

Prediction of Potential Suitable Areas and Priority Protection for Cupressus gigantea on the Tibetan Plateau

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Optimal Model and Accuracy Evaluation
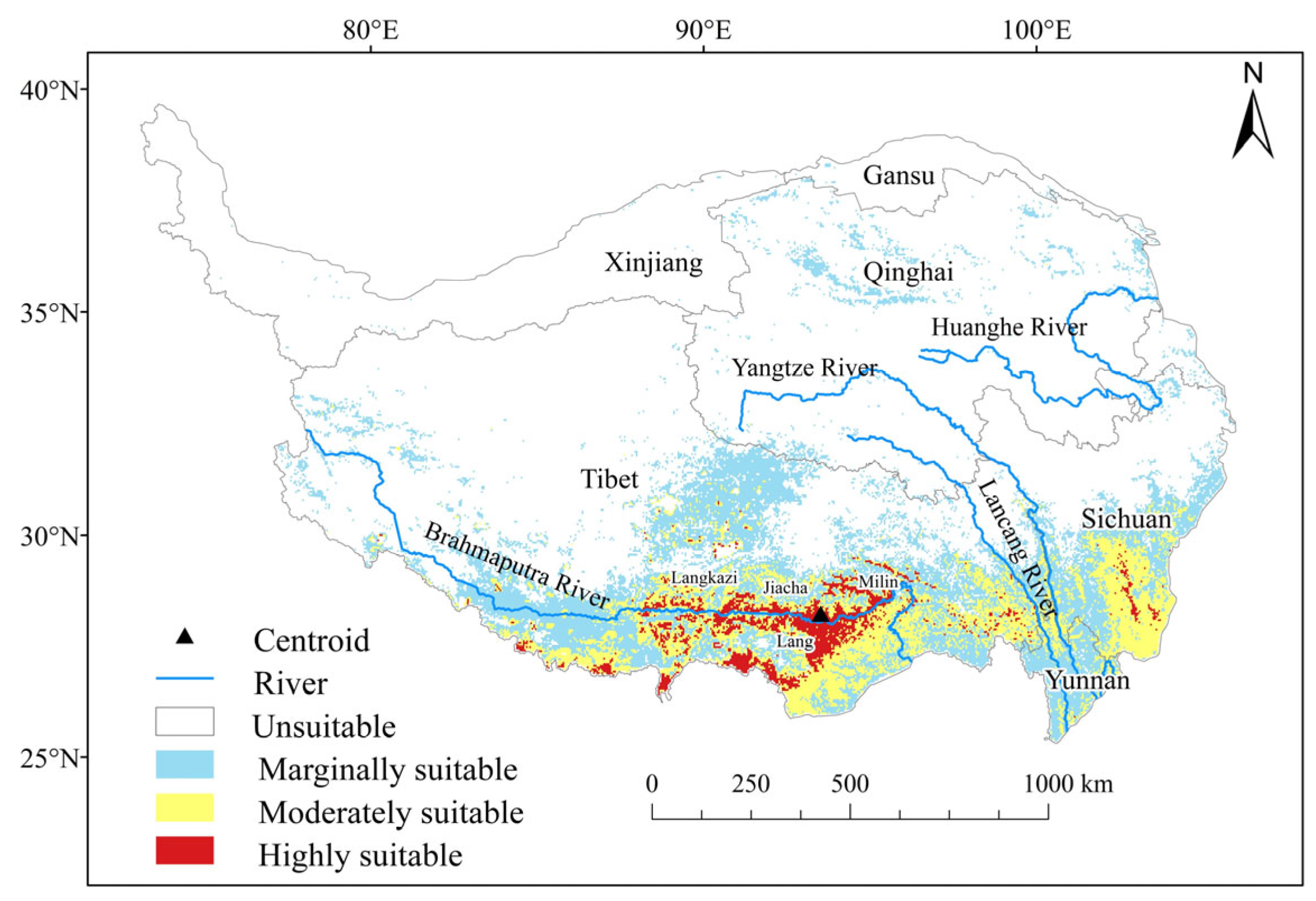
2.2. Current Potential Suitable Areas and Environmental Drivers
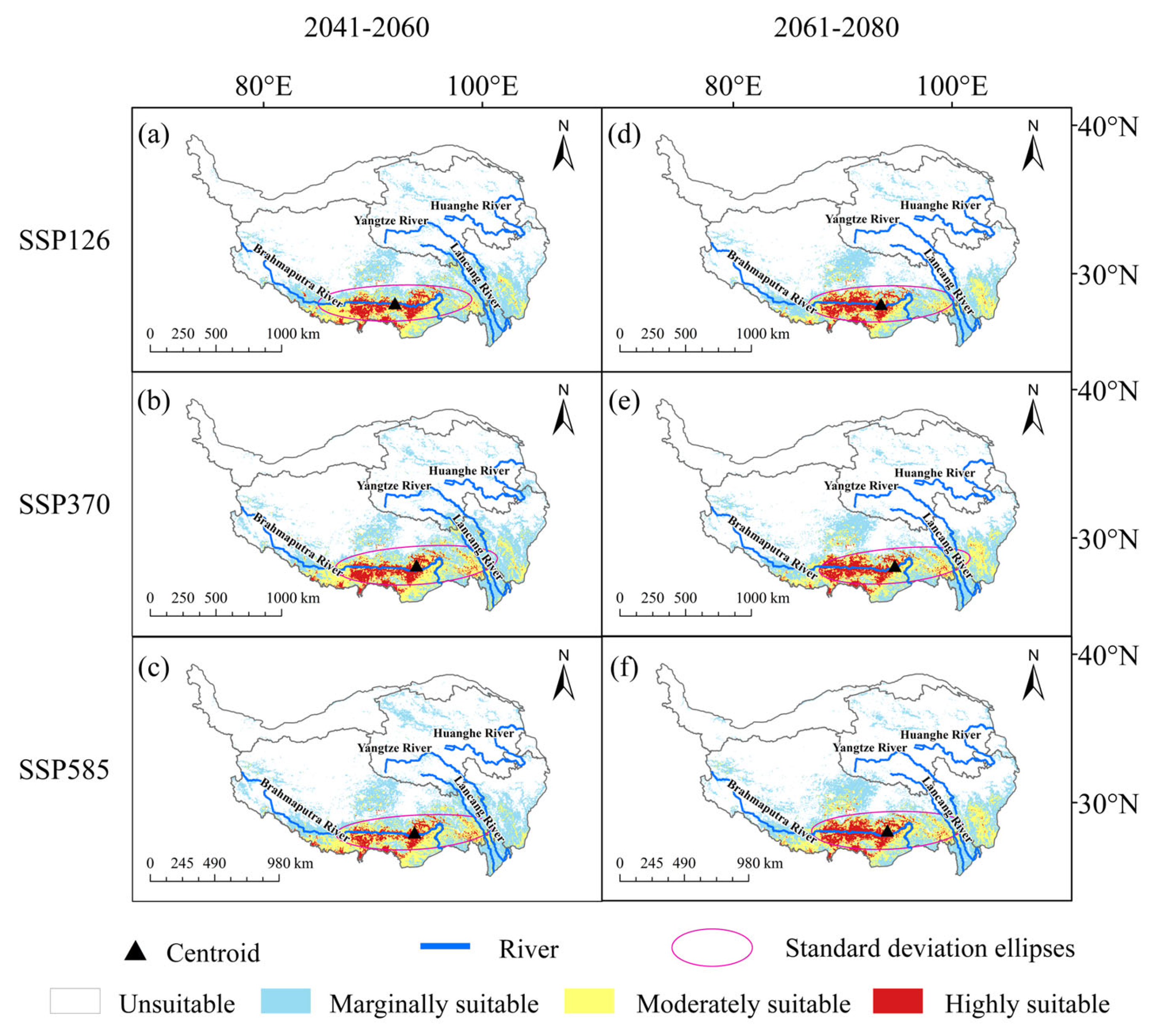
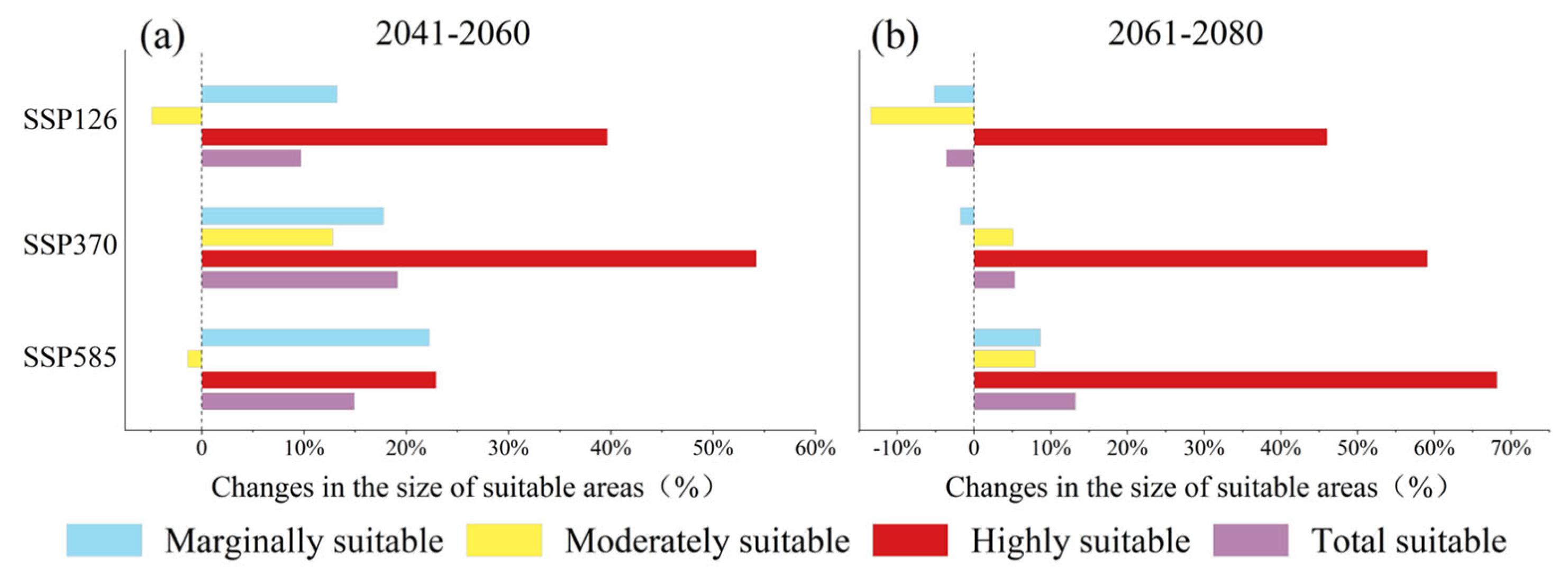
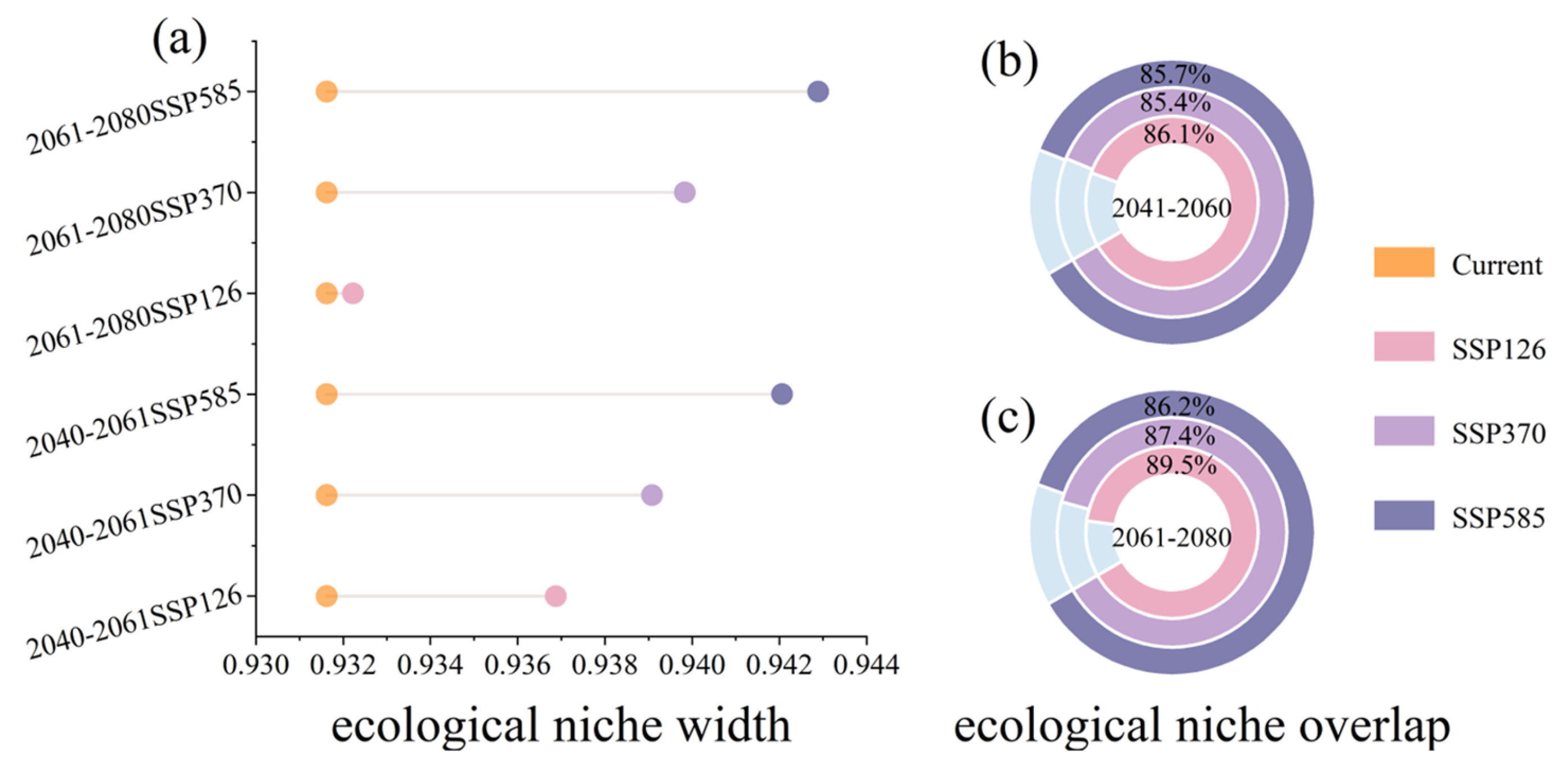
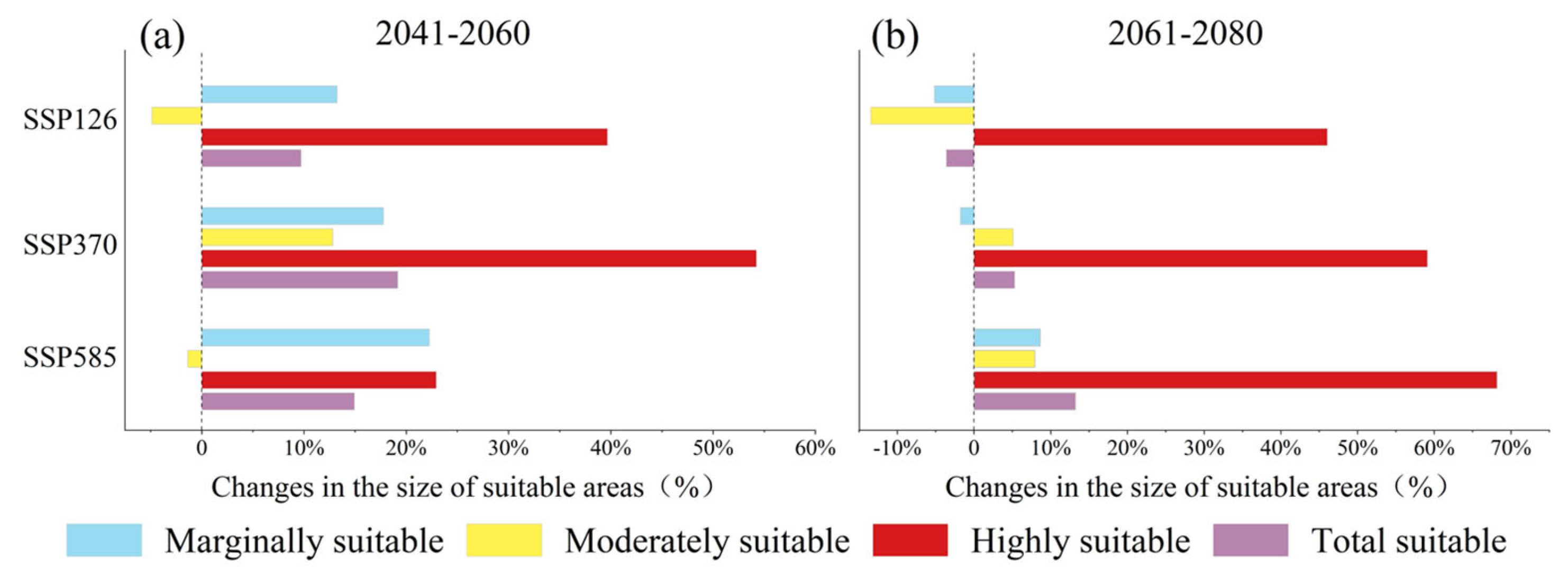
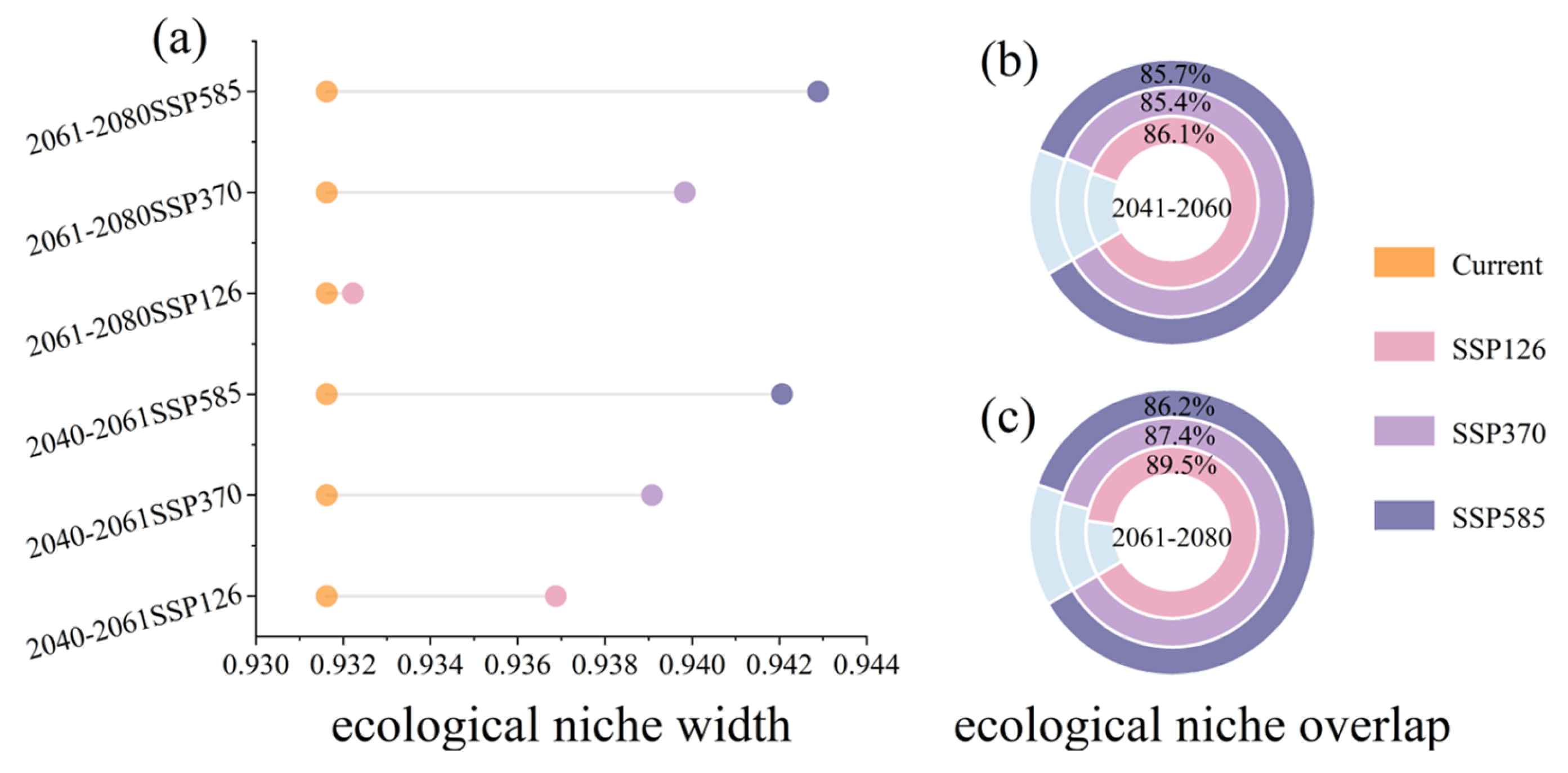
2.3. Future Contraction and Expansion of Potential Suitable Areas
2.4. Priority Protected Areas
3. Discussion
4. Materials and Methods
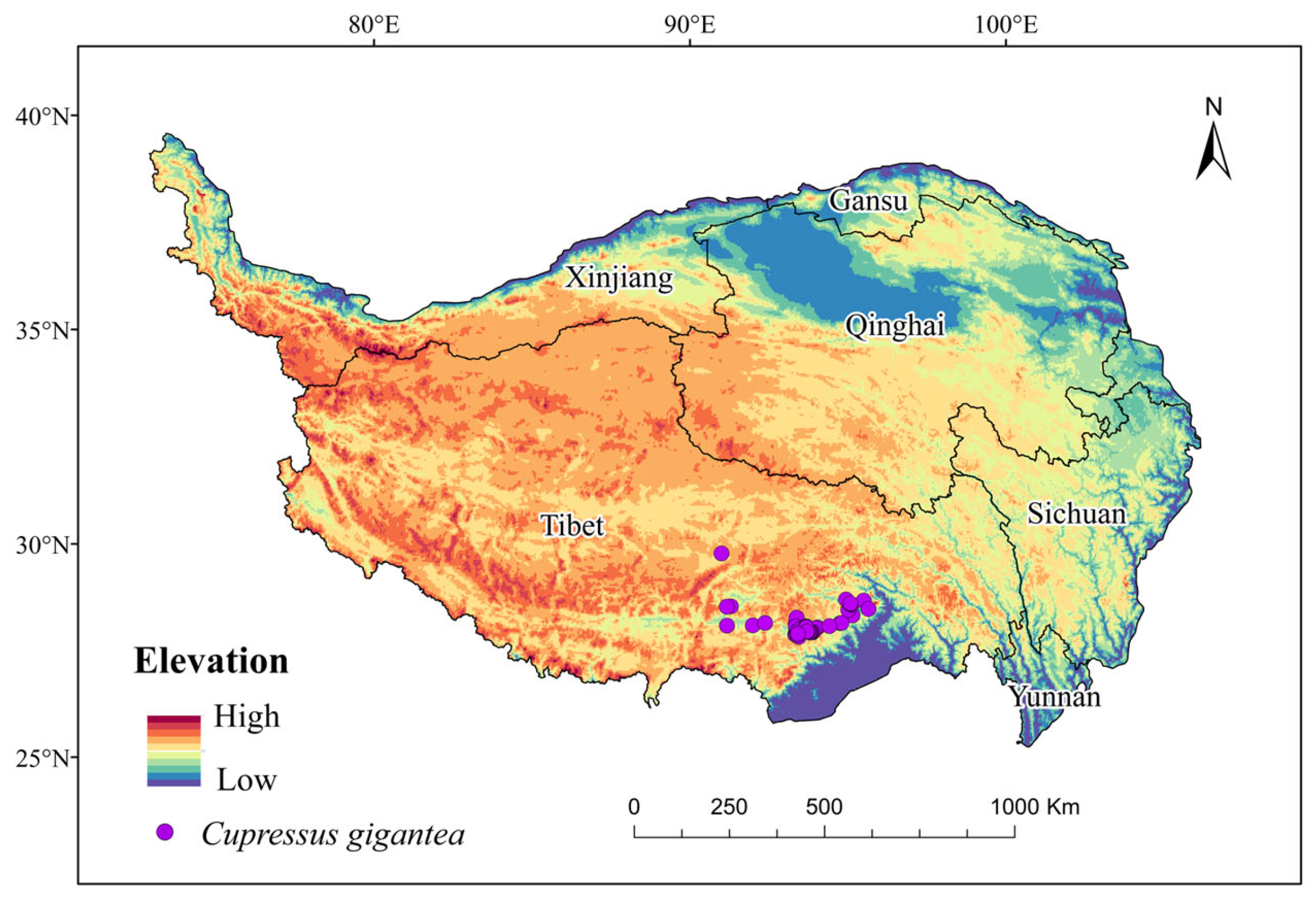
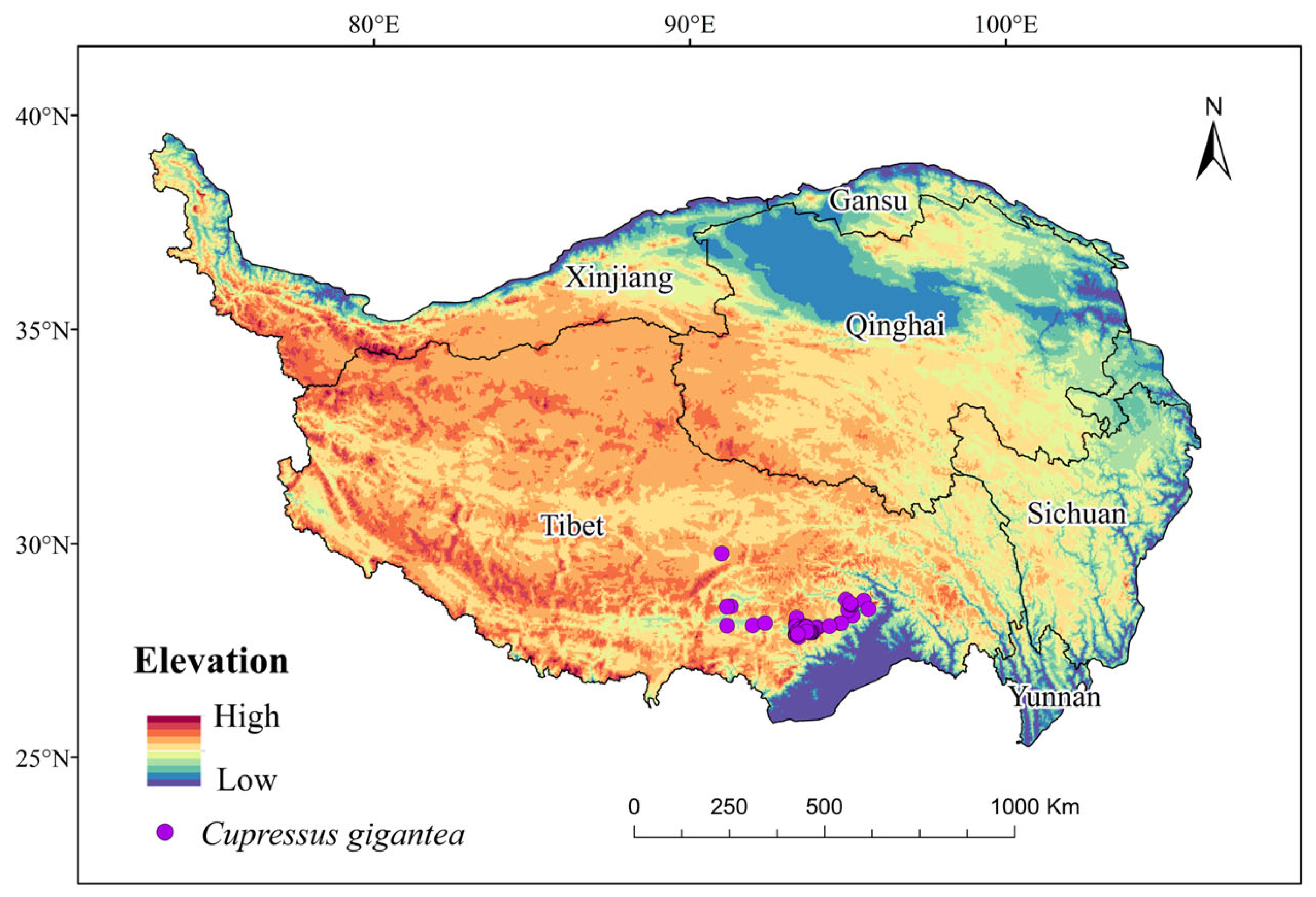
4.1. Data Collection and Assembly
4.2. Calculation of Potential Suitable Areas
4.3. Recognition of Priority Protection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guo, B.; Zang, W.; Yang, F.; Han, B.; Chen, S.; Liu, Y.; Yang, X.; He, T.; Chen, X.; Liu, C.; et al. Spatial and temporal change patterns of net primary productivity and its response to climate change in the Qinghai-Tibet Plateau of China from 2000 to 2015. J. Arid Land 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Shen, X.; Liu, Y.; Zhang, J.; Wang, Y.; Ma, R.; Liu, B.; Lu, X.; Jiang, M. Asymmetric impacts of diurnal warming on vegetation carbon sequestration of marshes in the Qinghai Tibet Plateau. Glob. Biogeochem. Cycles 2022, 36, e2022GB007396. [Google Scholar] [CrossRef]
- Duan, H.C.; Xue, X.; Wang, T.; Kang, W.P.; Liao, J.; Liu, S.L. Spatial and temporal differences in alpine meadow, Alpine Steppe and all vegetation of the Qinghai-Tibetan Plateau and Their Responses to climate change. Remote Sens. 2021, 13, 669. [Google Scholar] [CrossRef]
- Liu, J.H.; Xin, Z.B.; Huang, Y.Z.; Yu, J. Climate suitability assessment on the Qinghai-Tibet Plateau. Sci. Total Environ. 2022, 816, 151653. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, B.; Wen, A.M.; Xao, X.J. Predicting potential distribution of under global climate change in China. Appl. Ecol. Environ. Res. 2022, 20, 3977–3993. [Google Scholar] [CrossRef]
- Ma, Q.; Li, X.; Wu, S.; Zeng, F. Potential geographical distribution of Stipa purpurea across the Tibetan Plateau in China under climate change in the 21st century. Glob. Ecol. Conserv. 2022, 35, e02064. [Google Scholar] [CrossRef]
- Yin, Y.Y.; Wang, J.A.; Leng, G.Y.; Zhao, J.T.; Wang, L.; Ma, W.D. Future potential distribution and expansion trends of highland barley under climate change in the Qinghai-Tibet plateau (QTP). Ecol. Indic. 2022, 136, 108702. [Google Scholar] [CrossRef]
- Shi, N.; Naudiyal, N.; Wang, J.; Gaire, N.P.; Wu, Y.; Wei, Y.; He, J.; Wang, C. Assessing the impact of climate change on potential distribution of Meconopsis punicea and Its influence on ecosystem services Supply in the Southeastern Margin of Qinghai-Tibet Plateau. Front. Plant Sci. 2022, 12, 830119. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhou, S.; Yin, X.; Zhang, C.; Li, R.; Chen, J.; Ma, D.; Wang, Y.; Yu, Z.; Chen, Y. Patterns of tree species richness in Southwest China. Environ. Monit. Assess. 2021, 193, 97. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Li, W. Genetic diversity of Rheum tanguticum (Polygonaceae), an endangered species on Qinghai-Tibetan Plateau. Biochem. Syst. Ecol. 2016, 69, 132–137. [Google Scholar] [CrossRef]
- Xue, T.; Gadagkar, S.R.; Albright, T.P.; Yang, X.; Li, J.; Xia, C.; Wu, J.; Yu, S. Prioritizing conservation of biodiversity in an alpine region: Distribution pattern and conservation status of seed plants in the Qinghai-Tibetan Plateau. Glob. Ecol. Conserv. 2021, 32, e01885. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, N.; Duan, D.; Chen, Y. Research progress and expectation of Cupressus gigantea the rare and endemic plant in Xizang. J. Fujian For. Sci. Technol. 2005, 3, 160–164. [Google Scholar] [CrossRef]
- Yang, B.; Kang, X.; Bräuning, A.; Liu, J.; Qin, C.; Liu, J. A 622-year regional temperature history of southeast Tibet derived from tree rings. Holocene 2010, 20, 181–190. [Google Scholar] [CrossRef]
- Lu, X.; Xu, H.Y.; Li, Z.H.; Shang, H.Y.; Adams, R.P.; Mao, K.S. Genetic diversity and conservation implications of four species in China as revealed by Microsatellite markers. Biochem. Genet. 2014, 52, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gu, R.; Liu, B. Ethnobotanical study on wild aromatic plants used in a few Tibetan communities and Northwest of Yunnan. North. Hortic. 2017, 112–120. [Google Scholar]
- Fu, Y.R.; Li, S.K.; Guo, Q.Q.; Zheng, W.L.; Yang, R.; Li, H.E. Genetic diversity and population structure of two endemic (Cupressaceae) species on the Qinghai-Tibetan plateau. J. Genet. 2019, 98, 14. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Y.; Chen, Q.; Liu, C.; Liu, G.; Cheng, S.; Wang, L. Potential spatiotemporal distribution changes and conservation recommendations of two connected endangered tree peony species (Paeonia decomposita & P. rotundiloba). Flora 2022, 294, 152131. [Google Scholar] [CrossRef]
- Chi, X.; Zheng, W.; Guo, Q.; Wang, S.; Li, H. Primary research on seed plant flora of Cupressus gigantea froest and Cupressus torulosa forest in Southeast Tibet. J. Plateau Agric. 2018, 2, 12. [Google Scholar] [CrossRef]
- Li, Y.; Yang, X.; Zhao, K.; Yao, Q.; Zhao, Y. Research on the growth characteristics and model of Cupressus gigantea in southeast Tibet. J. Plateau Agric. 2020, 4, 269–276. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, W.; Pan, G. Relation between being endangered and seed vigor about Cupressus gigantea in Tibet. Sci. Silvae Sin. 2005, 41, 37–41. [Google Scholar] [CrossRef]
- Xia, T.; Meng, L.; Mao, K.; Tian, B.; Miehe, G.; Liu, J. Genetic variation in the Qinghai-Tibetan Plateau endemic and endangered conifer, detected using RAPD and ISSR markers. Silvae Genet. 2008, 57, 85–92. [Google Scholar] [CrossRef]
- He, P.; Li, J.; Li, Y.; Xu, N.; Gao, Y.; Guo, L.; Huo, T.; Peng, C.; Meng, F. Habitat protection and planning for three Ephedra using the MaxEnt and Marxan models. Ecol. Indic. 2021, 133, 108399. [Google Scholar] [CrossRef]
- Wang, Y.J.; Zhao, R.X.; Zhou, X.Y.; Zhang, X.L.; Zhao, G.H.; Zhang, F.G. Prediction of potential distribution areas and priority protected areas of based on Maxent model and Marxan model. Front. Plant Sci. 2023, 14, 1200796. [Google Scholar] [CrossRef] [PubMed]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Alarcón, D.; Cavieres, L.A. Relationships between ecological niche and expected shifts in elevation and latitude due to climate change in South American temperate forest plants. J. Biogeogr. 2018, 45, 2272–2287. [Google Scholar] [CrossRef]
- Guo, L.F.; Gao, Y.; He, P.; He, Y.; Meng, F.Y. Modeling for predicting the potential geographical distribution of three Herbs in China. Plants 2023, 12, 787. [Google Scholar] [CrossRef]
- Zhao, Z.F.; Wei, H.Y.; Guo, Y.L.; Gu, W. Potential distribution of Panax ginseng and its predicted responses to climate change. Chin. J. Appl. Ecol. 2016, 27, 3607–3615. [Google Scholar] [CrossRef]
- Wani, I.A.; Khan, S.; Verma, S.; Al-Misned, F.A.; Shafik, H.M.; El-Serehy, H.A. Predicting habitat suitability and niche dynamics of Dactylorhiza hatagirea and Rheum webbianum in the Himalaya under projected climate change. Sci. Rep. 2022, 12, 13205. [Google Scholar] [CrossRef]
- Xu, W.; Xiao, Y.; Zhang, J.; Yang, W.; Zhang, L.; Hull, V.; Wang, Z.; Zheng, H.; Liu, J.; Polasky, S.; et al. Strengthening protected areas for biodiversity and ecosystem services in China. Proc. Natl. Acad. Sci. USA 2017, 114, 1601–1606. [Google Scholar] [CrossRef]
- Tang, J.; Lu, H.; Xue, Y.; Li, J.; Li, G.; Mao, Y.; Deng, C.; Li, D. Data-driven planning adjustments of the functional zoning of Houhe National Nature Reserve. Glob. Ecol. Conserv. 2021, 29, e01708. [Google Scholar] [CrossRef]
- Lin, L.; He, J.; Lyu, R.; Luo, Y.; Yao, M.; Xie, L.; Cui, G. Targeted conservation management of white pines in China: Integrating phylogeographic structure, niche modeling, and conservation gap analyses. For. Ecol. Manag. 2021, 492, 119211. [Google Scholar] [CrossRef]
- Carrasco, J.; Price, V.; Tulloch, V.; Mills, M. Selecting priority areas for the conservation of endemic trees species and their ecosystems in Madagascar considering both conservation value and vulnerability to human pressure. Biodivers. Conserv. 2020, 29, 1841–1854. [Google Scholar] [CrossRef]
- Ball, I.R.; Possingham, H.P.; Watts, M.E. Marxan and relatives: Software for spatial conservation prioritization. In Spatial Conservation Prioritization: Quantitative Methods and Computational Tools; Moilanen, A., Wilson, K.A., Possingham, H.P., Eds.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Yin, J.; Zhao, K.; Zou, L. Impact of different substrates and temperatures on the growth and development of Cupressus gigantea transplant seedling roots. J. West China For. Sci. 2017, 46, 87–92. [Google Scholar] [CrossRef]
- Peng, J.; Liu, Z.; Liu, Y.; Wu, J.; Han, Y. Trend analysis of vegetation dynamics in Qinghai–Tibet Plateau using Hurst Exponent. Ecol. Indic. 2012, 14, 28–39. [Google Scholar] [CrossRef]
- Fan, Z.; Bai, X. Scenarios of potential vegetation distribution in the different gradient zones of Qinghai-Tibet Plateau under future climate change. Sci. Total Environ. 2021, 796, 148918. [Google Scholar] [CrossRef]
- Luo, D.; Qu, X.; Wang, H. Analysis of the characteristics of vertical microclimate in the ecological distribution area of Cupressus gigantea. J. Plateau Agric. 2017, 1, 22–26+72. [Google Scholar] [CrossRef]
- Xin, F.; Liu, W.; Zhang, X.; Li, Z. Effects of different fertilization methods on growth and photosynthesis of Cupressus gigantea seedlings. J. Xiamen Univ. 2022, 61, 59–69. [Google Scholar]
- Parmesan, C.; Hanley, M.E. Plants and climate change: Complexities and surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Gardner, A.S.; Maclean, I.M.D.; Gaston, K.J. Climatic predictors of species distributions neglect biophysiologically meaningful variables. Divers. Distrib. 2019, 25, 1318–1333. [Google Scholar] [CrossRef]
- Hellegers, M.; Ozinga, W.A.; van Hinsberg, A.; Huijbregts, M.A.J.; Hennekens, S.M.; Schaminée, J.H.J.; Dengler, J.; Schipper, A.M. Evaluating the ecological realism of plant species distribution models with ecological indicator values. Ecography 2020, 43, 161–170. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, J.; Dai, W.; Ye, K.; Chen, J.; Lai, Q.; Li, H.; Zhong, B.; Yu, X. Effects of climate warming and human activities on the distribution patterns of Fritillaria unibracteata in eastern Qinghai-Tibetan Plateau. Sci. Rep. 2023, 13, 15770. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, W.; Liu, B.; Yuan, Y.; Bao, A. Monitoring vegetation sensitivity to drought events in China. Sci. Total Environ. 2023, 893, 164917. [Google Scholar] [CrossRef]
- Liu, Y.; Bi, S.; Yu, Q. A study on forest soil under Cupressus gigantea forest in Tibet. Sci. Silvae Sin. 1988, 24, 466–470. [Google Scholar]
- Mölders, N. Plant and soil parameter-caused uncertainty of predicted surface fluxes. Mon. Weather Rev. 2005, 133, 3498–3516. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.; Lei, L.; Yan, D. Monitoring vegetation greenness variations in Qinghai-Tibet Plateau with MODIS vegetation index. In Proceedings of the 2011 IEEE International Geoscience and Remote Sensing Symposium, Vancouver, BC, Canada, 24–29 July 2011; pp. 760–762. [Google Scholar]
- Yu, L.; Cao, M.; Li, K. Climate-induced changes in the vegetation pattern of China in the 21st century. Ecol. Res. 2006, 21, 912–919. [Google Scholar] [CrossRef]
- Zhong, L.; Ma, Y.; Xue, Y.; Piao, S. Climate change trends and impacts on vegetation greening over the Tibetan Plateau. J. Geophys. Res. Atmos. 2019, 124, 7540–7552. [Google Scholar] [CrossRef]
- Lemordant, L.; Gentine, P. Vegetation response to rising CO2 impacts extreme temperatures. Geophys. Res. Lett. 2019, 46, 1383–1392. [Google Scholar] [CrossRef]
- Raghunathan, N.; François, L.; Dury, M.; Hambuckers, A. Contrasting climate risks predicted by dynamic vegetation and ecological niche-based models applied to tree species in the Brazilian Atlantic Forest. Reg. Environ. Chang. 2019, 19, 219–232. [Google Scholar] [CrossRef]
- Comer, P.J.; Hak, J.C.; Reid, M.S.; Auer, S.L.; Schulz, K.A.; Hamilton, H.H.; Smyth, R.L.; Kling, M.M. Habitat climate change vulnerability index applied to major vegetation types of the Western Interior United States. Land 2019, 8, 108. [Google Scholar] [CrossRef]
- Gallagher, R.V.; Allen, S.; Wright, I.J. Safety margins and adaptive capacity of vegetation to climate change. Sci. Rep. 2019, 9, 8241. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Li, W.; Wang, J.; Du, F.; Yang, Y.M. Effects of disturbance on niche breadth, overlap, and fifferentiation of dominant plant species in Napa Plateau Wetland, Shangrila. In Proceedings of the 2013 Third International Conference on Intelligent System Design and Engineering Applications, Hong Kong, China, 16–18 January 2013; pp. 666–670. [Google Scholar] [CrossRef]
- Pierotti, M.E.R.; Martín-Fernández, J.A.; Barceló-Vidal, C. The peril of proportions: Robust niche indices for categorical data. Methods Ecol. Evol. 2017, 8, 223–231. [Google Scholar] [CrossRef]
- Zheng, W.; Xue, H.; Luo, D.; Wang, J.; Luo, J. Eco-geographic distribution and coenology characteristics of Cupressus gigantea. Sci. Silvae Sin. 2007, 43, 8–15. [Google Scholar]
- Harvey, B.J.; Cook, P.; Shaffrey, L.C.; Schiemann, R. The Response of the Northern Hemisphere Storm Tracks and Jet Streams to Climate Change in the CMIP3, CMIP5, and CMIP6 Climate Models. J. Geophys. Res. Atmos. 2020, 125, e2020JD032701. [Google Scholar] [CrossRef]
- Zhu, H.; Jiang, Z.; Li, J.; Li, W.; Sun, C.; Li, L. Does CMIP6 Inspire More Confidence in Simulating Climate Extremes over China? Adv. Atmos. Sci. 2020, 37, 1119–1132. [Google Scholar] [CrossRef]
- Tokarska, K.B.; Stolpe, M.B.; Sippel, S.; Fischer, E.M.; Smith, C.J.; Lehner, F.; Knutti, R. Past warming trend constrains future warming in CMIP6 models. Sci. Adv. 2020, 6, eaaz9549. [Google Scholar] [CrossRef]
- Ma, D.; Lun, X.; Li, C.; Zhou, R.; Zhao, Z.; Wang, J.; Zhang, Q.; Liu, Q. Predicting the Potential Global Distribution of Amblyomma americanum (Acari: Ixodidae) under Near Current and Future Climatic Conditions, Using the Maximum Entropy Model. Biology 2021, 10, 1057. [Google Scholar] [CrossRef]
- Tu, W.; Xiong, Q.; Qiu, X.; Zhang, Y. Dynamics of invasive alien plant species in China under climate change scenarios. Ecol. Indic. 2021, 129, 107919. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, J.; Wang, M.; Li, Z.; Lin, S.; Chen, H. Potential distribution prediction of Amaranthus palmeri S. Watson in China under current and future climate scenarios. Ecol. Evol. 2022, 12, e9505. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Rathore, P.; Roy, A.; Karnatak, H. Modelling the vulnerability of Taxus wallichiana to climate change scenarios in South East Asia. Ecol. Indic. 2019, 102, 199–207. [Google Scholar] [CrossRef]
- Jenks, G.F. The Data Model Concept in Statistical Mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Evans, A.; Jacquemyn, H. Range size and niche breadth as predictors of climate-induced habitat change in Epipactis (Orchidaceae). Front. Ecol. Evol. 2022, 10, 894616. [Google Scholar] [CrossRef]
- Jaime, R.; Alcántara, J.M.; Bastida, J.M.; Rey, P.J. Complex patterns of environmental niche evolution in Iberian columbines (genus Aquilegia, Ranunculaceae). J. Plant Ecol. 2015, 8, 457–467. [Google Scholar] [CrossRef]
- Galvis-Martinez, C.A.; Moo-Llanes, D.A.; Altamiranda-Saavedra, M. Similarity but not equivalence: Ecological niche comparison between sandflies from the Pleistocene and future scenarios in Central and South America. Med. Vet. Entomol. 2023, 37, 111–123. [Google Scholar] [CrossRef]
- Luck, G.W.; Ricketts, T.H.; Daily, G.C.; Imhoff, M. Alleviating spatial conflict between people and biodiversity. Proc. Natl. Acad. Sci. USA 2004, 101, 182–186. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Evaluation Index | Model | Evaluation Index | ||
|---|---|---|---|---|---|
| AUC | TSS | AUC | TSS | ||
| XGBOOST | 0.999 | 0.999 | FDA | 0.999 | 0.990 |
| RF | 0.999 | 0.999 | MARS | 0.997 | 0.975 |
| GBM | 0.999 | 0.996 | MAXENT | 0.980 | 0.942 |
| GLM | 0.999 | 0.992 | ANN | 0.956 | 0.900 |
| CTA | 0.976 | 0.922 | SRE | 0.881 | 0.749 |
| EM | 0.999 | 0.999 | |||
| Category | Abbreviations | Environmental Variables | Important Value (%) |
|---|---|---|---|
| Temperature | MAT | Annual mean temperature | 1.104 |
| Precipitation | MDR | Mean diurnal range | 1.313 |
| Iso | Isothermality | 0.782 | |
| TSN | Temperature seasonality | 32.343 | |
| ART | Temperature annual range | 24.302 | |
| MAP | Annual precipitation | 2.127 | |
| PDQ | Precipitation of driest quarter | 9.858 | |
| PCOQ | Precipitation of coldest quarter | 0.709 | |
| Soil texture | SAND | Sand content (%) | 16.047 |
| BDOD | Bulk density (kg/m3) | 0.178 | |
| CFVO | Coarse fragments volumetric (%) | 0.646 | |
| Soil fertility | NITROGEN | Total nitrogen (g/kg) | 1.603 |
| SOC | Soil organic carbon content (g/kg) | 3.906 | |
| CEC | Cation exchange capacity (cmolc/kg) | 4.069 | |
| Terrain | ELE | Elevation(m) | 0.388 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wei, Y.; Yue, J.; Wang, Z.; Zou, H.; Ji, X.; Zhang, S.; Liu, Z. Prediction of Potential Suitable Areas and Priority Protection for Cupressus gigantea on the Tibetan Plateau. Plants 2024, 13, 896. https://doi.org/10.3390/plants13060896
Zhang H, Wei Y, Yue J, Wang Z, Zou H, Ji X, Zhang S, Liu Z. Prediction of Potential Suitable Areas and Priority Protection for Cupressus gigantea on the Tibetan Plateau. Plants. 2024; 13(6):896. https://doi.org/10.3390/plants13060896
Chicago/Turabian StyleZhang, Huayong, Yanan Wei, Junjie Yue, Zhongyu Wang, Hengchao Zou, Xiande Ji, Shijia Zhang, and Zhao Liu. 2024. "Prediction of Potential Suitable Areas and Priority Protection for Cupressus gigantea on the Tibetan Plateau" Plants 13, no. 6: 896. https://doi.org/10.3390/plants13060896