
Development of a TaqMan Real-Time PCR for Early and Accurate Detection of Anthracnose Pathogen Colletotrichum siamense in Pachira glabra

Abstract
:1. Introduction
2. Results
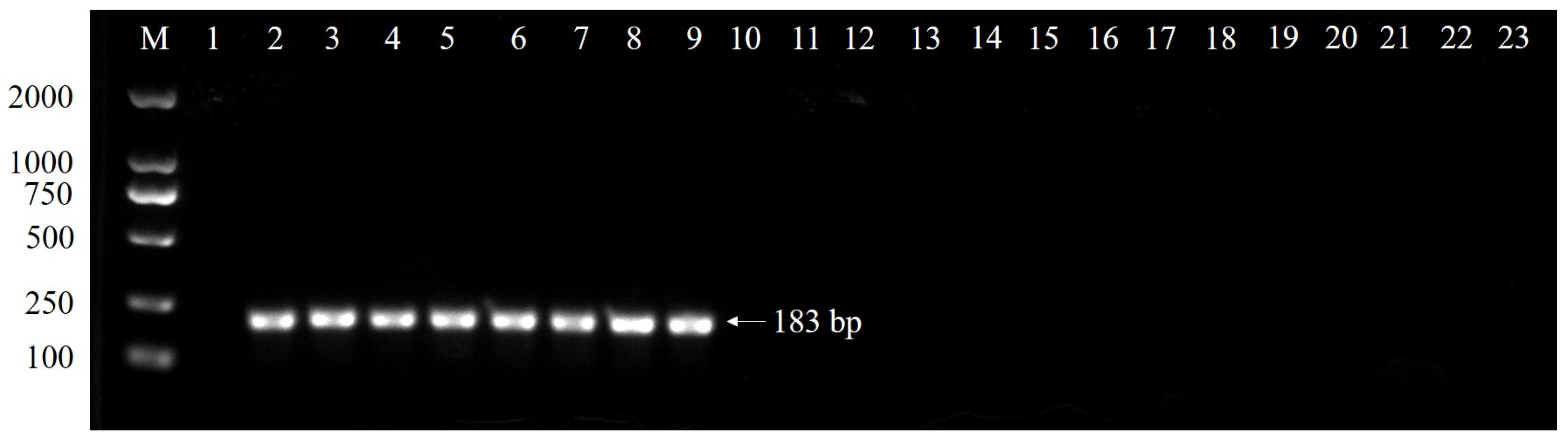
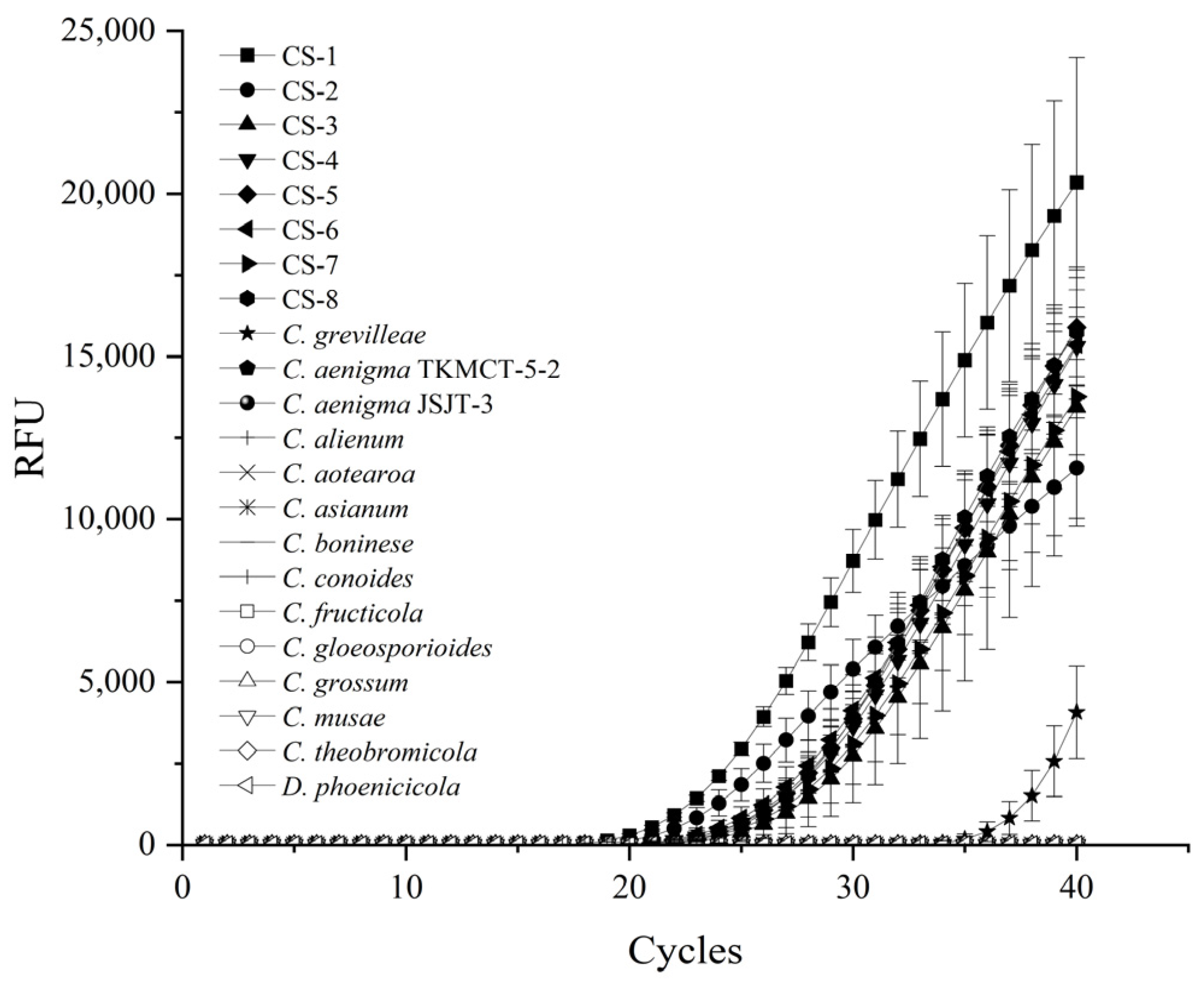
2.1. Specificity of Primer Set and TaqMan Probe
2.2. Sensitivity and Stability of the TaqMan Real-Time PCR Assay
2.3. Detection in Artificially Inoculated P. glabra
2.4. Detection in Naturally Infected P. glabra
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates
4.2. Plant Growth
4.3. DNA Extraction
4.4. Cloning of the Target Sequence
4.5. Real-Time Primers and TaqMan Probe Design
4.6. TaqMan Real-Time PCR Assay
4.7. Sensitivity and Stability of the TaqMan Real-Time PCR Assay
4.8. Validity of the TaqMan Real-Time PCR on Artificially Inoculated Plants
4.9. Validity of the TaqMan Real-Time PCR on Naturally Infected Plants in Fields
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milagres, C.A.; Belisário, R.; Silva, M.A.; Lisboa, D.O.; Pinho, D.B.; Furtado, G.Q. A novel species of Diaporthe causing leaf spot in Pachira glabra. Trop. Plant Pathol. 2018, 43, 460–467. [Google Scholar] [CrossRef]
- Lawal, O.A.; Ogunwande, I.A.; Salvador, A.F.; Sanni, A.A.; Opoku, A.R. Pachira glabra Pasq. essential oil: Chemical constituents, antimicrobial and insecticidal activities. J. Oleo Sci. 2014, 63, 629–635. [Google Scholar] [CrossRef] [PubMed]
- El-Din, M.I.G.; Youssef, F.S.; Said, R.S.; Ashour, M.L.; Eldahshan, O.A.; Singab, A.N.B. Chemical constituents and gastro-protective potential of Pachira glabra leaves against ethanol-induced gastric ulcer in experimental rat model. Inflammopharmacology 2021, 29, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.H.; Peng, X.X.; Wu, Q.T.; Wen, B.Y.; Deng, C.C.; Wang, H.H. First report of white leaf spot caused by Hypomontagnella monticulosa on Pachira glabra in China. Plant Dis. 2023, 107, 2244. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.T.; Xiao, Z.Y.; Tian, J.H.; Li, S.Q.; Peng, X.X.; Wang, H.H. First report of anthracnose caused by Colletotrichum siamense on Pachira glabra in China. Plant Dis. 2023, 107, 572. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.C.; Wang, H.H.; Wen, B.Y.; Gu, J.Q.; Peng, X.X.; Zhang, Z.F. First report of leaf brown spot caused by Diaporthe phoenicicola on Pachira glabra in China. Plant Dis. 2024, in press. [CrossRef] [PubMed]
- Jayawardena, R.S.; Hyde, K.D.; Damm, U.; Cai, L.; Liu, M.; Li, X.H.; Zhang, W.; Zhao, W.S.; Yan, J.Y. Notes on currently accepted species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [Google Scholar] [CrossRef]
- Khodadadi, F.; González, J.; Martin, P.L.; Giroux, E.; Bilodeau, G.J.; Peter, K.A.; Doyle, V.P.; Aćimović, S.G. Identification and characterization of Colletotrichum species causing apple bitter rot in New York and description of C. noveboracense sp. nov. Sci. Rep. 2020, 10, 11043. [Google Scholar] [CrossRef]
- Sharma, G.; Maymon, M.; Freeman, S. Epidemiology, pathology and identification of Colletotrichum including a novel species associated with avocado (Persea americana) anthracnose in Israel. Sci. Rep. 2017, 7, 15839. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum: Current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Cai, L.; Ehc, M.K.; Yang, Y.L.; Prihastuti, H. Colletotrichum: A catalogue of confusion. Fungal Divers. 2009, 39, 1–17. [Google Scholar]
- Vieira, W.A.; Bezerra, P.A.; da Silva, A.C.; Veloso, J.S.; Câmara, M.P.S.; Doyle, V.P. Optimal markers for the identification of Colletotrichum species. Mol. Phylogenet Evol. 2020, 143, 106694. [Google Scholar] [CrossRef] [PubMed]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Khakimov, A.; Salakhutdinov, I.; Omolikov, A.; Utaganov, S. Traditional and current-prospective methods of agricultural plant diseases detection: A review. IOP Conf. Ser. Earth Environ. Sci. 2022, 951, 012002. [Google Scholar] [CrossRef]
- Kaur, H.; Singh, R.; Doyle, V.P.; Valverde, R. A diagnostic TaqMan real-time PCR assay for in planta detection and quantification of Colletotrichum theobromicola, causal agent of boxwood dieback. Plant Dis. 2021, 105, 2395–2401. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wang, M.; Zou, L.; Long, M.; Yang, Y.; Zhang, Y.; Liang, X. Quantitative detection and monitoring of Colletotrichum siamense in rubber trees using real-time PCR. Plant Dis. 2021, 105, 2861–2866. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Duan, K.; Liu, Y.; Song, L.; Gao, Q.H. Method to detect and quantify colonization of anthracnose causal agent Colletotrichum gloeosporioides species complex in strawberry by real-time PCR. J. Phytopathol. 2022, 170, 326–336. [Google Scholar] [CrossRef]
- Wan, M.; Yang, L.; Zhang, S.; Gao, J.; Jiang, L.; Luo, L. Real-time PCR for detection and quantification of C. gloeosporioides s.l. growth in Stylosanthes and Arabidopsis. Crop Prot. 2022, 159, 106021. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Conner, R.L.; Gillard, C.L.; Mclaren, D.L.; Boland, G.J.; Balasubramanian, P.M.; Stasolla, C.; Zhou, Q.X.; Hwang, S.F.; Chang, K.F.; et al. A quantitative real-time PCR assay for detection of Colletotrichum lindemuthianum in navy bean seeds. Plant Pathol. 2013, 62, 900–907. [Google Scholar] [CrossRef]
- Tao, G.; Hyde, K.; Cai, L. Species-specific real-time PCR detection of Colletotrichum kahawae. J. Appl. Microbiol. 2013, 114, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Kong, D.; Teng, S.; Deng, Z. Establishment of a TaqMan real-time PCR test method for detecting Colletotrichum karstii. Guizhou Agric. Sci. China 2017, 45, 5–9. [Google Scholar]
- Wang, C.; Zhang, X.J.; Zhang, X.L.; Zhang, W.; Zhang, Y.; Li, Y.W.; Sun, Y.F. Detection of Colletotrichum orbiculare by the real-time PCR. Plant Prot. China 2016, 42, 129–135. [Google Scholar]
- He, J.; Sun, M.L.; Li, D.W.; Zhu, L.H.; Ye, J.R.; Huang, L. A real-time PCR for detection of pathogens of anthracnose on Chinese fir using TaqMan probe targeting ApMat gene. Pest Manag. Sci. 2023, 79, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Dowling, M.; Peres, N.; Villani, S.; Schnabel, G. Managing Colletotrichum on fruit crops: A “complex” challenge. Plant Dis. 2020, 104, 2301–2316. [Google Scholar] [CrossRef] [PubMed]
- Hayden, K.J.; Rizzo, D.; Tse, J.; Garbelotto, M.; Matthiesen, R.L.; Schmidt, C.; Robertson, A.E.; Du, Y.; Wang, M.; Long, M.; et al. Detection and quantification of Phytophthora ramorum from California forests using a real-time polymerase chain reaction assay. Phytopathology 2004, 94, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Vieira, W.A.; Lima, W.G.; Nascimento, E.S.; Michereff, S.J.; Câmara, M.P.; Doyle, V.P. The impact of phenotypic and molecular data on the inference of Colletotrichum diversity associated with Musa. Mycologia 2017, 109, 912–934. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Li, T.; Zhang, J.; Shao, H.; Kageyama, K.; Feng, W. Rapid detection of Colletotrichum siamense from infected tea plants using filter-disc DNA extraction and loop-mediated isothermal amplification. Plant Dis. 2024, 108, 35–40. [Google Scholar] [CrossRef]
- Schena, L.; Nigro, F.; Ippolito, A.; Gallitelli, D. Real-time quantitative PCR: A new technology to detect and study phytopathogenic and antagonistic fungi. Eur. J. Plant Pathol. 2004, 110, 893–908. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internaltranscribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Ginzinger, D.G. Gene amplification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Species | Strains | Host | Origin | Ct Value |
|---|---|---|---|---|---|
| 1 | C. aenigma | TKMCT-5-2 | Trichosanthes kirilowii Maxim | China | >40 |
| 2 | JSJT-3 | Carya illinoinensis | Jiangsu, China | >40 | |
| 3 | C. alienum | CBS 113192 | Protea cynaroides | South Africa | >40 |
| 4 | C. aotearoa | CBS 114139 | Knightia sp. | New Zealand | >40 |
| 5 | C. asianum | SY04S | Mangifera indica L. | Guangxi/Guizhou/Hainan, China | >40 |
| 6 | C. boninese | JFRL-03-35 | Aucuba japonica | Guizhou, China | >40 |
| 7 | C. conoides | CAUG17 | Capsicum annuum | Jiangsu, China | >40 |
| 8 | C. fructicola | HNLD-10 | Fragaria ananassa | China | >40 |
| 9 | C. gloeosporioides | C20 | Robinia pseudoacacia | Shandong, China | >40 |
| 10 | C. grevilleae | CBS 132879 | Grevillea sp. | Italy | 39.2 |
| 11 | C. grossum | CAUG7 | Coprosma sp. | Hainan, China | >40 |
| 12 | C. musae | LJ1-2 | Musa spp. | Guangxi, China | >40 |
| 13 | C. siamense | CS-1 | Pachira glabra | Hunan, China | 22.65 |
| 14 | CS-2 | Pachira glabra | Hunan, China | 23.97 | |
| 15 | CS-3 | Pachira glabra | Hunan, China | 27.74 | |
| 16 | CS-4 | Pachira glabra | Hunan, China | 26.56 | |
| 17 | CS-5 | Pachira glabra | Hunan, China | 26.42 | |
| 18 | CS-6 | Pachira glabra | Hunan, China | 26.15 | |
| 19 | CS-7 | Pachira glabra | Guangdong, China | 27.86 | |
| 20 | CS-8 | Pachira glabra | Hainan, China | 26.65 | |
| 21 | C. theobromicola | CCOD4 | Gossypium hirsutum L. | Guizhou/Hubei/Henan, China | >40 |
| 22 | D. phoenicicola | gpg2023-1 | Pachira glabra | Hunan, China | >40 |
| Templates | Pachira glabra Extract (μL) | Ct (Mean ± SD) |
|---|---|---|
| 5 ng CS-1 gDNA | 2.5 | 28.61 ± 0.23 |
| 2.0 | 29.50 ± 0.81 | |
| 1.5 | 29.74 ± 1.06 | |
| 1.0 | 28.10 ± 0.32 | |
| 0.5 | 28.43 ± 0.42 | |
| 0 | 27.95 ± 0.16 | |
| p = 0.785 | ||
| Disease Grade | Number of Leaves | CS-1 DNA Amount [log (pg per 50 ng of Sample DNA)] | Detection with Conventional PCR 1 | Detection with the Tissue Isolation Method 1 |
|---|---|---|---|---|
| 0 | 18 | 0 | ND | ND |
| 2 | 1.93 ± 0.42 | ND | D | |
| 5 | D | D | ||
| 1 | 3 | 2.65 ± 0.14 | D | D |
| 2 | 3 | 3.24 ± 0.07 | D | D |
| 3 | 3 | 3.46 ± 0.10 | D | D |
| 4 | 3 | 3.95 ± 0.09 | D | D |
| 5 | 3 | 4.14 ± 0.12 | D | D |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Wang, H.; Huang, X.; Liao, L.; Xie, H.; Peng, X. Development of a TaqMan Real-Time PCR for Early and Accurate Detection of Anthracnose Pathogen Colletotrichum siamense in Pachira glabra. Plants 2024, 13, 1149. https://doi.org/10.3390/plants13081149
Gu J, Wang H, Huang X, Liao L, Xie H, Peng X. Development of a TaqMan Real-Time PCR for Early and Accurate Detection of Anthracnose Pathogen Colletotrichum siamense in Pachira glabra. Plants. 2024; 13(8):1149. https://doi.org/10.3390/plants13081149
Chicago/Turabian StyleGu, Jiaqi, Haihua Wang, Xiaoyan Huang, Limei Liao, Huan Xie, and Xixu Peng. 2024. "Development of a TaqMan Real-Time PCR for Early and Accurate Detection of Anthracnose Pathogen Colletotrichum siamense in Pachira glabra" Plants 13, no. 8: 1149. https://doi.org/10.3390/plants13081149