
Lysophosphatidylinositol Signalling and Metabolic Diseases



1
Inositide Signalling Group, Centre for Cell Biology and Cutaneous Research, Blizard Institute, Queen Mary University of London, 4 Newark Street, London E1 2AT, UK
2
Department of Basic Medical Science for Nursing, Faculty of Nursing, IIUM, Bandar Indera Mahkota, Kuantan Pahang 25200, Malaysia
3
Metabolic Signalling Group, School of Biomedical Sciences, Curtin Health Innovation Research Institute Biosciences, Curtin University, Perth 6102, Australia
*
Author to whom correspondence should be addressed.
Metabolites 2016, 6(1), 6; https://doi.org/10.3390/metabo6010006
Submission received: 11 December 2015
/
Revised: 7 January 2016
/
Accepted: 11 January 2016
/
Published: 15 January 2016
(This article belongs to the Special Issue Metabolomic Studies in Metabolic Diseases)
{kind=link}
Abstract
:Metabolism is a chemical process used by cells to transform food-derived nutrients, such as proteins, carbohydrates and fats, into chemical and thermal energy. Whenever an alteration of this process occurs, the chemical balance within the cells is impaired and this can affect their growth and response to the environment, leading to the development of a metabolic disease. Metabolic syndrome, a cluster of several metabolic risk factors such as abdominal obesity, insulin resistance, high cholesterol and high blood pressure, and atherogenic dyslipidaemia, is increasingly common in modern society. Metabolic syndrome, as well as other diseases, such as diabetes, obesity, hyperlipidaemia and hypertension, are associated with abnormal lipid metabolism. Cellular lipids are the major component of cell membranes; they represent also a valuable source of energy and therefore play a crucial role for both cellular and physiological energy homeostasis. In this review, we will focus on the physiological and pathophysiological roles of the lysophospholipid mediator lysophosphatidylinositol (LPI) and its receptor G-protein coupled receptor 55 (GPR55) in metabolic diseases. LPI is a bioactive lipid generated by phospholipase A (PLA) family of lipases which is believed to play an important role in several diseases. Indeed LPI can affect various functions such as cell growth, differentiation and motility in a number of cell-types. Recently published data suggest that LPI plays an important role in different physiological and pathological contexts, including a role in metabolism and glucose homeostasis.
1. Introduction
Metabolic diseases, such as diabetes, cardiovascular disease and obesity, can be a result of genetics factors, a deficiency in a certain hormone or enzyme, an unbalanced diet or very often a combination of these factors. Obesity, diabetes and cardiovascular disease are strictly interlinked, since obesity is known to increase the risk of cardiovascular disease in adults and to be highly associated with insulin resistance in normo-glycaemic individuals and in patients with type 2 diabetes [1]. Diabetes is the most common metabolic disease and currently one of the major health problems worldwide and it is growing at a very fast rate, mainly because of its strong link with obesity. Diabetes is associated with several complications, including micro and macrovascular complications that are mainly caused by poorly controlled blood glucose levels and that ultimately result in reduced life expectancy. Diabetes can be classified in type 1, an auto-immune disease in which the insulin-producing β cells in the pancreas are destroyed, and type 2, which can be the consequence of acquired environmental agents, as well as genetic factors. Transition from normal glucose tolerance to impaired glucose tolerance and ultimately to type 2 diabetes is accompanied by a decrease in both insulin sensitivity and pancreatic β cell function. Metabolic disorders are often associated to an abnormal increase in dietary fats and the caloric surplus leads to impaired lipid metabolism and fat accumulation which is a fundamental step in the progression of metabolic disorders [2]. Lipids are the major components of cellular membranes and a source of energy and play a crucial role in cellular and physiological energy homeostasis. Lipids include phospholipids and neutral lipids, primarily triacylglycerols and sterol esters. Phospholipids are key components of the membrane structure and major substrates for enzymes, such as phospholipase A (PLA) phospholipase C (PLC), phospholipase D (PLD) and lysophospholipase D. Lysophospholipids are the product of the activity of PLA2 on phospholipids and are well known to act as extracellular signals. Lysophospholipids acting as signalling mediator include lysophosphatidic acid, sphingosine-1-phosphate, lysophosphatidylserine and LPI. In the past two decades lysophospholipids have emerged as second-messenger molecules regulating intracellular signalling pathways that are involved in many physiological and pathological functions, such as inflammation, angiogenesis, nervous system regulation, atherosclerosis and tumorigenesis [3,4,5]. This review will focus on the production and signalling pathways downstream LPI and the emerging evidence suggesting its role in metabolism, with specific interest in LPI, and its main receptor GPR55 involvement in metabolic diseases.
2. Lysophosphatidylinositol
LPI is a bioactive lipid generated by the PLA family of lipases which is believed to play an important role in several physiological and pathological processes [6]. In the past thirty year, LPI has been shown to affect various cellular actions such as cell growth, differentiation and motility, in a wide range of cell-types, including cancer cells, endothelial cells and nervous cells. LPI can be the product of the PLA family of lipases comprising PLA1 and PLA2. PLA1 and PLA2 enzymes remove fatty acids from the sn-1 and sn-2 position of glycerophospholipids respectively, generating 2-acyl-lysophospholipids and 1-acyl-lysophospholipids. There are few data on the different biological activity of 2-acyl compared to 1-acyl-LPI, as well as the activity of distinct LPI species containing different fatty acid chains [6]. Recently published data suggest that LPI plays an important role in different physio pathological circumstances, including a role in metabolism and glucose homeostasis [7,8]. The first evidence showing a possible physiological role for LPI came with two papers published in the mid-1980s showing a stimulatory effect of LPI in the release of insulin by pancreatic islets [9,10]. Subsequently, our work clearly identified LPI as a potent mitogenic factor [11,12,13]. Interestingly, our early work identified a synergistic effect of LPI and insulin in promoting cell proliferation on thyroid cells FRTL5 [11]. Importantly, a key role in metabolically active tissues has also been identified [14] and evidence further suggests that LPI is involved in the regulation of fat deposition. Specifically, it has been observed that plasma levels of LPI were significantly higher in obese female patients and positively correlated with weight and percentage body fat [7]. Furthermore, in visceral adipose tissue explants and primary differentiated visceral adipocytes, LPI induces the expression of genes involved in fat deposition.
The LPI biological activities can be divided in non-receptor and receptor-mediated [6]. Recently, the orphan receptor GPR55 has been proposed as potential LPI receptor [15,16]. GPR55 is widely expressed in the brain, especially in the cerebellum as well as in the jejunum and ileum. Importantly, recent studies have demonstrated that GPR55 is also expressed in the endocrine pancreas and in pancreatic β cells where it is involved in stimulus-secretion coupling of insulin secretion, suggesting a role in glucose homeostasis [17]. This would be consistent with the reported effect of LPI on the stimulation of insulin secretion, which has been partly ascribed to its ability to induce intracellular calcium release in rat and mouse pancreatic islets [9,10]. It has been suggested that the LPI-induced insulin release could be correlated with its ability to stimulate the calcium-dependent exocytosis activity, as it has been very recently shown in PC12 neuroendocrine cells [18]. Indeed, exocytosis in PC12 cells is Ca2+-dependent, and therefore the increase in intracellular Ca2+ induced by LPI could trigger exocytosis. It is currently unknown whether this LPI action is mediated by GPR55 or other receptors, or by its physical properties. However, it is well-known that LPI effect on calcium is mediated by GPR55 in different cells [6]. In addition to acting on GPR55, LPI could also have non-receptor mediated effects. For instance, it has been recently demonstrated a receptor-independent effect of LPI on the Ca2+-activated K+ channels single-channel activity of endothelial cells [19]. Similarly, it has been shown that LPI activates TREK channels such as bTREK-1 and bKv1.4 K+ currents [20]. Furthermore, it has been shown that LPI activates TRPV2 channels in prostate cancer cells [21].
3. GPR55
3.1. Structure and Distribution
The human GPR55 is composed of 319 amino acids and has an expected molecular weight of 37 kDa and is encoded by the GPR55 gene located on chromosome 2q27. It was first cloned in 1999 and belongs to the purine cluster of rhodopsin family receptors [22]. It displays sequence similarity to cannabinoid receptors CB1 (13%) and CB2 (14%). Furthermore, it has homologies with other GPCRs such as GPR23 (30%), P2Y5 (29%), GPR35 (27%) and chemokine receptor CCR4 (23%). In human, GPR55 mRNA transcript have been found in the brain regions of caudate and putamen [22], adipose tissue, testis, myometrium, tonsil, adenoid and spleen [23]. In mouse, GPR55 mRNA expression was identified in adrenal, spleen, jejunum, ileum, frontal cortex, hippocampus, cerebellum, dorsal striatum and hypothalamus [17,24]. In addition, diverse range of human cancer cell lines are also expressing GPR55 including ovary, prostate [25], breast [26,27], skin [28], as well as cervix, liver, blood and pancreas [26]. Despite being listed as an orphan receptor in the IUPHAR database, several endogenous and pharmacological ligands have been reported to activate GPR55 [24]. Initially, GPR55 was considered as an atypical cannabinoid receptor (CB) due to its activation shown by ∆9-tetrahydrocannabinol, abnormal cannabidiol, and its synthetic derivative, O-1602, as well as by endogenous cannabinoids anandamide, palmitoyl ethanolamine and oleoyl ethanolamine [24]. Interestingly, another paper published in the same year by Oka et al. [15], has identified a lysophospholipid, LPI, as the endogenous ligand for GPR55. The potent LPI agonist activity toward GPR55 was also demonstrated by other studies [29,30,31,32]. Recently, a nomenclature review for lysophospholipids receptors considered GPR55 as a provisional LPI receptor with the receptor name LPI1 and gene names LPIR1/Lpir1 for human and non-human genes, respectively [33].
3.2. GPR55 Signalling
The pharmacology of GPR55 appears to be much entangled. It is unclear whether this receptor is another member of the CB family or not, due to conflicting data about its activation by endocannabinoids and non-cannabinoid ligands [34]. The sensitivity of GPR55 to endocannabinoids such as anandamide [24] and not to other endocannabinoids [30] makes it a good candidate. On the other hand, its phylogenetically distinction from traditional CB receptor has prevented its classification as a novel CB receptor. However, the weight of evidence point to LPI as the most promising endogenous ligand for GPR55 [15,29,35,36]. The selectivity of LPI as the GPR55 ligand was studied by Kotsikorou et al. [37]. They discovered that GPR55 accommodates LPI in the horizontal binding pocket within the transmembrane domain 2 of its polar head group. It has now been demonstrated that GPR55 is associated to Gα12/13 and Gαq subunits and that it can activate several signalling pathways. Upon LPI stimulation of human osteosarcoma cell line U20S, Gαq subunit is able to stimulate PLC activity that induces Ca2+ release from the endoplasmic reticulum, activating different PKC isoforms. PKCs catalyse the phosphorylation of different intracellular proteins, such as MAPK and related signalling pathways. GPR55 activation by LPI stimulation was shown to activate ERK1/2 and to be able to activate two transcription factors, such as the cAMP response element-binding protein (CREB) and the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), which can then regulate gene transcription [38]. Moreover, upon LPI stimulation, Gα12/13 activates the RhoA/ROCK signalling pathway. GPR55 activation of RhoA/ROCK signalling pathway regulates PLC, actin cytoskeleton and p38/Activating transcription factor 2 (ATF2) activities [38]. Finally, prolonged oscillatory Ca2+ release from intracellular stores leads to the activation and translocation of the nuclear factor of activated T cells (NFAT), which can then regulate DNA transcription and gene expression [29]. Both MAPK and RhoA/ROCK signalling pathways are able to control and regulate a plethora of cellular functions, such as cell proliferation, cell division, apoptosis, cell differentiation and actin cytoskeleton remodelling. Moreover, MAPKs and RhoA/ROCK signalling pathways were found to be deregulated in various cancers and several studies suggest the use of inhibitors of these pathways to counteract cancer cell growth and metastatic development [39].
4. LPI/GPR55 Signalling and Physiological Function
4.1. Metabolism
There are increasing reports that GPR55 may play a role in the modulation of energy homeostasis. Initial indication of the possible involvement of GPR55 in metabolism, was provided by the evidence that LPI induced insulin release from pancreatic islets via mobilisation of Ca2+ ion [9,10]. Consistent with this, a study on isolated islet has revealed high GPR55 mRNA expression and its activation with pharmacological agonist O-1602 showed an increase in Ca2+ release and insulin secretion stimulated by glucose [17]. The in vivo data from the latter study also showed that GPR55 activation increases glucose tolerance and plasma insulin levels. Furthermore, similar results were obtained by another study on isolated islet stimulated with various GPR55 agonists. It was shown that GPR55 agonists such as Abn-CBD PEA, OEA, O-1602, and AM-251, significantly stimulate insulin secretion and later confirmed to lower blood glucose in in vivo studies [40].
4.2. Endothelial Cells and Vasculature
Increasing evidence supports an endothelial site of action for LPI and suggests a possible role for LPI/GPR55 axis in vasculature. For instance, studies suggested that GPR55 relaxes rat mesenteric resistance artery when activated by LPI and not by atypical cannabinoids. LPI, indeed, induces calcium release in rat mesenteric artery endothelial cells with vasodilator effects [41,42]. LPI is an endothelium-dependent vasodilator in rat small mesenteric artery and a hypotensive agent. Signalling pathways activated by LPI in endothelial cells include PLC-IP3 and ROCK-RhoA to elevate intracellular Ca2+. Studies using the selective antagonist CID16020046 uncovered a potential role for G protein-coupled receptor 55 in platelet and endothelial cell function [43]. It has been shown that CID16020046 reduced LPI-induced wound healing in primary human lung microvascular endothelial cells and impeded LPI-inhibited platelet aggregation, suggesting a novel role for LPI/GPR55 axis in platelet and endothelial cell function [43].
4.3. Gastrointestinal Functions
The interest on bioactive phospholipids present in the gastrointestinal mucosa has rapidly grown during the past decade. Recently, it has become increasingly evident that lysophospholipids can influence gastrointestinal functions and ultimately hormones and peptides release from the gut. Indeed, it has been shown that food-derived lipids can directly modulate gastrointestinal functions. In addition, it has been observed that the digestive tract itself is able to release different types of bioactive lipids, together with polypeptide hormones, into the lumen. GPR55 has been found to be largely expressed in the gastrointestinal tract and therefore several studies have focused their attention on the potential pathophysiological role of GPR55 in gastrointestinal functions [44]. It has been suggested that GPR55 may have a role in intestinal inflammation [44]. Indeed, the GPR55 antagonist CID16020046 has been shown to have anti-inflammatory activity in mice subjected to different models of intestinal inflammation [45]. It has been recently shown that postprandial plasma phospholipids in men are influenced by the source of dietary fat [46]. In particular, postprandial elevation of LPI has been found after dairy and soy meals with important implications for LPI effect on metabolic processes. Interestingly, it has been recently demonstrated that LPI can also activate GPR119 in RH7777 rat hepatoma cells stably expressing human GPR119 [47]. GPR119 is expressed predominantly in the pancreas and gastrointestinal tract and its activation has been shown to cause a reduction in food intake and body weight gain in animal models [48]. In addition, it has been shown that the activation of GPR119 can induce incretins release in the intestine. However, GPR119 appears to be activated by other lipids, including lysophospholipids, with higher affinity compared to LPI and therefore further studies are required in order to assess if, in particular conditions, GPR119 may mediate LPI effects in vivo.
4.4. Central and Peripheral Nervous System
GPR55 mRNA is expressed in microglial from primary mouse, BV-2 cell line [49] and neurons [50]. It has been suggested that LPI activated GPR55 is able to induce exocytosis and catecholamine release from the neuroendocrine pheochromocytoma-12 cells [18,35] Furthermore, the activation of dorsal root ganglion (DRG) neurons by GPR55 increases intracellular calcium and inhibits potassium current through M-type potassium channel [30]. This neuronal excitability of GPR55, particularly on large diameter DRG neurons, suggested its pro-nociceptive role [30]. The hypothesis that GPR55 activation inhibits neurogenic contractions in the gut has been recently investigated. By testing the inhibitory effect of the atypical cannabinoid O-1602, a GPR55 agonist, in mouse colon, it has been found that the activation of GPR55 leads to inhibition of neurogenic contractions in the gut [51]. Noteworthy, chronic intake of high fat and obesogenic diet has resulted in a significant reduction in the response of intestinal afferents to satiety mediators such as cholecystokinin and 5HT [52]. The authors of the latter study suggested that the impairment of vagal afferent function involves decreased satiety signalling from gastrointestinal organs to the brain [52]. Consistently with this, diminished vagal nerve signalling in obesity could lead to enhanced inflammation and metabolic complications [53].
4.5. Nociception and Inflammation
Several studies have implicated GPR55 in neuropathic and inflammatory pain. In two models of hyperalgesia, it has been reported that inflammatory mechanical hyperalgesia induced by administration of Freund’s complete adjuvant was completely absent in GPR55 knockout mice [54]. A similar response is obtained when GPR55−/− mice undergo partial nerve ligation [55], an established pre-clinical model for neuropathic hyperalgesia. Moreover, the authors demonstrated that the levels of the anti-inflammatory cytokines Interleukin-4, Interleukin-10, Interferon γ and Granulocyte Macrophage-Colony Stimulating Factor, were higher in GPR55−/− mice compared to control mice 14 days post FCA-injection [54]. A recent study may provide an explanation for the absence of hyperalgesia in GPR55−/− mice [50]. The authors identified the GPR55 involvement in central pain processing and unravelled a novel Ca2+-mediated mechanism through activation of periaqueductal grey neurons by GPR55 [50]. Consistently, new data suggested a possible role of GPR55 in the inflammatory system. Indeed, GPR55 is overexpressed in monocytes and NK cells and its activation using a synthetic agonist (O-1602) is able to stimulate IL-2, IL-12, and TNFα. Moreover, the activation also increases CD69 activation marker expression, granzyme B, CD107-dependent cytotoxicity and IFN-γ, suggesting a pro-inflammatory role of GPR55 in the innate immunity [56].
4.6. Bone
Whyte and colleagues have found that GPR55 agonist O-1602, affects the regulation of bone mass (osteoclastogenesis), stimulating osteoclast resorption, suppressing formation in vitro and reducing bone resorption in vivo in mice [57]. They also reported an increased GPR55 expression in osteoclasts, compared to macrophages precursor. Consistently with this, activation of CB2 and the inhibition of GPR55 by cannabidiol promotes migration of mesenchymal stem cells in the regenerative process of bone healing [58].
5. LPI/GPR55 Axis and Disease
5.1. Obesity and Type-2 Diabetes
The imbalance of energy homeostasis could lead to metabolic disorders such as obesity and type 2 diabetes. It was reported that a polymorphism in Val195 allele of GPR55 gene has a link with anorexia nervosa, a disorder characterised by low body weight and obsessive fear of weight gain [59]. This report was later confirmed by a study in humans that discovered higher levels of GPR55 expression in visceral adipose tissue and sub-cutaneous tissue correlated with an increase in body weight and fat percentage [7]. In addition, another study also showed that O-1602 influences obesity by acutely stimulate food intake and chronically increases adiposity in Sprague-Dawley rats [60]. Imbernon et al. (2014) compared the expression of GPR55 and CB1 receptors expression in rat white adipose tissue and discovered increased levels of the GPR55 expression after fasting and recovered after leptin treatment [8]. However, GPR55 expression showed an increase throughout 90 days of the experiment and only reduced during the gestation period. Recently, Meadows et al. (2015), reported significant increase in fat mass and insulin resistance in GPR55−/− mice [61]. They also discover that locomotor and voluntary physical activity substantially reduced with intact muscle function, thus suggesting that the selective decrease in physical activity is caused by GPR55 ablation and not due to changing of feeding behaviour as CB1. However, the fact that GPR55 is expressed in enteric neurons could suggest that GPR55 has a role in gastrointestinal functions, such as secretion and motility [62]. In addition, LPI has been found to counteract the symptoms of diabetes such as high blood glucose, lower body weight, increase amplitude of slow wave, and to improve gastric emptying in streptozotocin induced diabetes in rat. The authors also discover that GPR55 mRNA is up regulated in the STZ-induced diabetic condition. Furthermore, treatment of the diabetic rats with LPI induces increased levels of motilin, gastrin, vasoactive intestinal peptide and somatostatin, indicating the role of GPR55 in controlling movement in the gut [63]. Recently, the functional role of GPR55 in diabetes-induced nephropathy was studied in rat proximal tubules. It was shown that GPR55 mRNA and protein expression in proximal tubule has increased in response to high glucose in presence or absence of high albumin [64]. The authors suggest that GPR55 may play a functional role in the tubules that is altered in response to pathophysiological conditions. A potential role for LPI in type-1 diabetes pathogenesis has been recently suggested. Indeed, a lipidomic characterization of a genetically modified mouse model of the early stages of human type-1 diabetes revealed an increase in LPI in the pre-type 1 diabetic mouse compared to control mice. These findings open an intriguing scenario for LPI suggesting that LPI may play a role in autoimmune diseases and inflammation [65].
5.2. Cardiovascular Disease
A recent study has suggested that GPR55 is able to control the adrenergic signalling pathway in the heart and to have a potential role in the pathogenesis of heart failure [66]. Interestingly, the cardiac function of young GPR55 knockout mice was the same as control mice, whereas mature GPR55 knockout mice were characterised by systolic dysfunction and ventricular remodelling compared to control mice [66]. Recently, LPI has been found to increase significantly following asphyxia-induced cardiac arrest using a rat model [67]. Interestingly, in the same rat model when cardiac arrest was followed by cardiopulmonary resuscitation the LPI levels decrease drastically. These data suggest that LPI levels could be regulated by cellular conditions associated with ischemia and reperfusion and have possible important implications for improving atherosclerosis. Interestingly, measurements of the levels of lysophospholipids in 141 patients undergoing coronary angiography revealed that lysophospholipids, including LPI, were elevated in acute coronary syndrome patients [68]. Furthermore, a study on human macrophages and foam cells revealed that activation of GPR55 by O-1602 exacerbates oxidative low-density lipoprotein-induced lipid accumulation and inflammatory responses, reducing cholesterol efflux [69].
5.3. Cancer
Several studies highlighted the role of GPR55 in several types of cancer and we have recently reviewed the role of the LPI/GPR55 axis in cancer [70]. Our group demonstrated that GPR55 has a key role in cancer by defining a new autocrine loop that involves GPR55, LPI and the ABC transporter, ABCC1, in prostate and ovarian cancer cell lines [71]. Another report revealed that this receptor plays an important role also in the modulation of migration and invasion of human breast cancer cell lines [27]. Furthermore, another study demonstrated that downregulation of GPR55 in a xenograft mice model of glioblastoma reduces tumour growth [26] and that GPR55 knockout mice are more resistant to skin carcinogenesis [28].
6. Conclusions
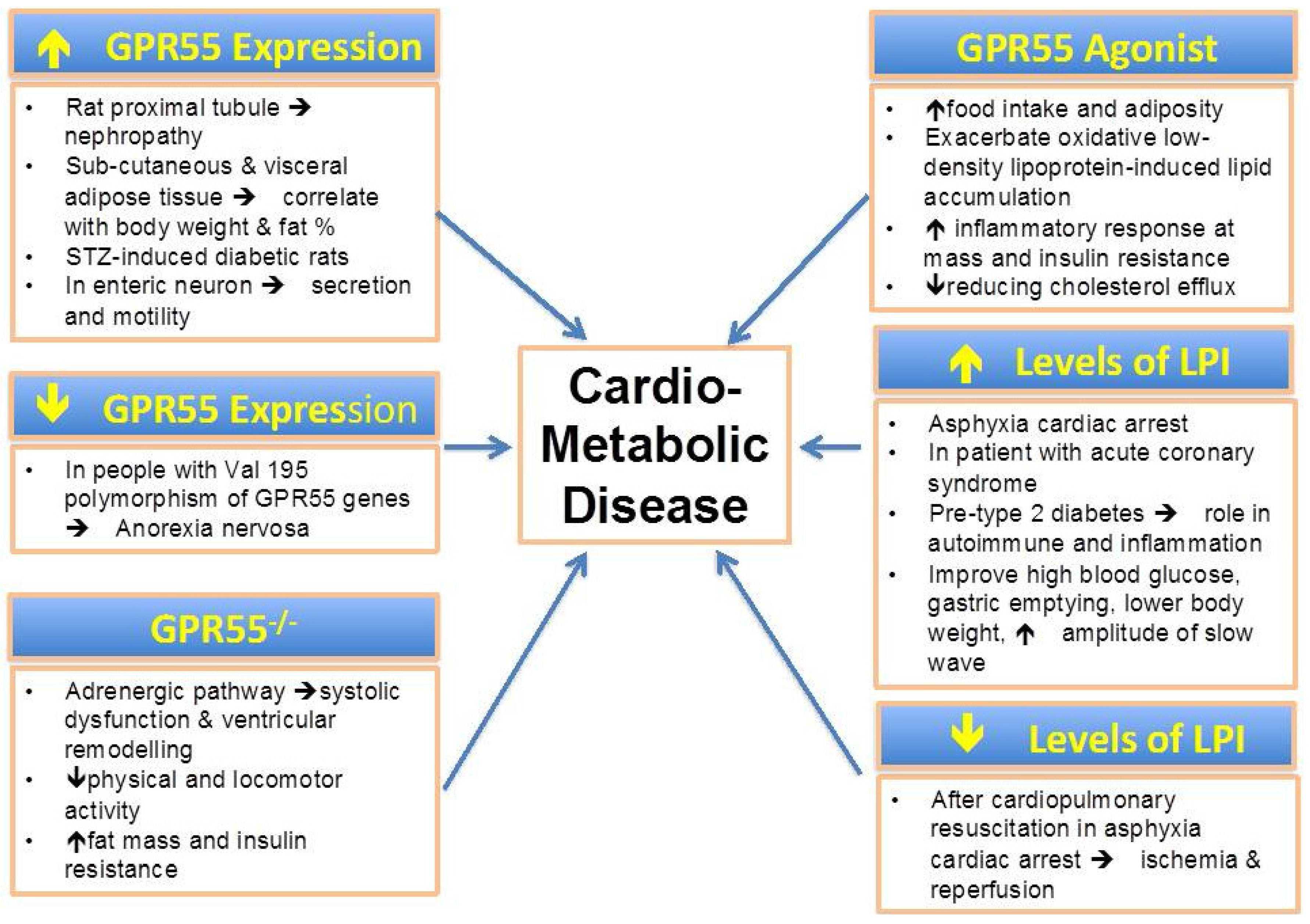
Increasing evidence suggest that LPI play a key role in several metabolic functions and possibly in metabolic disorders (Figure 1). In parallel, GPR55, the main LPI receptor, has been proposed as a metabolic regulator [72,73]. The LPI/GPR55 axis has been shown to be positively associated with obesity in human. Altered levels of LPI have been found in obesity and diabetes. In addition, the LPI/GPR55 system is also involved in inflammation and cancer. Further mechanistic studies are required to elucidate the potential role of LPI in the pathogenesis of metabolic disorders.
Figure 1.
The pathophysiological relevance of GPR55 expression, levels of LPI and potential role of its agonist in cardio-metabolic diseases.
Figure 1.
The pathophysiological relevance of GPR55 expression, levels of LPI and potential role of its agonist in cardio-metabolic diseases.

Author Contributions
Syamsul Arifin: Idea, concept, collection of data, review, paper writing, and paper revision; Marco Falasca: Idea, Collection of data, paper revision, and editing, concept, and review of paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eckel, R.H.; Alberti, K.; Grundy, S.M.; Zimmet, P.Z. The Metabolic Syndrome. Lancet 2010, 375, 181–183. [Google Scholar] [CrossRef]
- Lee, C.-H.; Olson, P.; Evans, R.M. Minireview: Lipid Metabolism, Metabolic Diseases, and Peroxisome Proliferator-Activated Receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.J.W.; Chun, J. Lysophospholipids and Their Receptors in the Central Nervous System. Biochim. Biophys. Acta 2013, 1831, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Tigyi, G. New Trends in Lysophospholipid Research. Biochim. Biophys. Acta 2013. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y.; Mizuno, H.; Chun, J. Lysophospholipid Receptors in Drug Discovery. Exp. Cell Res. 2015, 333, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Piñeiro, R.; Falasca, M. Lysophosphatidylinositol Signalling: New Wine from an Old Bottle. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2012, 1821, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Catalán, V.; Whyte, L.; Díaz-Arteaga, A.; Vázquez-Martínez, R.; Rotellar, F.; Guzmán, R.; Gómez-Ambrosi, J.; Pulido, M.R.; Russell, W.R.; et al. The L-α-Lysophosphatidylinositol/GPR55 System and its Potential Role in Human Obesity. Diabetes 2012, 61, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Imbernon, M.; Whyte, L.; Diaz-Arteaga, A.; Russell, W.R.; Moreno, N.R.; Vazquez, M.J.; Gonzalez, C.R.; Díaz-Ruiz, A.; Lopez, M.; Malagón, M.M.; et al. Regulation of GPR55 in Rat White Adipose Tissue and Serum LPI by Nutritional Status, Gestation, Gender and Pituitary Factors. Mol. Cell. Endocrinol. 2014, 383, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Metz, S.A. Lysophosphatidylinositol, but not Lysophosphatidic Acid Stimulates Insulin Release. Biochem. Biophys. Res. Commun. 1986, 138, 720–727. [Google Scholar] [CrossRef]
- Metz, S. Mobilization of Cellular Ca2+ by Lysophospholipids in Rat Islets of Langerhans. Biochim. Biophys. Acta 1988, 968, 239–252. [Google Scholar] [CrossRef]
- Falasca, M.; Corda, D. Elevated Levels and Mitogenic Activity of Lysophosphatidylinositol in K-ras-transformed Epithelial Cells. Eur. J. Biochem. 1994, 221, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Falasca, M.; Siletta, M.; Carvelli, A.; di Francesco, A.; Fusco, A.; Ramakrishna, V.; Corda, D. Signalling Pathways Involved in the Mitogenic Action of Lysophosphatidylinositol. Oncogene 1995, 10, 2113–2124. [Google Scholar] [PubMed]
- Falasca, M.; Lurisci, C.; Carvelli, A.; Sacchetti, A.; Corda, D. Release of the Mitogen Lysophosphatidylinositol from H-Ras-transformed Fibroblasts; A Possible Mechanism of Autocrine Control of Cell Proliferation. Oncogene 1998, 16, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Henstridge, C.M.; Balenga, N.A.B.; Kargl, J.; Andradas, C.; Brown, A.J.; Irving, A.; Sanchez, C.; Waldhoer, M. Minireview: Recent Developments in the Physiology and Pathology of the Lysophosphatidylinositol-sensitive Receptor GPR55. Mol. Endocrinol. 2011, 25, 1835–1848. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Nakajima, K.; Yamashita, A.; Kishimoto, S.; Sugiura, T. Identification of GPR55 as a Lysophosphatidylinositol Receptor. Biochem. Biophys. Res. Commun. 2007, 362, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.A. L-α-lysophosphatidylinositol Meets GPR55: A Deadly Relationship. Trends Pharmacol. Sci. 2011, 32, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Romero-Zerbo, S.Y.; Rafacho, A.; Díaz-Arteaga, A.; Suárez, J.; Quesada, I.; Imbernon, M.; Ross, R.A.; Dieguez, C.; Rodríguez de Fonseca, F.; Nogueiras, R.; et al. A Role for the Putative Cannabinoid Receptor GPR55 in the Islets of Langerhans. J. Endocrinol. 2011, 211, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.-T.; Yeo, J.-F.; Farooqui, A.A.; Zhang, J.; Chen, P.; Ong, W.-Y. Differential Effects of Lysophospholipids on Exocytosis in Rat PC12 Cells. J. Neural Transm. 2010, 117, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, A.I.; Malli, R.; Graier, W.F. The GPR55 Agonist Lysophosphatidylinositol Directly Activates Intermediate-conductance Ca2+-activated K+ Channels. Cardiovascular Phys. 2011, 262, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Danthi, S.; Enyeart, J.A.; Enyeart, J.J. Modulation of Native TREK-1 and Kv1.4 K+ Channels by Polyunsaturated Fatty Acids and Lysophospholipids. J. Membr. Biol. 2003, 195, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Monet, M.; Gkika, D.; Lehen’kyi, V.; Pourtier, A.; Abeele, F.V.; Bidaux, G.; Juvin, V.; Rassendren, F.; Humez, S.; Prevarsakaya, N. Lysophospholipids Stimulate Prostate Cancer Cell Migration via TRPV2 Channel Activation. Biochim. Biophys. Acta—Mol. Cell Res. 2009, 1793, 528–539. [Google Scholar] [CrossRef] [PubMed]
- Sawzdargo, M.; Nguyen, T.; Lee, D.K.; Lynch, K.R.; Cheng, R.; Heng, H.H.Q.; George, S.R.; O’Dowd, B.F. Identification and Cloning of Three Novel Human G protein-coupled receptor Genes GPR52, Psi GPR53 and GPR55: GPR55 is Extensively Expressed in Human Brain. Mol. Brain Res. 1999, 64, 193–198. [Google Scholar] [CrossRef]
- Brown, A.; Wise, A. Identification of Modulators of GPR55 Activity. U.S. Patent 10/275,200, 19 June 2003. [Google Scholar]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The Orphan Receptor GPR55 is a Novel Cannabinoid Receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Piñeiro, R.; Maffucci, T.; Falasca, M. The Putative Cannabinoid Receptor GPR55 Defines a Novel Autocrine Loop in Cancer Cell Proliferation. Oncogene 2011, 30, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Andradas, C.; Caffarel, M.M.; Pérez-Gómez, E.; Salazar, M.; Lorente, M.; Velasco, G.; Guzmán, M.; Sánchez, C. The Orphan G Protein-coupled Receptor GPR55 Promotes Cancer Cell Proliferation via ERK. Oncogene 2011, 30, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Ford, L.A.; Roelofs, A.J.; Anavi-Goffer, S.; Mowat, L.; Simpson, D.G.; Irving, A.J.; Rogers, M.J.; Rajnicek, A.M.; Ross, R.A. A Role for L-alpha-lysophosphatidylinositol and GPR55 in the Modulation of Migration, Orientation and Polarization of Human Breast Cancer Cells. Br. J. Pharmacol. 2010, 160, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Gomez, E.P.; Andradas, C.; Flores, J.; Quantanilla, M.; Pamamio, J.; Guzman, M.; Sanchez, C. The Orphan Receptor GPR55 Drives Skin Carcinogenesis and is Upregulated in Human Squamous Cell Carcinomas. Oncogene 2012, 1–9. [Google Scholar]
- Henstridge, C.M.; Balenga, N.A.B.; Ford, L.A.; Ross, R.A.; Waldhoer, M.; Irving, A.J. The GPR55 Ligand L-alpha-Lysophosphatidylinositol Promotes RhoA-dependent Ca2+ Signaling and NFAT Activation. FASEB J. 2009, 23, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.-Y.; Lu, H.-C.; Hille, B.; Mackie, K. GPR55 is a Cannabinoid Receptor that Increases Intracellular Calcium and Inhibits M Current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Toshida, T.; Maruyama, K.; Nakajima, K.; Yamashita, A.; Sugiura, T. 2-Arachidonoyl-sn-glycero-3-phosphoinositol: A Possible Natural Ligand for GPR55. J. Biochem. 2009, 145, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Waldeck-Weiermair, M.; Zoratti, C.; Osibow, K.; Balenga, N.A.B.; Goessnitzer, E.; Waldhoer, M.; Malli, R.; Graier, W.F. Integrin Clustering Enables Anandamide-induced Ca2+ Signaling in Endothelial Cells via GPR55 by Protection Against CB1-receptor-Triggered Repression. J. Cell Sci. 2008, 121, 1704–1717. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y.; Maceyka, M.; Spiegel, S.; Chun, J. Lysophospholipid Receptor Nomenclature Review: IUPHAR Review 8. Br. J. Pharmacol. 2014, 171, 3575–3594. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.H.; Marzo, V.; di Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid Receptors and Their Ligands: Beyond CB 1 and CB 2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [PubMed]
- Obara, Y.; Ueno, S.; Yanagihata, Y.; Nakahata, N. Lysophosphatidylinositol Causes Neurite Retraction via GPR55, G13 and RhoA in PC12 Cells. PLoS ONE 2011, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Oka, S.; Tanikawa, T.; Hayashi, Y.; Nemoto-Sasaki, Y.; Sugiura, T. The Actions and Metabolism of Lysophosphatidylinositol, an Endogenous Agonist for GPR55. Prostaglandins Other Lipid Mediat. 2013, 107, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Kotsikorou, E.; Madrigal, K.E.; Hurst, D.P.; Sharir, H.; Lynch, D.L.; Heynen-Genel, S.; Milan, L.B.; Chung, T.D.Y.; Seltzman, H.H.; Bai, Y.; et al. Identification of the GPR55 Agonist Binding Site Using a Novel Set of High-Potency GPR55 Selective Ligands. Biochemistry 2011, 50, 5633–5647. [Google Scholar] [CrossRef] [PubMed]
- Henstridge, C.M.; Balenga, N.A.; Schroder, R.; Kargl, J.K.; Platzer, W.; Martini, L.; Arthur, S.; Penman, J.; Whistler, J.L.; Kostenis, E.; et al. GPR55 Ligands Promote Receptor Coupling to Multiple Signalling Pathways. Br. J. Pharmacol. 2010, 160, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zheng, Y. Approaches of Targeting Rho GTPases in Cancer Drug Discovery. Expert Opin. Drug Discov. 2015, 10, 991–1010. [Google Scholar] [CrossRef] [PubMed]
- McKillop, A.M.; Moran, B.M.; Abdel-Wahab, Y.H.A.; Flatt, P.R. Evaluation of the Insulin Releasing and antihyperglycaemic Activities of GPR55 Lipid Agonists Using Clonal Beta-cells, Isolated Pancreatic Islets and Mice. Br. J. Pharmacol. 2013, 170, 978–990. [Google Scholar] [CrossRef] [PubMed]
- AlSuleimani, Y.M.; Hiley, C.R. The GPR55 Agonist Lysophosphatidylinositol Mediates Vasorelaxation of the Rat Mesenteric Resistance Artery and Induces Calcium Release in Rat Mesenteric Artery Endothelial Cells. Br. J. Pharmacol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Johns, D.G.; Behm, D.J.; Walker, D.J.; Ao, Z.; Shapland, E.M.; Daniels, D.A.; Riddick, M.; Dowell, S.; Staton, P.C.; Green, P.; et al. The Novel Endocannabinoid Receptor GPR55 is Activated by Atypical Cannabinoids but Does Not Mediate Their Vasodilator Effects. Br. J. Pharmacol. 2007, 152, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Kargl, J.; Brown, A.J.; Andersen, L.; Dorn, G.; Schicho, R.; Waldhoer, M.; Heinemann, A. A Selective Antagonist Reveals a Potential Role of G protein-coupled Receptor 55 in Platelet and Endothelial Cell Function. J. Pharmacol. Exp. Ther. 2013, 346, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Schicho, R.; Storr, M. A Potential Role for GPR55 in Gastrointestinal Functions. Curr. Opin. Pharmacol. 2012, 12, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Stančić, A.; Jandl, K.; Hasenöhrl, C.; Reichmann, F.; Marsche, G.; Schuligoi, R.; Heinemann, A.; Storr, M.; Schicho, R. The GPR55 Antagonist CID16020046 Protects Against Intestinal Inflammation. Neurogastroenterol. Motil. 2015. [Google Scholar] [CrossRef] [PubMed]
- Meikle, P.J.; Barlow, C.K.; Mellett, N.A.; Mundra, P.A.; Bonham, M.P.; Larsen, A.; Cameron-Smith, D.; Sinclair, A.; Nestel, P.J.; Wong, G. Postprandial Plasma Phospholipids in Men Are Influenced by the Source of Dietary Fat. J. Nutr. 2015, 145, 2012–2018. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Ohishi, T.; Matsui, T.; Saito, T.; Matsumoto, M.; Takasaki, J.; Matsumoto, S.-I.; Kamohara, M.; Hiyama, H.; Yoshida, S.; et al. Lysophosphatidylcholine Enhances Glucose-dependent Insulin Secretion via an Orphan G-protein-coupled Receptor. Biochem. Biophys. Res. Commun. 2005, 326, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.S.; Rosenkilde, M.M.; Holst, J.J.; Schwartz, T.W. GPR119 as a Fat Sensor. Trends Pharmacol. Sci. 2012, 33, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Pietr, M.; Kozela, E.; Levy, R.; Rimmerman, N.; Lin, Y.H.; Stella, N.; Vogel, Z.; Juknat, A. Differential Changes in GPR55 During Microglial Cell Activation. FEBS Lett. 2009, 583, 2071–2076. [Google Scholar] [CrossRef] [PubMed]
- Deliu, E.; Sperow, M.; Console-Bram, L.; Carter, R.L.; Tilley, D.G.; Kalamarides, D.J.; Kirby, L.G.; Brailoiu, G.C.; Brailoiu, E.; Benamar, K.; et al. The Lysophosphatidylinositol Receptor GPR55 Modulates Pain Perception in the Periaqueductal Gray. Mol. Pharmacol. 2015, 88, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.R.; Lichtman, A.; Dewey, W.L.; Akbarali, H.I. Evidence for the Putative Cannabinoid Receptor (GPR55)-mediated Inhibitory Effects on Intestinal Contractility in Mice. Pharmacology 2012, 90, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Daly, D.M.; Park, S.J.; Valinsky, W.C.; Beyak, M.J. Impaired Intestinal Afferent Nerve Satiety Signalling and Vagal Afferent Excitability in Diet Induced Obesity in the Mouse. J. Physiol. 2011, 589, 2857–2870. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, V.A.; Tracey, K.J. The Vagus Nerve and the Inflammatory Reflex-linking Immunity and Metabolism. Nat. Rev. Endocrinol. 2012, 8, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Staton, P.C.; Hatcher, J.P.; Walker, D.J.; Morrison, A.D.; Shapland, E.M.; Hughes, J.P.; Chong, E.; Mander, P.K.; Green, P.J.; Billinton, A.; et al. The Putative Cannabinoid Receptor GPR55 Plays a Role in Mechanical Hyperalgesia Associated with Inflammatory and Neuropathic Pain. Pain 2008, 139, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Seltzer, Z.; Dubner, R.; Shir, Y. A Novel Behavioral Model of Neuropathic Pain Disorders Produced in Rats by Partial Sciatic Nerve Injury. Pain 1990, 43, 205–218. [Google Scholar] [CrossRef]
- Muzio, M.; Bosisio, D.; Polentarutti, N.; D’amico, G.; Stoppacciaro, A.; Mancinelli, R.; van’t Veer, C.; Penton-Rol, G.; Ruco, L.P.; Allavena, P.; et al. Differential Expression and Regulation of Toll-like Receptors (TLR) in Human Leukocytes: Selective Expression of TLR3 in Dendritic Cells. J. Immunol. 2000, 164, 5998–6004. [Google Scholar] [CrossRef] [PubMed]
- Whyte, L.S.; Ryberg, E.; Sims, N.A.; Ridge, S.A.; Mackie, K.; Greasley, P.J.; Ross, R.A.; Rogers, M.J. The Putative Cannabinoid Receptor GPR55 Affects Osteoclast Function in Vitro and Bone Mass in Vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 16511–16516. [Google Scholar] [CrossRef] [PubMed]
- Schmuhl, E.; Ramer, R.; Salamon, A.; Peters, K.; Hinz, B. Increase of Mesenchymal Stem Cell Migration by Cannabidiol via Activation of p42/44 MAPK. Biochem. Pharmacol. 2014, 87, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, H.; Onaivi, E.S.; Horiuchi, Y.; Imai, K.; Komaki, G.; Ishikawa, T.; Suzuki, M.; Watanabe, Y.; Ando, T.; Higuchi, S.; et al. Functional Polymorphism in the GPR55 Gene is Associated with Anorexia Nervosa. Synapse 2011, 65, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Arteaga, A.; Vázquez, M.J.; Vazquez-Martínez, R.; Pulido, M.R.; Suarez, J.; Velásquez, D.A.; López, M.; Ross, R.A.; de Fonseca, F.R.; Bermudez-Silva, F.J.; et al. The Atypical Cannabinoid O-1602 Stimulates Food Intake and Adiposity in Rats. Diabetes. Obes. Metab. 2012, 14, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Meadows, A.; Jh, L.; Cs, W.; Wei, Q.; Pradhan, G.; Yafi, M.; Hc, L. Deletion of G-protein Coupled Receptor 55 Promotes Obesity by Reducing Physical Activity. Int J. Obes. 2015. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Fichna, J.; Schicho, R.; Saur, D.; Bashashati, M.; Mackie, K.; Li, Y.; Zimmer, A.; Göke, B.; Sharkey, K.; et al. A Role for O-1602 and G protein-Coupled Receptor GPR55 in the Control of Colonic Motility in Mice. Neuropharmacology 2013, 71, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.H.; Wei, D.D.; Wang, H.C.; Wang, B.; Bai, C.Y.; Wang, Y.Q.; Li, G.E.; Li, H.P.; Ren, X.Q. Role of Orphan G protein-coupled Receptor 55 in Diabetic Gastroparesis in Mice. Sheng Li Xue Bao 2014, 66, 332–340. [Google Scholar] [PubMed]
- Jenkin, K.; McAinch, A.J.; Zhang, Y.; Kelly, D.J.; Hryciw, D.H. Elevated Cannabinoid Receptor 1 and G protein-coupled Receptor 55 Expression in Proximal Tubule Cells and Whole Kidney Exposed to Diabetic Conditions. Clin. Exp. Pharmacol. Physiol. 2015, 42, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Overgaard, A.J.; Weir, J.M.; de Souza, D.P.; Tull, D.; Haase, C.; Meikle, P.J.; Pociot, F. Lipidomic and Metabolomic Characterization of a Genetically Modified Mouse Model of the Early Stages of Human Type 1 Diabetes Pathogenesis. Metabolomics 2015. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.K.; Hector, E.E.; Andréasson, A.-C.; Jönsson-Rylander, A.-C.; Wainwright, C.L. GPR55 Deletion in Mice Leads to Age-Related Ventricular Dysfunction and Impaired Adrenoceptor-mediated Inotropic Responses. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lampe, J.; Yin, T.; Shinozaki, K.; Becker, L. Phospholipid Alterations in the Brain and Heart in a Rat Model of Asphyxia-Induced Cardiac Arrest and Cardiopulmonary Bypass Resuscitation. Mol. Cell. Biochem. 2015, 408, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Kurano, M.; Suzuki, A.; Inoue, A.; Tokuhara, Y.; Kano, K.; Matsumoto, H.; Igarashi, K.; Ohkawa, R.; Nakamura, K.; Dohi, T.; et al. Possible Involvement of Minor Lysophospholipids in the Increase in Plasma Lysophosphatidic Acid in Acute Coronary Syndrome. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Lanuti, M.; Talamonti, E.; Maccarrone, M.; Chiurchiù, V. Activation of GPR55 Receptors Exacerbates OxLDL-induced Lipid Accumulation and Inflammatory Responses, While Reducing Cholesterol Efflux from Human Macrophages. PLoS ONE 2015. [Google Scholar] [CrossRef]
- Falasca, M.; Ferro, R. Role of the Lysophosphatidylinositol/GPR55 Axis in Cancer. Adv. Biol. Regul. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ruban, E.L.; Ferro, R.; Arifin, S.A.; Falasca, M. Lysophosphatidylinositol: A novel link between ABC transporters and G-protein-coupled receptors. Biochem. Soc. Trans. 2014, 42, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Simcocks, A.C.; O’Keefe, L.; Jenkin, K.A.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. A Potential Role for GPR55 in the Regulation of Energy Homeostasis. Drug Discov. Today 2013. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Song, S.; Jones, P.M.; Persaud, S.J. GPR55: From Orphan to Metabolic Regulator? Pharmacol. Ther. 2015, 145, 35–42. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arifin, S.A.; Falasca, M. Lysophosphatidylinositol Signalling and Metabolic Diseases. Metabolites 2016, 6, 6. https://doi.org/10.3390/metabo6010006
AMA Style
Arifin SA, Falasca M. Lysophosphatidylinositol Signalling and Metabolic Diseases. Metabolites. 2016; 6(1):6. https://doi.org/10.3390/metabo6010006
Chicago/Turabian StyleArifin, Syamsul A., and Marco Falasca. 2016. "Lysophosphatidylinositol Signalling and Metabolic Diseases" Metabolites 6, no. 1: 6. https://doi.org/10.3390/metabo6010006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.