Harnessing Metabolites as Serum Biomarkers for Liver Graft Pathology Prediction Using Machine Learning

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. Data Analysis
2.3. Integration of Clinical Variables with VIP Metabolites for Prediction of Patient Outcomes Post Transplantation
3. Results
3.1. Alanine, Aspartate, and Glutamate Metabolism Pathway Exhibited Notable Alterations in a Comparative Analysis of MASH (n = 10) and TCMR (n = 18) Patients
3.2. Aminoacyl-tRNA Biosynthesis Emerged as the Pathway Exhibiting the Most Pronounced Alterations When Comparing MASH with Biliary Complications Group
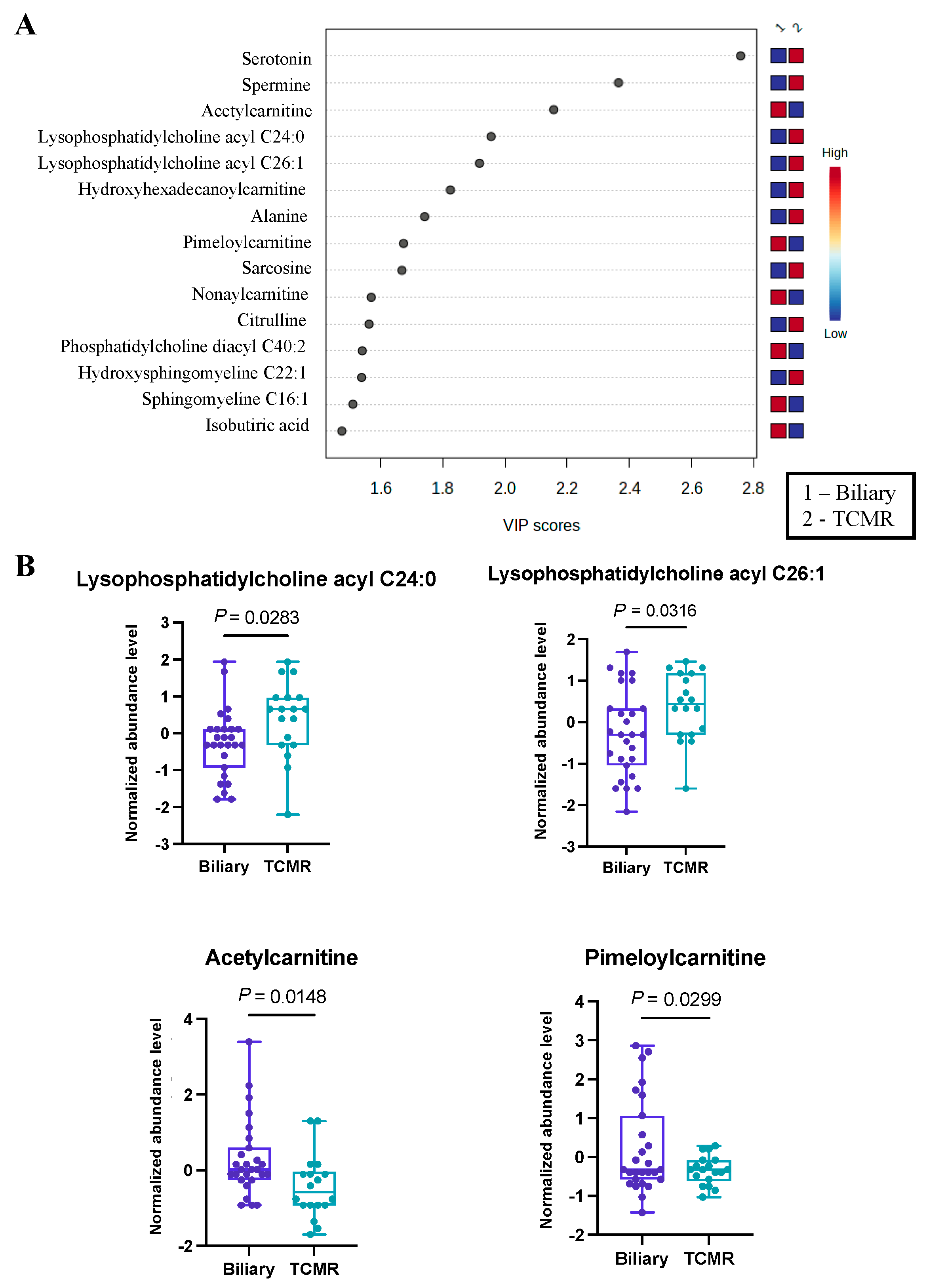
3.3. Significant Alterations in Butanoate (Butyrate) Metabolism Were Revealed in the Biliary vs. TCMR Group Comparison
3.4. Carnitines Were Identified as Differentially Abundant in Our Disease-Wise Sex-Based Analysis
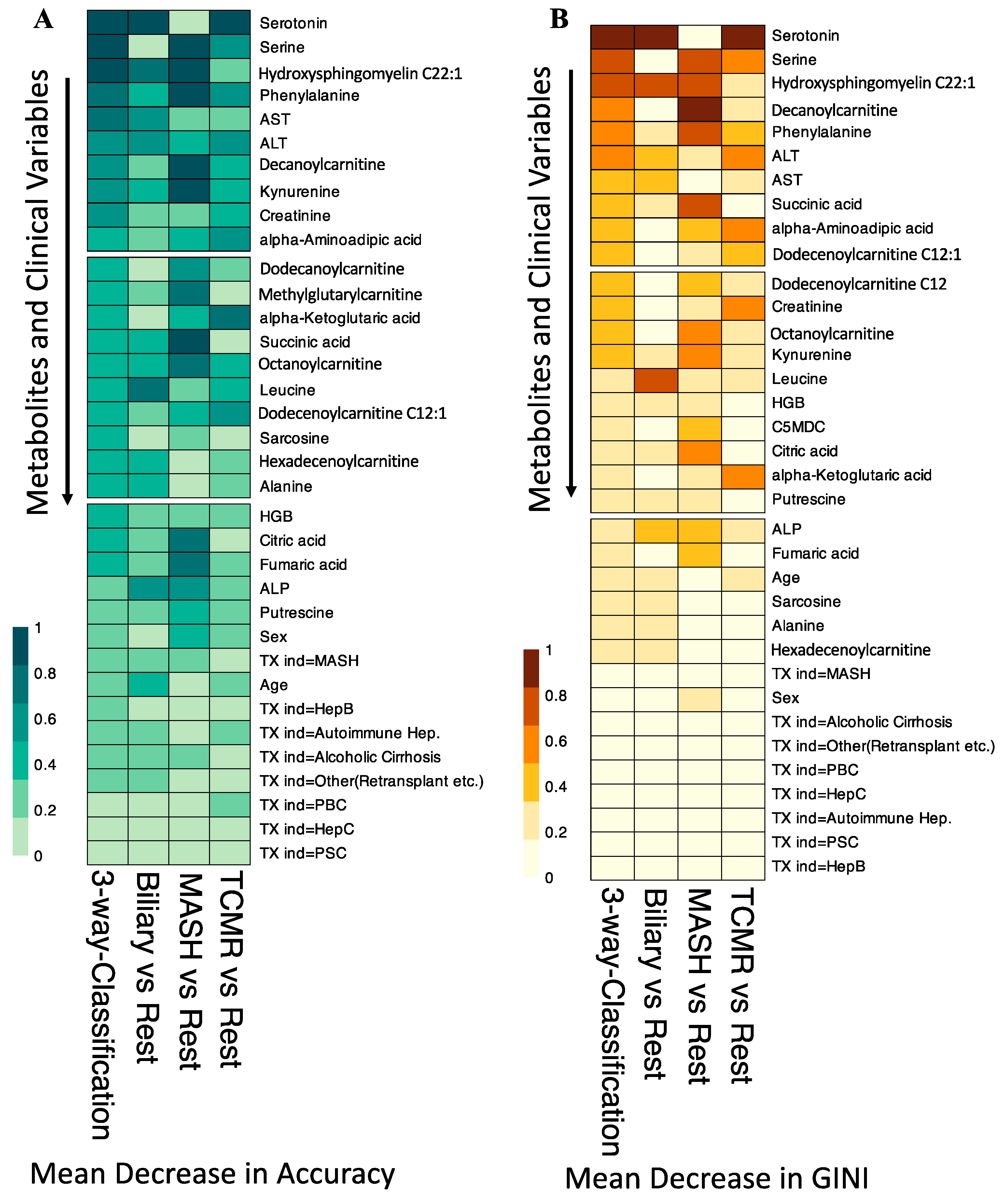
3.5. Integration of Clinical Information with Metabolomics Data
3.6. Comparative Analysis of Integrated Model versus Individual Modalities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statista. 2023. Available online: https://www.statista.com/statistics/398685/liver-transplants-by-world-region/ (accessed on 5 February 2024).
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef]
- Moreno, R.; Berenguer, M. Post-liver transplantation medical complications. Ann. Hepatol. 2006, 5, 77–85. [Google Scholar] [CrossRef]
- Watt, K.D.; Pedersen, R.A.; Kremers, W.K.; Heimbach, J.K.; Charlton, M.R. Evolution of causes and risk factors for mortality post-liver transplant: Results of the NIDDK long-term follow-up study. Am. J. Transplant. 2010, 10, 1420–1427. [Google Scholar] [CrossRef] [PubMed]
- Jadlowiec, C.C.; Morgan, P.E.; Nehra, A.K.; Hathcock, M.A.; Kremers, W.K.; Heimbach, J.K.; Wiesner, R.H.; Taner, T. Not All Cellular Rejections Are the Same: Differences in Early and Late Hepatic Allograft Rejection. Liver Transpl. 2019, 25, 425–435. [Google Scholar] [CrossRef]
- Levitsky, J.; Goldberg, D.; Smith, A.R.; Mansfield, S.A.; Gillespie, B.W.; Merion, R.M.; Lok, A.S.; Levy, G.; Kulik, L.; Abecassis, M.; et al. Acute Rejection Increases Risk of Graft Failure and Death in Recent Liver Transplant Recipients. Clin. Gastroenterol. Hepatol. 2017, 15, 584–593.e582. [Google Scholar] [CrossRef]
- Saeed, N.; Glass, L.; Sharma, P.; Shannon, C.; Sonnenday, C.J.; Tincopa, M.A. Incidence and Risks for Nonalcoholic Fatty Liver Disease and Steatohepatitis Post-liver Transplant: Systematic Review and Meta-analysis. Transplantation 2019, 103, e345–e354. [Google Scholar] [CrossRef]
- Abraham, S.C.; Poterucha, J.J.; Rosen, C.B.; Demetris, A.J.; Krasinskas, A.M. Histologic abnormalities are common in protocol liver allograft biopsies from patients with normal liver function tests. Am. J. Surg. Pathol. 2008, 32, 965–973. [Google Scholar] [CrossRef]
- Sanchez-Fueyo, A.; Restrepo, J.C.; Quinto, L.; Bruguera, M.; Grande, L.; Sanchez-Tapias, J.M.; Rodes, J.; Rimola, A. Impact of the recurrence of hepatitis C virus infection after liver transplantation on the long-term viability of the graft. Transplantation 2002, 73, 56–63. [Google Scholar] [CrossRef]
- Sebagh, M.; Samuel, D.; Antonini, T.M.; Coilly, A.; Degli Esposti, D.; Roche, B.; Karam, V.; Dos Santos, A.; Duclos-Vallee, J.C.; Roque-Afonso, A.M.; et al. Twenty-year protocol liver biopsies: Invasive but useful for the management of liver recipients. J. Hepatol. 2012, 56, 840–847. [Google Scholar] [CrossRef]
- Bhat, M.; Tazari, M.; Sebastiani, G. Performance of transient elastography and serum fibrosis biomarkers for non-invasive evaluation of recurrent fibrosis after liver transplantation: A meta-analysis. PLoS ONE 2017, 12, e0185192. [Google Scholar] [CrossRef]
- Alpna, R.L.; Lisa, R.D.; Roberto, J.F. Liver Biopsy After Liver Transplantation. In Liver Biopsy; Nobumi, T., Ed.; IntechOpen: Rijeka,Croatia, 2012. [Google Scholar]
- Schütz, E.; Fischer, A.; Beck, J.; Harden, M.; Koch, M.; Wuensch, T.; Stockmann, M.; Nashan, B.; Kollmar, O.; Matthaei, J.; et al. Graft-derived cell-free DNA, a noninvasive early rejection and graft damage marker in liver transplantation: A prospective, observational, multicenter cohort study. PLoS Med. 2017, 14, e1002286. [Google Scholar] [CrossRef]
- Levitsky, J.; Kandpal, M.; Guo, K.; Kleiboeker, S.; Sinha, R.; Abecassis, M. Donor-derived cell-free DNA levels predict graft injury in liver transplant recipients. Am. J. Transplant. 2022, 22, 532–540. [Google Scholar] [CrossRef]
- Levitsky, J.; Kandpal, M.; Guo, K.; Zhao, L.; Kurian, S.; Whisenant, T.; Abecassis, M. Prediction of Liver Transplant Rejection With a Biologically Relevant Gene Expression Signature. Transplantation 2022, 106, 1004–1011. [Google Scholar] [CrossRef]
- Idle, J.R.; Gonzalez, F.J. Metabolomics. Cell Metab. 2007, 6, 348–351. [Google Scholar] [CrossRef]
- Cortes, M.; Pareja, E.; Garcia-Canaveras, J.C.; Donato, M.T.; Montero, S.; Mir, J.; Castell, J.V.; Lahoz, A. Metabolomics discloses donor liver biomarkers associated with early allograft dysfunction. J. Hepatol. 2014, 61, 564–574. [Google Scholar] [CrossRef]
- Ferrarini, A.; Di Poto, C.; He, S.; Tu, C.; Varghese, R.S.; Kara Balla, A.; Jayatilake, M.; Li, Z.; Ghaffari, K.; Fan, Z.; et al. Metabolomic Analysis of Liver Tissues for Characterization of Hepatocellular Carcinoma. J. Proteome Res. 2019, 18, 3067–3076. [Google Scholar] [CrossRef]
- Nimer, N.; Choucair, I.; Wang, Z.; Nemet, I.; Li, L.; Gukasyan, J.; Weeks, T.L.; Alkhouri, N.; Zein, N.; Tang, W.H.W.; et al. Bile acids profile, histopathological indices and genetic variants for non-alcoholic fatty liver disease progression. Metabolism 2021, 116, 154457. [Google Scholar] [CrossRef]
- Liu, J.; Geng, W.; Sun, H.; Liu, C.; Huang, F.; Cao, J.; Xia, L.; Zhao, H.; Zhai, J.; Li, Q.; et al. Integrative metabolomic characterisation identifies altered portal vein serum metabolome contributing to human hepatocellular carcinoma. Gut 2022, 71, 1203–1213. [Google Scholar] [CrossRef]
- Kalhan, S.C.; Guo, L.; Edmison, J.; Dasarathy, S.; McCullough, A.J.; Hanson, R.W.; Milburn, M. Plasma metabolomic profile in nonalcoholic fatty liver disease. Metabolism 2011, 60, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Frediani, J.K.; Beyh, Y.S.; Gupta, N.; Westbrook, A.L.; Cleeton, R.; Cordero, M.; Hernandez, A.; Tran, V.; Jones, D.P.; Vos, M.B. Metabolomics profiling in acute liver transplant rejection in a pediatric population. Sci. Rep. 2022, 12, 18663. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, X.P.; Troisi, R.I.; Colle, I.; Geerts, A.; van Vlierberghe, H. Biomarkers for the diagnosis of acute cellular rejection in liver transplant recipients: A review. Hepatol. Res. 2013, 43, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC–HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Chawla, N.V.; Bowyer, K.W.; Hall, L.O.; Kegelmeyer, P.W. SMOTE: Synthetic minority over-sampling technique. J. Artif. Intell. Res. 2002, 16, 321–357. [Google Scholar] [CrossRef]
- van der Maaten, l.; Hinton, G. Visualizing Data using t-SNE. J. Mach. Learn. Res. 2008, 9, 2579–2605. [Google Scholar]
- Mardinoglu, A.; Agren, R.; Kampf, C.; Asplund, A.; Uhlen, M.; Nielsen, J. Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014, 5, 3083. [Google Scholar] [CrossRef]
- Sim, W.C.; Lee, W.; Sim, H.; Lee, K.Y.; Jung, S.H.; Choi, Y.J.; Kim, H.Y.; Kang, K.W.; Lee, J.Y.; Choi, Y.J.; et al. Downregulation of PHGDH expression and hepatic serine level contribute to the development of fatty liver disease. Metabolism 2020, 102, 154000. [Google Scholar] [CrossRef]
- Murr, C.; Grammer, T.B.; Meinitzer, A.; Kleber, M.E.; März, W.; Fuchs, D. Immune activation and inflammation in patients with cardiovascular disease are associated with higher phenylalanine to tyrosine ratios: The ludwigshafen risk and cardiovascular health study. J. Amino Acids 2014, 2014, 783730. [Google Scholar] [CrossRef]
- Sikalidis, A.K. Amino acids and immune response: A role for cysteine, glutamine, phenylalanine, tryptophan and arginine in T-cell function and cancer? Pathol. Oncol. Res. 2015, 21, 9–17. [Google Scholar] [CrossRef]
- Ma, E.H.; Bantug, G.; Griss, T.; Condotta, S.; Johnson, R.M.; Samborska, B.; Mainolfi, N.; Suri, V.; Guak, H.; Balmer, M.L.; et al. Serine Is an Essential Metabolite for Effector T Cell Expansion. Cell Metab. 2017, 25, 345–357. [Google Scholar] [CrossRef]
- Nikolova-Karakashian, M. Alcoholic and non-alcoholic fatty liver disease: Focus on ceramide. Adv. Biol. Regul. 2018, 70, 40–50. [Google Scholar] [CrossRef]
- Ruddell, R.G.; Mann, D.A.; Ramm, G.A. The function of serotonin within the liver. J. Hepatol. 2008, 48, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Gama, J.F.G.; Cardoso, L.; Bisaggio, R.D.C.; Lagrota-Candido, J.; Henriques-Pons, A.; Alves, L.A. Immunological Tolerance in Liver Transplant Recipients: Putative Involvement of Neuroendocrine-Immune Interactions. Cells 2022, 11, 2327. [Google Scholar] [CrossRef]
- Wu, H.; Denna, T.H.; Storkersen, J.N.; Gerriets, V.A. Beyond a neurotransmitter: The role of serotonin in inflammation and immunity. Pharmacol. Res. 2019, 140, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, M.; Caterino, M.; Sotgiu, G.; Ruoppolo, M.; Franconi, F.; Campesi, I. Sex differences in the human metabolome. Biol. Sex Differ. 2022, 13, 30. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhao, H. Role of Carnitine in Non-alcoholic Fatty Liver Disease and Other Related Diseases: An Update. Front. Med. 2021, 8, 689042. [Google Scholar] [CrossRef]
- Ji, M.; Jo, Y.; Choi, S.J.; Kim, S.M.; Kim, K.K.; Oh, B.C.; Ryu, D.; Paik, M.J.; Lee, D.H. Plasma Metabolomics and Machine Learning-Driven Novel Diagnostic Signature for Non-Alcoholic Steatohepatitis. Biomedicines 2022, 10, 1669. [Google Scholar] [CrossRef]
- Mowry, C.J.; Alonso, C.; Iruarrizaga-Lejarreta, M.; Ortiz, P.; Levitsky, J.; Rinella, M. Utility of Metabolomic Biomarkers to Identify Nonalcoholic Fatty Liver Disease in Liver Transplant Recipients. Transplant. Direct 2021, 7, e784. [Google Scholar] [CrossRef]
- Jia, J.; Nie, Y.; Geng, L.; Li, J.; Liu, J.; Peng, Y.; Huang, J.; Xie, H.; Zhou, L.; Zheng, S.S. Identification of HO-1 as a novel biomarker for graft acute cellular rejection and prognosis prediction after liver transplantation. Ann. Transl. Med. 2020, 8, 221. [Google Scholar] [CrossRef] [PubMed]
- Lake, A.D.; Novak, P.; Shipkova, P.; Aranibar, N.; Robertson, D.G.; Reily, M.D.; Lehman-McKeeman, L.D.; Vaillancourt, R.R.; Cherrington, N.J. Branched chain amino acid metabolism profiles in progressive human nonalcoholic fatty liver disease. Amino Acids 2015, 47, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.T.; Cresci, G.A.M. The Immunomodulatory Functions of Butyrate. J. Inflamm. Res. 2021, 14, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Jhun, J.; Lee, S.Y.; Choi, S.; Choi, S.S.; Park, M.S.; Lee, S.Y.; Cho, K.H.; Lee, A.R.; Ahn, J.; et al. A decrease in functional microbiomes represented as Faecalibacterium affects immune homeostasis in long-term stable liver transplant patients. Gut Microbes 2022, 14, 2102885. [Google Scholar] [CrossRef] [PubMed]
- Guasch-Ferre, M.; Bhupathiraju, S.N.; Hu, F.B. Use of Metabolomics in Improving Assessment of Dietary Intake. Clin. Chem. 2018, 64, 82–98. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.A.; Rinaldi, S.; Ferrari, P.; Carayol, M.; Achaintre, D.; Scalbert, A.; Cross, A.J.; Gunter, M.J.; Fensom, G.K.; Appleby, P.N.; et al. Metabolic profiles of male meat eaters, fish eaters, vegetarians, and vegans from the EPIC-Oxford cohort. Am. J. Clin. Nutr. 2015, 102, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Wegermann, K.; Howe, C.; Henao, R.; Wang, Y.; Guy, C.D.; Abdelmalek, M.F.; Diehl, A.M.; Moylan, C.A. Serum Bile Acid, Vitamin E, and Serotonin Metabolites Are Associated With Future Liver-Related Events in Nonalcoholic Fatty Liver Disease. Hepatol. Commun. 2021, 5, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Gao, X.Q.; Shen, N.; He, J.; Fan, X.; Chen, K.; Lin, X.H.; Li, H.M.; Tian, F.S.; Li, H. A targeted metabolomic profiling of plasma acylcarnitines in nonalcoholic fatty liver disease. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7433–7441. [Google Scholar]
- Mir, I.H.; Thirunavukkarasu, C. The relevance of acid sphingomyelinase as a potential target for therapeutic intervention in hepatic disorders: Current scenario and anticipated trends. Arch. Toxicol. 2023, 97, 2069–2087. [Google Scholar] [CrossRef]
- Piras, C.; Noto, A.; Ibba, L.; Deidda, M.; Fanos, V.; Muntoni, S.; Leoni, V.P.; Atzori, L. Contribution of Metabolomics to the Understanding of NAFLD and NASH Syndromes: A Systematic Review. Metabolites 2021, 11, 694. [Google Scholar] [CrossRef]
- Lovric, A.; Graner, M.; Bjornson, E.; Arif, M.; Benfeitas, R.; Nyman, K.; Stahlman, M.; Pentikainen, M.O.; Lundbom, J.; Hakkarainen, A.; et al. Characterization of different fat depots in NAFLD using inflammation-associated proteome, lipidome and metabolome. Sci. Rep. 2018, 8, 14200. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.H.; Yoon, J.E.; Ko, M.S.; Leem, J.; Yun, J.Y.; Hong, C.H.; Cho, Y.K.; Lee, S.E.; Jang, J.E.; Baek, J.Y.; et al. Sphingomyelin synthase 1 mediates hepatocyte pyroptosis to trigger non-alcoholic steatohepatitis. Gut 2021, 70, 1954–1964. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Orešič, M.; Leivonen, M.; Gopalacharyulu, P.; Hyysalo, J.; Arola, J.; Verrijken, A.; Francque, S.; Van Gaal, L.; Hyötyläinen, T.; et al. Noninvasive Detection of Nonalcoholic Steatohepatitis Using Clinical Markers and Circulating Levels of Lipids and Metabolites. Clin. Gastroenterol. Hepatol. 2016, 14, 1463–1472.e1466. [Google Scholar] [CrossRef] [PubMed]
- Teunis, C.; Nieuwdorp, M.; Hanssen, N. Interactions between Tryptophan Metabolism, the Gut Microbiome and the Immune System as Potential Drivers of Non-Alcoholic Fatty Liver Disease (NAFLD) and Metabolic Diseases. Metabolites 2022, 12, 514. [Google Scholar] [CrossRef]
- Merola, J.; Emond, J.C.; Levitsky, J. Novel Noninvasive Biomarkers in Liver Transplantation: A Tool on the Doorstep of Clinical Utilization. Transplantation 2023, 107, 2120–2125. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | MASH (n = 10) | TCMR (n = 18) | Biliary Obstruction (n = 27) | p-Value |
|---|---|---|---|---|
| Recipient age at LT (years) | 54.5 [47,67] | 52 [46,60] | 59 [53,67] | 0.246 a |
| Sex | 0.489 b | |||
| M | 5 (50%) | 13 (72.2%) | 18 (66.7%) | |
| F | 5 (50%) | 5 (27.8%) | 9 (33.3%) | |
| Laboratory readings (U/L) | ||||
| ALT | 77 [45,165] | 115 [83,305] | 55 [37,90] | 0.006 a |
| AST | 45.5 [28,132] | 76 [47,198] | 36 [25,43] | 0.001 a |
| ALP | 168.0 [104,684] | 205 [122,329] | 241 [147,408] | 0.478 a |
| Creatinine | 98.5 [71,140] | 80 [71,107] | 100 [81,131] | 0.546 a |
| Hgb | 123.5 [85,146] | 112 [105,120] | 104 [94,115] | 0.161 a |
| Indication for transplant | 0.837 b | |||
| ArLD | 4 (40%) | 4 (22.2%) | 9 (33.4%) | |
| MASH | 3 (30%) | 2 (11.1%) | 4 (14.8%) | |
| HBV | 1(10%) | 2 (11.1%) | 1 (3.7%) | |
| HCV | - | 3 (16.7%) | 3 (11.1%) | |
| PBC | - | 2 (11.1%) | 1 (3.7%) | |
| PSC | - | 1 (5.6%) | 2 (7.4%) | |
| Autoimmune hepatitis | - | - | 3 (11.1%) | |
| Other | 2 (20%) | 4 (22.2%) | 4 (14.8%) |
| Group Comparison | Pathway Name | Altered Metabolites from Input List | FDR |
|---|---|---|---|
| MASH vs. TCMR | Alanine, aspartate and glutamate metabolism | L-Asparagine; Citrate; Fumarate; Succinate; 2-Oxoglutarate | 1.09 × 10−3 |
| Citrate cycle (TCA cycle) | 2-Oxoglutarate; Succinate; Citrate; Fumarate | 2.91 × 10−2 | |
| Arginine biosynthesis | L-Citrulline; Oxoglutarate; Fumarate | 1.46 × 10−2 | |
| MASH vs. Biliary | Aminoacyl-tRNA biosynthesis | L-Asparagine; L-Phenylalanine; Glycine; L-Aspartate; L-Valine; L-Alanine; Isoleucine; L-Leucine; L-Tryptophan | 1.35 × 10−6 |
| Valine, leucine and isoleucine biosynthesis | L-Leucine; L-Isoleucine; L-Valine | 2.99 × 10−3 | |
| Alanine, aspartate and glutamate metabolism | L-Aspartate; L-Asparagine; L-Alanine; Citrate; Succinate | 6.15 × 10−3 | |
| Biliary vs. TCMR | Butanoate metabolism | (R)-3-Hydroxybutanoate, Butanoic acid; 2-Oxoglutarate; Succinate | 8.13 × 10−4 |
| Alanine, aspartate and glutamate metabolism | L-Aspartate; L-Alanine; Succinate; 2-Oxoglutarate | 3.75 × 10−3 | |
| Arginine biosynthesis | L-Aspartate; L-Citrulline; 2-Oxoglutarate | 6.45 × 10−3 |
| Metabolite | Biliary | MASH | TCMR |
|---|---|---|---|
| Decanoylcarnitine (C10) | 0.811 | 0.901 | 0.901 |
| Citric acid | 0.832 | 0.868 | 0.868 |
| Succinic acid | 0.811 | 0.835 | 0.835 |
| Phenylalanine | 0.779 | 0.846 | 0.846 |
| Serine | 0.686 | 0.868 | 0.868 |
| Dodecanoylcarnitine (C12) | 0.739 | 0.835 | 0.835 |
| Serotonin | 0.812 | 0.780 | 0.812 |
| Methylglutarylcarnitine | 0.754 | 0.824 | 0.824 |
| Hydroxysphingomyeline C22:1 | 0.832 | 0.832 | 0.72 |
| alpha-Aminoadipic acid | 0.725 | 0.824 | 0.824 |
| Dodecenoylcarnitine (C12:1) | 0.700 | 0.830 | 0.83 |
| Hexadecenoylcarnitine | 0.732 | 0.808 | 0.808 |
| Octanoylcarnitine | 0.736 | 0.802 | 0.802 |
| Alanine | 0.779 | 0.779 | 0.737 |
| alpha-Ketoglutaric acid | 0.710 | 0.786 | 0.786 |
| Fumaric acid | 0.689 | 0.791 | 0.791 |
| Putrescine | 0.775 | 0.775 | 0.72 |
| Leucine | 0.786 | 0.786 | 0.687 |
| Sarcosine | 0.785 | 0.686 | 0.785 |
| Kynurenine | 0.689 | 0.775 | 0.775 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baciu, C.; Ghosh, S.; Naimimohasses, S.; Rahmani, A.; Pasini, E.; Naghibzadeh, M.; Azhie, A.; Bhat, M. Harnessing Metabolites as Serum Biomarkers for Liver Graft Pathology Prediction Using Machine Learning. Metabolites 2024, 14, 254. https://doi.org/10.3390/metabo14050254
Baciu C, Ghosh S, Naimimohasses S, Rahmani A, Pasini E, Naghibzadeh M, Azhie A, Bhat M. Harnessing Metabolites as Serum Biomarkers for Liver Graft Pathology Prediction Using Machine Learning. Metabolites. 2024; 14(5):254. https://doi.org/10.3390/metabo14050254
Chicago/Turabian StyleBaciu, Cristina, Soumita Ghosh, Sara Naimimohasses, Arya Rahmani, Elisa Pasini, Maryam Naghibzadeh, Amirhossein Azhie, and Mamatha Bhat. 2024. "Harnessing Metabolites as Serum Biomarkers for Liver Graft Pathology Prediction Using Machine Learning" Metabolites 14, no. 5: 254. https://doi.org/10.3390/metabo14050254