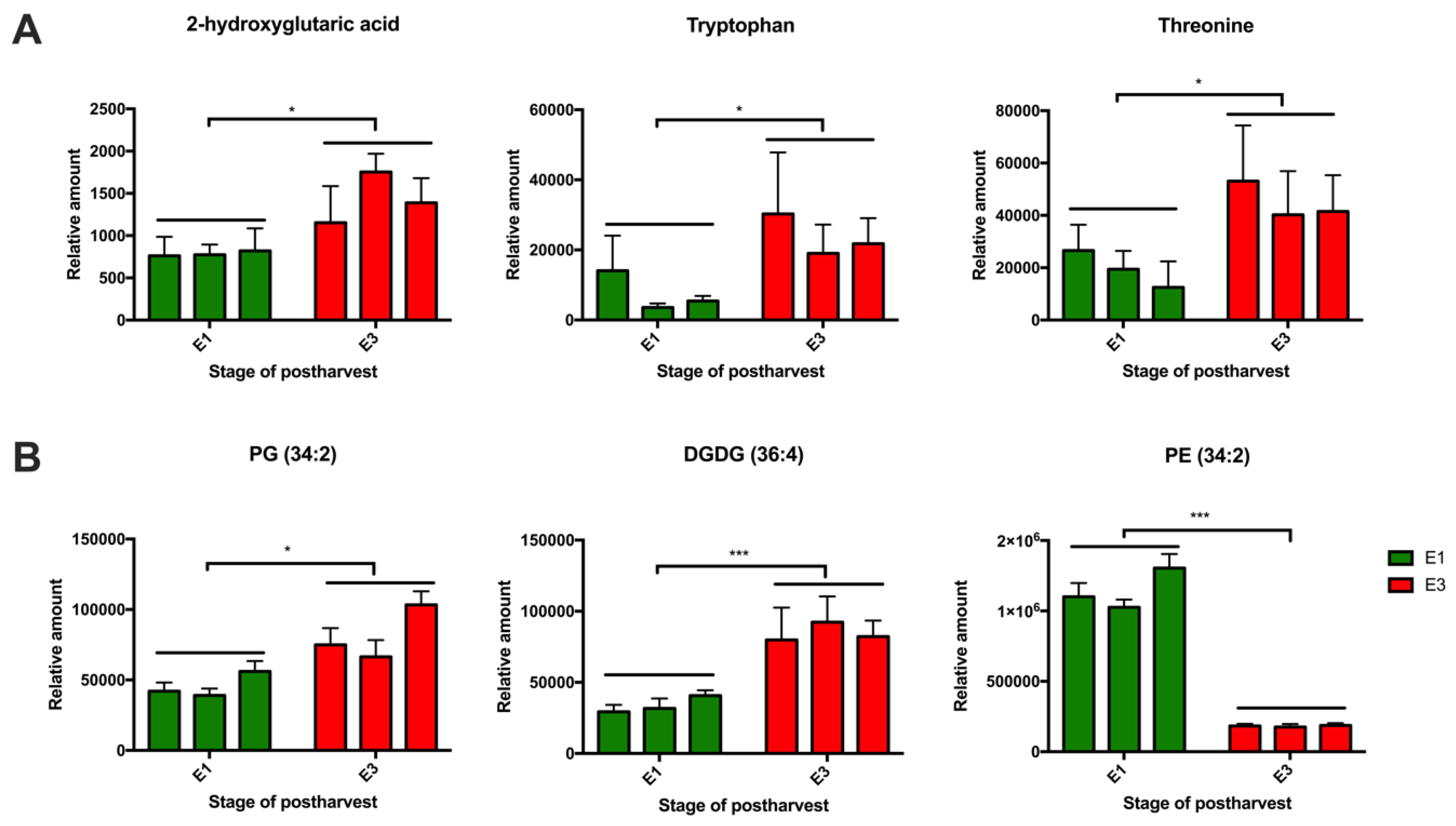
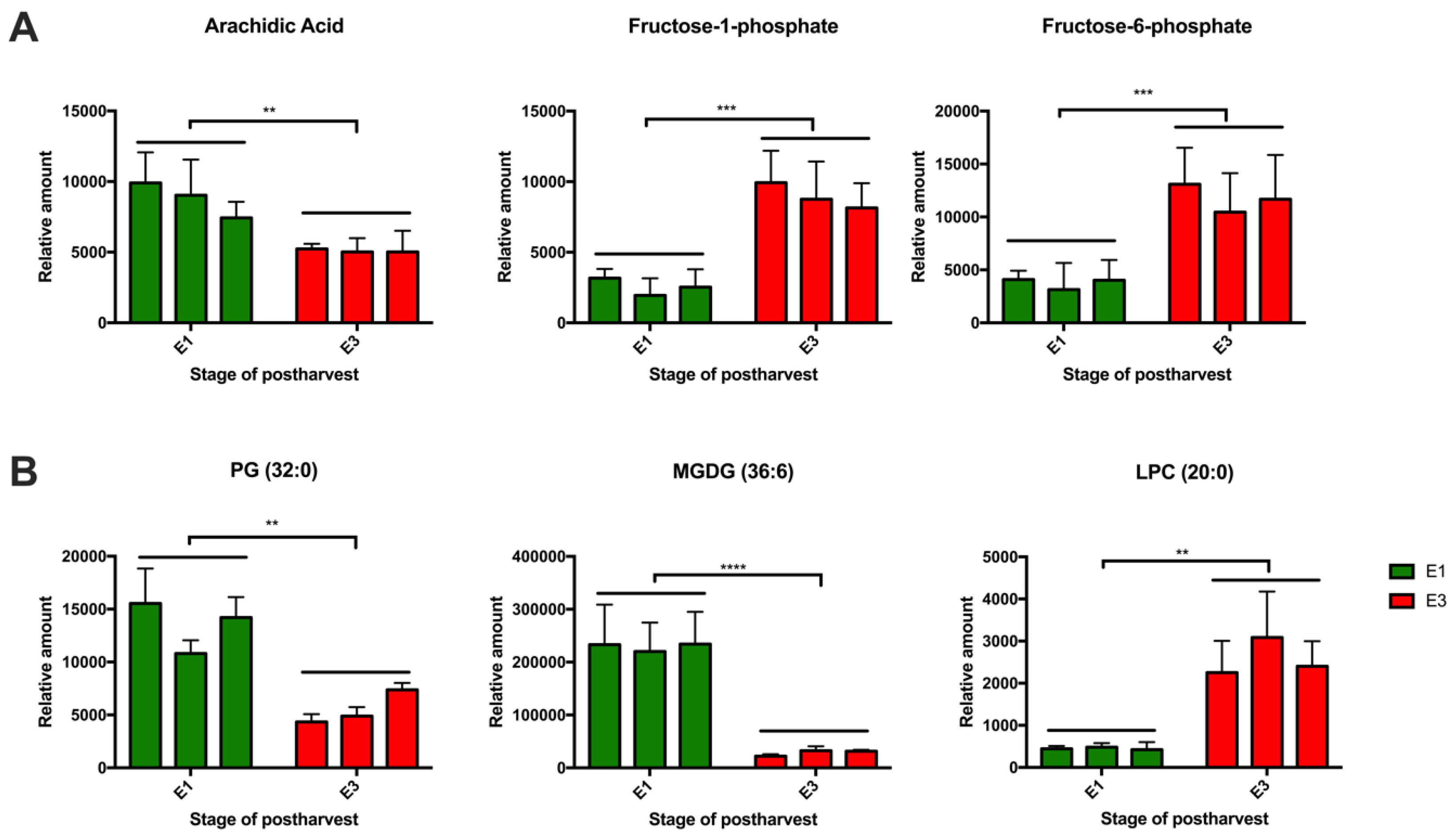
2.3. Effect of Cold Storage on the Metabolic and Lipid Profiles
2.4. Metabolites and Lipids Associated with Cold Stress and Possible Candidate Biomarkers of Mealiness
3. Materials and Methods
3.1. Plant Material and Phenotyping
3.2. Metabolite and Lipid Extraction
3.3. Metabolite and Lipid Analysis
3.4. Metabolic Pathway Assessment
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#home (accessed on 17 December 2019).
- Lurie, S.; Crisosto, C.H. Chilling injury in peach and nectarine. Postharvest Biol. Technol. 2005, 37, 195–208. [Google Scholar] [CrossRef]
- Lill, R.E.; O´Donaghue, E.M.; King, G.A. Postharvest physiology of peaches and nectarines. Hortic. Rev. (Am. Soc. Hortic. Sci) 1989, 11, 413–452. [Google Scholar]
- Peace, C.P.; Crisosto, C.H.; Garner, D.T.; Dandekar, A.M.; Gradziel, T.M.; Bliss, F.A. Genetic control of internal breakdown in peach. In 6th International Peach Symposium; Infante, R., Ed.; Acta Horticulturae: Shanxi, China, 2006; pp. 489–496. [Google Scholar]
- Brummell, D.A.; Cin, V.D.; Lurie, S.; Crisosto, C.H.; Labavitch, J.M. Cell wall metabolism during the development of chilling injury in cold-stored peach fruit: Association of mealiness with arrested disassembly of cell wall Pectins. J. Exp. Bot. 2004, 55, 2041–2052. [Google Scholar] [CrossRef] [PubMed]
- Obenland, D.M.; Carroll, T.R. Mealiness and Pectolytic Activity in Peaches and Nectarines in Response to Heat Treatment and Cold Storage. J. Am. Soc. Hortic. Sci. 2000, 125, 723–728. [Google Scholar] [CrossRef]
- Von Mollendorff, L.J.; de Villiers, O.T. Physiological changes associated with the development of woolliness in ‘peregrine’ peaches during low-temperature storage. J. Hortic. Sci. 1988, 63, 47–51. [Google Scholar] [CrossRef]
- Zhou, H.; Verlag, F.; Dong, L.; Ben-arie, R.; Lurie, S. The role of ethylene in the prevention of chilling injury in nectarines. J. Plant Phisiol. 2001, 158, 55–61. [Google Scholar] [CrossRef]
- Del Pozo, T.; Miranda, S.; Latorre, M.; Olivares, F.; Pavez, L.; Gutierrez, R.; Maldonado, J.; Hinrichsen, P.; Defilippi, B.; Orellana, A.; et al. Comparative transcriptome profiling in a segregating peach population with contrasting juiciness phenotype. J. Agric. Food Chem. 2019, 67, 1598–1607. [Google Scholar] [CrossRef]
- Parkin, K.L.; Marangon, A.; Jackman, R.L.; Yada, R.Y. Chilling injury. A review of possible mechanisms. J. Food Biochem. 1989, 13, 127–153. [Google Scholar] [CrossRef]
- Vizoso, P.; Meisel, L.A.; Tittarelli, A.; Latorre, M.; Saba, J.; Caroca, R.; Maldonado, J.; Cambiazo, V.; Campos-Vargas, R.; Gonzalez, M.; et al. Comparative EST Transcript profiling of peach fruits under different post-harvest conditions reveals candidate genes associated with peach fruit quality. BMC Genom. 2009, 10, 423. [Google Scholar] [CrossRef]
- Nuñez-Lillo, G.; Cifuentes-Esquivel, A.; Troggio, M.; Micheletti, D.; Infante, R.; Campos-Vargas, R.; Orellana, A.; Blanco-Herrera, F.; Meneses, C. Identification of candidate genes associated with mealiness and maturity date in peach [Prunus Persica (L.) Batsch] using QTL analysis and deep sequencing. Tree Genet. Genomes 2015, 11. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Fresnedo-Ramírez, J.; Parfitt, D.E.; Gradziel, T.M.; Crisosto, C.H. Effect prediction of Identified SNPs linked to fruit quality and chilling injury in peach [Prunus Persica (L.) Batsch]. Plant Mol. Biol. 2013, 81, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Sanhueza, D.; Vizoso, P.; Balic, I.; Campos-Vargas, R.; Meneses, C. Transcriptomic analysis of fruit stored under cold conditions using controlled atmosphere in Prunus Persica Cv. “Red Pearl.” Front. Plant Sci. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, C.A.; Monti, L.L.; Gabilondo, J.; Scossa, F.; Valentini, G.; Budde, C.O.; Lara, M.V.; Fernie, A.R.; Drincovich, M.F. Differential metabolic rearrangements after cold storage are correlated with chilling injury resistance of peach fruits. Front. Plant Sci. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hekneby, M.; Antolin, M.C.; Sanchez-Diaz, M. Frost resistance and biochemical changes during cold acclimation in different annual legumes. Environ. Exp. Bot. 2006, 55, 305–314. [Google Scholar] [CrossRef]
- Patton, A.J.; Cunningham, S.M.; Volenec, J.J.; Reicher, Z.J. Differences in freeze tolerance of Zoysiagrasses: II. Carbohydrate and proline accumulation. Crop Sci. 2007, 47, 2170–2181. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Barupal, D.K.; Fiehn, O. Chemical Similarity Enrichment Analysis (ChemRICH) as alternative to biochemical pathway mapping for metabolomic datasets. Sci. Rep. 2017, 7, 14567. [Google Scholar] [CrossRef]
- Cowan, A.K. Phospholipids as plant growth regulators. Plant Growth Regul. 2006, 48, 97–109. [Google Scholar] [CrossRef]
- Barrero-Sicilia, C.; Silvestre, S.; Haslam, R.P.; Michaelson, L.V. Lipid remodelling: Unravelling the response to cold stress in arabidopsis and its extremophile relative eutrema salsugineum. Plant Sci. 2017, 263, 194–200. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, S. Crucial contribution of membrane lipids unsaturation to acquisition of chilling-tolerance in peach fruit stored at 0 ° C. Food Chem. 2009, 115, 405–411. [Google Scholar] [CrossRef]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 1029–1048. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, W.; Li, M.; Welti, R. Profiling lipid changes in plant response to low temperatures. Physiol. Plant. 2006, 126, 90–96. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, S.; Dai, H.; Kong, X.; Hao, J.; Wang, S.; Zhou, X.; Zhao, Y.; Wei, B.; Cheng, S.; et al. Changes in membrane lipid metabolism accompany pitting in blueberry during refrigeration and subsequent storage at room temperature. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses: Role of phospholipase Dα in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Schwender, J.; Ohlrogge, J.; Shachar-hill, Y. Understanding flux in plant metabolic networks. Curr. Opin. Plant Biol. 2004, 7, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Aziz, T.; Hussain, M.; Rehman, H.; Jabran, K.; Khan, M.B. Glycinebetaine improves chilling tolerance in hybrid maize. J. Agron. Crop Sci. 2008, 194, 152–160. [Google Scholar] [CrossRef]
- Kaur, G.; Kumar, S.; Thakur, P.; Malik, J.A.; Bhandhari, K.; Sharma, K.D.; Nayyar, H. Involvement of Proline in response of chickpea (Cicer Arietinum L.) to chilling stress at reproductive stage. Sci. Hortic. (Amsterdam) 2011, 128, 174–181. [Google Scholar] [CrossRef]
- Nayyar, H. Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum Aesti v Um) and Maize (Zea Mays) as affected by calcium and its antagonists. Environ. Exp. Bot. 2003, 50, 253–264. [Google Scholar] [CrossRef]
- Nayyar, H.; Bains, T.; Kumar, S. Low temperature induced floral abortion in chickpea: Relationship to abscisic acid and cryoprotectants in reproductive organs. Environ. Exp. Bot. 2005, 53, 39–47. [Google Scholar] [CrossRef]
- Nayyar, H.; Bains, T.S.; Kumar, S. Chilling stressed chickpea seedlings: Effect of cold acclimation, calcium and abscisic acid on cryoprotective solutes and oxidative damage. Environ. Exp. Bot. 2005, 54, 275–285. [Google Scholar] [CrossRef]
- Nayyar, H.; Chander, S. Protective effects of polyamines against oxidative stress induced by water and cold stress in chickpea. J. Agron. Crop Sci. 2004, 190, 355–365. [Google Scholar] [CrossRef]
- Nayyar, H.; Bains, T.S.; Kumar, S.; Kaur, G. Chilling effects during seed filling on accumulation of seed reserves and yield of chickpea. J. Sci. Food Agric. 2005, 85, 1925–1930. [Google Scholar] [CrossRef]
- Nayyar, H.; Chander, K.; Kumar, S.; Bains, T.; Nayyar, H.; Chander, K.; Kumar, S.; Glycine, T.B. Glycine betaine mitigates cold stress damage in chickpea. Agron. Sustain. Dev. 2005, 25, 381–388. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Strawn, L.; Babb, A.; Testerink, C.; Kooijman, E.E.; Zachowski, A.; Pierre, U. The physical chemistry of the enigmatic phospholipid diacylglycerol pyrophosphate. Front. Plant Sci. 2012, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, C.A.; Brotman, Y.; Monti, L.L.; Gabilondo, J.; Budde, C.O.; Lara, M.V.; Fernie, A.R.; Drincovich, M.F. Differential lipidome remodeling during postharvest of peach varieties with different susceptibility to chilling injury. Physiol. Plant. 2018, 163, 2–17. [Google Scholar] [CrossRef]
- Amiard, V.; Morvan-Bertrand, A.; Billard, J.; Huault, C.; Keller, F.; Prud´homme, M. Fructans, but not the sucrosyl-galactosides, raffinose and loliose, are affected by drought stress in perennial ryegrass. Plant Physiol. 2003, 132, 2218–2229. [Google Scholar] [CrossRef]
- Del Viso, F.; Casabuono, A.C.; Couto, A.S.; Hopp, H.E.; Puebla, A.F.; Heinz, R.A. Functional characterization of a sucrose: Fructan 6-fructosyltransferase of the cold-resistant grass bromus pictus by heterelogous expression in Pichia Pastoris and Nicotiana Tabacum and its involvement in freezing tolerance. J. Plant Physiol. 2011, 168, 493–499. [Google Scholar] [CrossRef]
- Pontis, H.G. Review fructans and cold stress. J. Plant Physiol. 1989, 134, 148–150. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses, 2nd ed.; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Schnyder, H.; Nelson, C.J. Growth rates and carbohydrate fluxes within the elongation zone of tall fescue leaf blades. Plant Physiol. 1987, 85, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Darbyshire, B.; Allaway, W.G. Soluble carbohydrates in leaf epidermis of. Plant Sci. Lett. 1981, 22, 141–145. [Google Scholar] [CrossRef]
- Valluru, R.; Van den Ende, W. Plant fructans in stress environments: Emerging concepts and future prospects. J. Exp. Bot. 2008, 59, 2905–2916. [Google Scholar] [CrossRef]
- Figueroa, C.; Lunn, J. A Tale of two sugars: Trehalose 6-phosphate and sucrose. Plant Physiol. 2016, 172, 7–27. [Google Scholar] [CrossRef] [PubMed]
- Iordachescu, M.; Imai, R. Trehalose biosynthesis in response to abiotic stresses. J. Integr. Plant Biol. 2008, 50, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Sundaramurthi, P.; Patapoff, T.W.; Suryanarayanan, R. Crystallization of trehalose in frozen solutions and its phase behavior during drying. Pharm. Res. 2010, 27, 2374–2383. [Google Scholar] [CrossRef] [PubMed]
- Brizzolara, S.; Hertog, M.; Tosetti, R.; Nicolai, B.; Tonutti, P. Metabolic responses to low temperature of three peach fruit cultivars differently sensitive to cold storage. Front. Plant Sci. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Kapalka, G. Substances involved in neurotransmission. In Nutritional and Herbal Therapies for Children and Adolescents; Inc, E., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 71–99. [Google Scholar] [CrossRef]
- Almeida, A.M.; Urra, C.; Moraga, C.; Jego, M.; Flores, A.; Meisel, L.; González, M.; Infante, R.; Defilippi, B.G.; Campos-Vargas, R.; et al. Proteomic analysis of a segregant population reveals candidate proteins linked to mealiness in peach. J. Proteom. 2016, 131, 71–81. [Google Scholar] [CrossRef]
- Diehl, R.; Guinn, E.; Capp, M.; Tsodikov, O.; Record, T. Quantifying additive interactions of the osmolyte proline with individual functional groups of proteins: Comparisons with urea and glycine betaine, interpretation of m-values. Biochemistry 2013, 52. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Abdelly, C.; Savouré, A. Plant physiology and biochemistry how reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Law, J.A.; Jacobsen, S.E. Patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2009, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.R.S.S.; Rao, S.; Reddy, K.J.; Theriappan, P.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar]
- Kozukue, N.; Kozuekue, E.; Hirose, T.; Mizuno, S. Acummulation of alanine in chilling-sensitive crops. HortScience 1984, 19, 498–501. [Google Scholar]
- Patterson, B.; Pearson, J.; Payne, L.; Ferguson, I. Metabolic aspects of chilling resistance: Alanine accumulation and glutamate depletion in relation to chilling sensitivity in passionfruit species. Funct. Plant Biol. 1981, 8, 395–403. [Google Scholar] [CrossRef]
- Tsuchida, H.; Kozukue, N.; Han, G.; Choi, S.; Levin, C.E.; Friedman, M. Low-temperature storage of cucumbers induces changes in the organic acid content and in citrate synthase activity. Postharvest Biol. Technol. 2010, 58, 129–134. [Google Scholar] [CrossRef]
- Tsuchida, H.; Dan-hong, C. Changes in pyruvic acid content and GPT activity in chilling-sensitive and nonsensitive crops. HortScience 1990, 25, 952–953. [Google Scholar] [CrossRef]
- Tanou, G.; Minas, I.S.; Scossa, F.; Belghazi, M.; Ganopoulos, I.; Madesis, P.; Fernie, A. Exploring priming responses involved in peach fruit acclimation to cold stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Joshi, V.; Zhangjun, J.J.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: Accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Hincha, D.K.; Oliver, A.E.; Crowe, J.H. The effects of chloroplast lipids on the stability of liposomes during freezing and drying. Biochim. Biophys. 1998, 1368, 150–160. [Google Scholar] [CrossRef][Green Version]
- Abidi, W.; Cantín, C.M.; Jiménez, S.; Giménez, R.; Moreno, M.Á.; Gogorcena, Y. Influence of antioxidant compounds, total sugars and genetic background on the chilling injury susceptibility of a non-melting peach (Prunus Persica (L.) Batsch) progeny. J. Sci. Food Agric. 2015, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shao, X.; Gong, Y.; Zhu, Y.; Wang, H.; Zhang, X.; Yu, D.; Yu, F.; Qiu, Z.; Lu, H. The metabolism of soluble carbohydrates related to chilling injury in peach fruit exposed to cold stress. Postharvest Biol. Technol. 2013, 86, 53–61. [Google Scholar] [CrossRef]
- Luengwilai, K.; Saltveit, M.; Beckles, D.M. Metabolite content of harvested micro-tom tomato (Solanum Lycopersicum L.) fruit is altered by chilling and protective heat-shock treatments as shown by GC–MS metabolic profiling. Postharvest Biol. Technol. 2012, 63, 116–122. [Google Scholar] [CrossRef]
- Steponkus, P.L.; Uemura, M.; Balsamo, R.A.; Arvintet, T.; Lynch, D.V. Transformation of the cryobehavior of rye protoplasts by modifications of the plasma membrane lipid composition. Proc. Natl. Acad. Sci. USA 1988, 85, 9026–9030. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Wei, B.; Gao, Z.; Zhou, Y.; Shi, F.; Zhou, X.; Zhou, Q.; Ji, S. Changes in membrane lipid composition and function accompanying chilling injury in bell peppers. Plant Cell Physiol. 2018, 59, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Moellering, E.R.; Muthan, B.; Benning, C. Freezing tolerance in plants requires lipid remodeling at the outer chloroplast membrane. Science (80) 2010, 330, 226–228. [Google Scholar] [CrossRef]
- Szymanski, J.; Brotman, Y.; Willmitzer, L. Linking gene expression and membrane lipid composition of arabidopsis. Plant Cell 2014, 26, 915–928. [Google Scholar] [CrossRef]
- Infante, R.; Meneses, C.; Crisosto, C.H. Preconditioning treatment maintains taste characteristic perception of ripe ‘September Sun’ peach following cold storage. Int. J. Food Sci. Technol. 2009, 44, 1011–1016. [Google Scholar] [CrossRef]
- Infante, R.; Meneses, C.; Rubio, P.; Seibert, E. Quantitative determination of flesh mealiness in peach [Prunus Persica L. (Batch.)] through paper absorption of free juice. Postharvest Biol. Technol. 2009, 51, 118–121. [Google Scholar] [CrossRef]
- Fiehn, O.; Wohlgemuth, G.; Scholz, M.; Kind, T.; Lee, D.Y.; Lu, Y.; Moon, S.; Nikolau, B. Quality control for plant metabolomics: Reporting MSI-compliant studies. Plant J. 2008, 53, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V. Lipid extraction by Methyl-Tert-Butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lillo-Carmona, V.; Espinoza, A.; Rothkegel, K.; Rubilar, M.; Nilo-Poyanco, R.; Pedreschi, R.; Campos-Vargas, R.; Meneses, C. Identification of Metabolite and Lipid Profiles in a Segregating Peach Population Associated with Mealiness in Prunus persica (L.) Batsch. Metabolites 2020, 10, 154. https://doi.org/10.3390/metabo10040154
Lillo-Carmona V, Espinoza A, Rothkegel K, Rubilar M, Nilo-Poyanco R, Pedreschi R, Campos-Vargas R, Meneses C. Identification of Metabolite and Lipid Profiles in a Segregating Peach Population Associated with Mealiness in Prunus persica (L.) Batsch. Metabolites. 2020; 10(4):154. https://doi.org/10.3390/metabo10040154
Chicago/Turabian StyleLillo-Carmona, Victoria, Alonso Espinoza, Karin Rothkegel, Miguel Rubilar, Ricardo Nilo-Poyanco, Romina Pedreschi, Reinaldo Campos-Vargas, and Claudio Meneses. 2020. "Identification of Metabolite and Lipid Profiles in a Segregating Peach Population Associated with Mealiness in Prunus persica (L.) Batsch" Metabolites 10, no. 4: 154. https://doi.org/10.3390/metabo10040154
APA StyleLillo-Carmona, V., Espinoza, A., Rothkegel, K., Rubilar, M., Nilo-Poyanco, R., Pedreschi, R., Campos-Vargas, R., & Meneses, C. (2020). Identification of Metabolite and Lipid Profiles in a Segregating Peach Population Associated with Mealiness in Prunus persica (L.) Batsch. Metabolites, 10(4), 154. https://doi.org/10.3390/metabo10040154