
Early Drought Stress Warning in Plants: Color Pictures of Photosystem II Photochemistry



1
Department of Botany, Aristotle University of Thessaloniki, 54124 Thessaloniki, Greece
2
Institute of Plant Breeding and Genetic Resources, Hellenic Agricultural Organisation–Demeter (ELGO-Demeter), Thermi, 57001 Thessaloniki, Greece
*
Author to whom correspondence should be addressed.
†
Current address: Department of Plant and Environmental Sciences, University of Copenhagen, Thorvaldsensvej 40, 1871 Frederiksberg C, Denmark.
Climate 2022, 10(11), 179; https://doi.org/10.3390/cli10110179
Submission received: 16 October 2022
/
Revised: 12 November 2022
/
Accepted: 16 November 2022
/
Published: 18 November 2022
(This article belongs to the Special Issue Drought Early Warning)
Abstract
:Drought, the major limiting factor for plant growth and crop productivity, affecting several physiological and biochemical processes, is expected to increase in duration, intensity, and frequency as a consequence of climate change. Plants have developed several approaches to either avoid or tolerate water deficit. Plants as a response to drought stress (DS), close stomata, reducing carbon dioxide (CO2) entry in the leaf, thus decreasing photosynthesis which results in reduced synthesis of essential organic molecules that sustain the life on earth. The reduced CO2 fixation, decreases electron transport rate (ETR), while the absorbed light energy overdoes what can be used for photochemistry resulting in excess reactive oxygen species (ROS) and oxidative stress. Current imaging techniques allow non-destructive monitoring of changes in the physiological state of plants under DS. Thermographic visualization, near-infrared imaging, and chlorophyll a fluorescence imaging are the most common verified imaging techniques for detecting stress-related changes in the display of light emission from plant leaves. Chlorophyll a fluorescence analysis, by use of the pulse amplitude modulation (PAM) method, can principally calculate the amount of absorbed light energy that is directed for photochemistry in photosystem II (PSII) (ΦPSII), dissipated as heat (ΦNPQ), or dissipated by the non-radiative fluorescence processes (ΦNO). The method of chlorophyll a fluorescence imaging analysis by providing colour pictures of the whole leaf PSII photochemistry, can successfully identify the early drought stress warning signals. Its implementation allowed visualization of the leaf spatial photosynthetic heterogeneity and discrimination between mild drought stress (MiDS), moderate drought stress (MoDS), and severe drought stress (SDS). The fraction of open reaction centers of PSII (qp) is suggested as the most sensitive and suitable indicator of an early drought stress warning and also for selecting drought tolerant cultivars.
1. Introduction
Plant growth and development experience a non-stop exposure to biotic and abiotic stresses conditions. Photosynthesis is the device of crop productivity, but besides it is a complicated process that is highly responsive to biotic and abiotic stresses with a complex association to plant growth [1]. In this review article we present the impact of water deficiency on plants and the drought avoidance and drought tolerance mechanisms of plants. We discuss on the drought stress effects on photosynthesis with special emphasis on the light reactions of photosynthesis and the production of reactive oxygen species in photosystems I (PSI) and II (PSII). We present a theoretical background of the method of chlorophyll fluorescence analysis that is used for the evaluation of photosynthetic function under abiotic and biotic stress conditions and we mention some other imaging techniques that allow a non-destructive monitoring of changes in the physiological state of plants under drought stress. As an application example of the method of chlorophyll fluorescence imaging analysis in drought stress phenotyping, we selected the model plant Arabidopsis thaliana as the most suitable plant species, and we provide colour pictures of the whole leaf PSII photochemistry that can be used successfully as early drought stress warning signals.
Drought is one of the major limiting factors for plant growth and crop productivity because it affects several physiological and biochemical processes [2,3,4,5] and is expected to increase in duration, intensity, and frequency as a consequence of climate change [6,7,8,9,10], reaching to alarm level by now days [11,12,13]. Drought stress (DS) is the main issue amongst all environmental situations associated with the forecast effects of climate change that will detrimentally influence global crop production [14,15]. Water deficit impairs plant’s cell division, elongation and differentiation, osmotic adjustment, causing loss of turgor, and harms photosynthetic rates and growth, disturbing energy balance, and eventually decreases plant productivity [16,17,18].
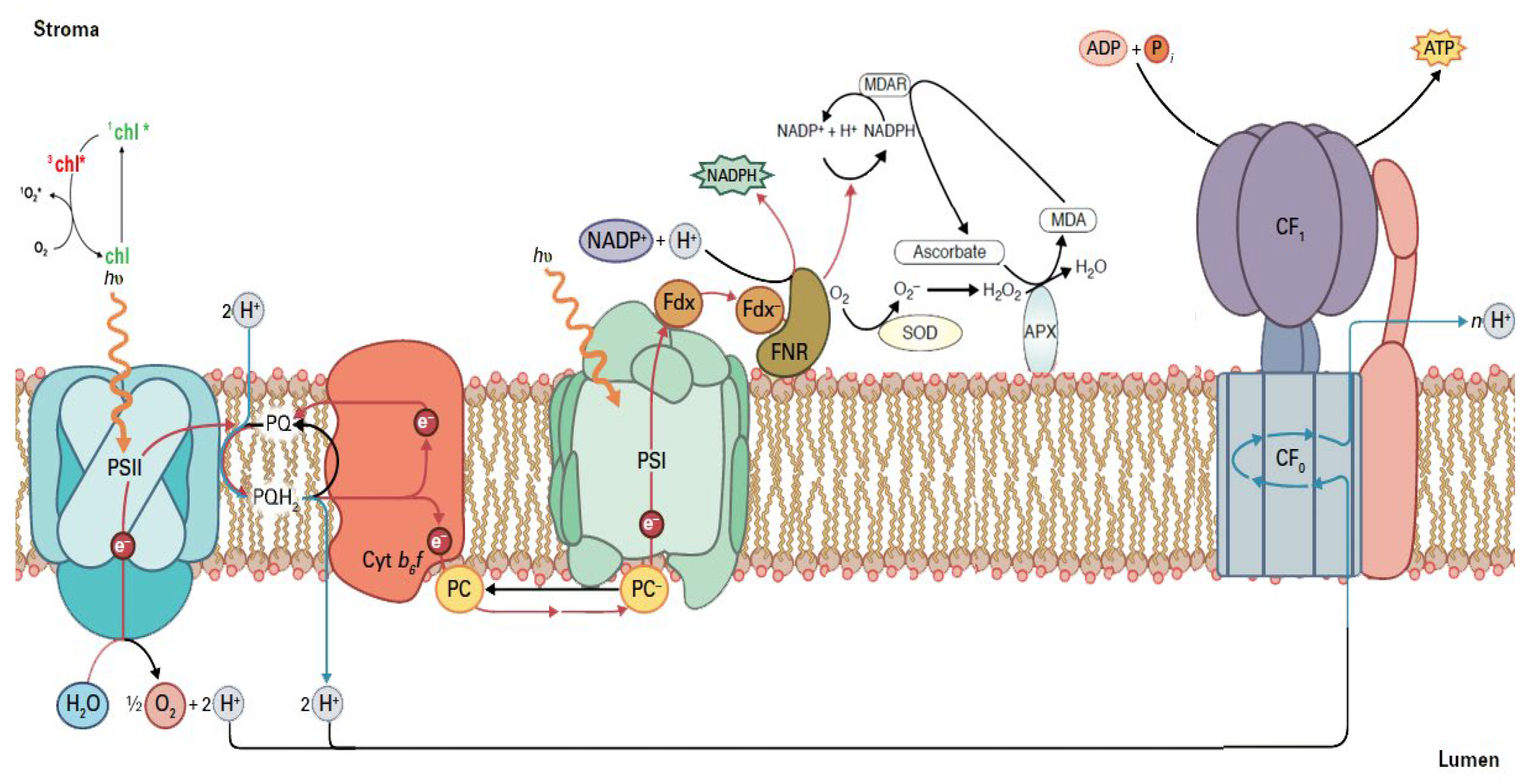
As a response to DS, plants close stomata to reduce water loss (transpiration) in order to prevent dehydration, but this results in limiting CO2 to penetrate the leaf, thus affecting detrimentally photosynthesis [2,3]. Thus, stomatal aperture must be strictly regulated to play a dual role, preventing dehydration while sustaining photosynthesis [2,3]. Photosynthesis is fundamental to plant growth, functioning and fitness, however the plant’s capability to acquire and retain highest photosynthetic capacity greatly relies upon the environmental conditions [19,20]. Photosynthesis of food crops under DS has been considered to be a real challenge for plant scientists and crop breeders in order to fulfill the huge demand for food in the world [21,22]. Photosynthetic capacity and plant productivity are frequently superior in environmental conditions with higher water availability since H2O accomplishes an essential role in photosynthesis [3,20]. The oxidation of H2O molecules in Photosystem II (PSII), that uses the light energy, provides protons (H+), and electrons (e−) that result in the formation of ATP and reducing power (reduced ferredoxin and NADPH) (Figure 1), for the synthesis of essential organic molecules that sustain the life on Earth [23,24,25]. PSII supramolecular complex consists of a water-splitting system (oxygen evolving complex, OEC), a light-harvesting chlorophyll protein complex (LHCII) and a reaction center (RC).
Light-harvesting complex of PSII (LHCII) absorbs light energy and transfers it to the RC of PSII. When there is an excess light energy, creation of singlet oxygen (1O2) via the triplet state of chlorophyll (3chl*) occurs [27] (Figure 1). At PSII the oxidation of water, at the water-splitting complex, results to O2, H+, and e− (Figure 1). The e− are transferred from H2O to NADP+, while accompanying this e− transfer, a proton gradient is established across the membrane that is utilized for the synthesis of ATP by the ATP synthase [28]. At the RC of PSII the excitation of specially bound chlorophyll molecules results in transfer of an e− from H2O oxidation to quinone A (QA). The fully reduced quinol molecule (PQH2) picks up two H+ from the stroma and is oxidized to a quinone (PQ) and while the e− are transferred through cytochrome b6f, to plastocyanin (PC) and to PSI, H+ are transferred from the stroma to the chloroplast lumen [28] (Figure 1). The structures of the soluble proteins ferredoxin (Fdx) and ferredoxin-NADP+ reductase (FNR), on the stromal side, that transfer the e− to NADP+ to form NADPH, are also shown in Figure 1. As a result of DS, stomatal closure, limits CO2 entry to the leaf, and since NADPH is not used in Calvin–Benson–Bassham cycle, NADP+ is not available. Under such circumstances, e− are transferred to molecular oxygen (O2) forming superoxide anions (O2•−) that are converted by the superoxide dismutase (SOD) to hydrogen peroxide (H2O2) that is reduced by ascorbate peroxidase (APX) to H2O and O2 [29,30] (Figure 1). Ascorbate peroxidase uses electrons from ascorbate (AsA) that is oxidized, but through monodehydroascorbate reductase (MDAR), AsA is reduced from NADPH, contributing to NADP+ availability [29] (Figure 1).
2. Drought Stress Impact on Plants
Plant metabolism is accomplished with water involvement, and adequate water is an important circumstance for growth and development [31]. Drought stress that negatively impacts plant growth is the major constraint to crop production and a growing concern for crop yields as a result of global climate changes that involve increased DS periods [32,33]. Plants are subjected to DS conditions when either the water supply to the roots is restricted or the water loss through transpiration is extreme [34]. Thus, the imbalance from a reduced water uptake with an excessive water loss, and the oxidative damage stimulated by the increased ROS generation, that are induced by water deficiency, result in noteworthy changes in plant growth, biomass production, photosynthesis, and enzymatic activities [34,35,36,37,38,39,40].
Drought stress impairs osmotic adjustment of plants and harms photosynthesis and growth [17,41,42], resulting in reduced crop yields that affect food security worldwide [7,43,44], with up to 21 and 40% yield reductions in wheat (Triticum aestivum L.) and maize (Zea mays L.), respectively [45]. Stomatal closure, as a response to water deficit, reduces excess water loss (transpiration) to prevent desiccation, but also limits CO2 to penetrate the leaf, thus harmfully affecting photosynthesis [46]. Due to this compromise, stomatal aperture must be strictly coordinated [33,47].
The unfavorable conditions of climate change are contributing to development of extended water deficit areas and consequently on the plant growth and crop productivity. Although a positive impact of elevated CO2 on crop yield by increasing photosynthesis is suggested by some researchers, it is debated by some others showing that increased CO2 does not counteract the effect of severe drought on photosynthesis and yield [46,48].
3. Plant Tolerance to Drought Stress
Plants have developed several energetic approaches at the morphological, physiological and biochemical levels, permitting them to avoid and/or tolerate water deficit [9,42,46,49,50]. Avoidance mechanisms are mainly morphological and physiological adjustments that provide an escape to the water deficit, e.g., by increased root system, increased leaf thickness, decreased leaf area, reduced stomatal number and conductance, and leaf rolling or folding to minimize evapotranspiration [46,51,52]. Drought tolerance traits are correlated with maintenance of the plant water status throughout osmotic adjustment by the accumulation of osmoprotective substances, for example, proline, glycine betaine and sugars, that help the plants to preserve their water status [41,42,53,54,55,56], and to acclimate to water deficit [9,42,57].
Drought stress can develop in a wide range, from mild drought stress (MiDS), to moderate drought stress (MoDS), and to severe drought stress (SDS), thru which plants experience dehydration and wilting, which ultimately leads to death [9,58]. Numerous studies have focused on SDS, although, MoDS is developing more frequently in actual circumstances [58]. Current studies have confirmed that plants employ diverse approaches to manage MiDS, compared to MoDS or to SDS [9,58,59,60,61,62,63]. For instance, in Arabidopsis young leaves photosynthetic efficiency is recovering under MoDS, but not in MiDS [38,64], while plants that were defined to be tolerant to SDS do not withstand MiDS [61].
4. Photosynthetic Function under Drought Stress
Drought stress significantly decreases photosynthetic activity and disrupts plant productivity [65]. The decreased photosynthetic activity is linked to both stomatal and non-stomatal effects, which are not totally understood [55,65,66,67,68]. Drought stress decreases photosynthesis by reducing carbon dioxide availability through increased resistance to carbon dioxide flow from stomata, disrupts either biochemical or/and photochemical activity and increases leaf membrane lipid peroxidation [9,37,38,69,70,71,72]. The diminished CO2 fixation under DS results in decreased electron transport rate (ETR) [73,74], while the absorbed light energy that overdoes what it can be used for photochemistry results in surplus reactive oxygen species (ROS) accumulation, that can harm the chloroplast, and particularly damage photosystem II (PSII) [9,24,75,76,77]. However, damage of PSII can be prevented by dissipation of excess light energy as heat, a process termed non-photochemical quenching (NPQ), and typically estimated by chlorophyll a fluorescence analysis [9,78,79,80]. The NPQ mechanism is considered to be the principal photoprotective mechanism and is more efficient in the combined existence of the PsbS (PSII subunit S) protein, and zeaxanthin of the xanthophyll cycle [81,82]. NPQ by dissipating harmless the excess absorbed light energy under DS, decreases ETR to prevent ROS formation, thus it can regulate to a level ROS formation [83,84,85]. Our understanding of the NPQ process has been advanced by use of the pulse amplitude modulation (PAM) fluorescence analysis that it is quantifying NPQ photoprotective potential in addition to the classical chlorophyll fluorescence induction analysis [86].
5. Reactive Oxygen Species Generation under Drought Stress
ROS, such as superoxide anion radical (O2•−), hydrogen peroxide (H2O2), and singlet oxygen (1O2), are continuously produced at basal levels, mainly in the light reactions of photosynthesis, but are kept in a homeostasis by the antioxidative enzymatic and non-enzymatic systems [17,30,79,87]. Drought stress breakdown the balance between the creation and elimination of ROS in plants [9]. Thus, during water deficit periods ROS creation rises extremely [88], and this triggers oxidative stress causing membrane injuries, protein degradation and enzyme inactivation that damage the cellular components [40,89,90]. To prevent oxidative damage, beside the NPQ mechanism that is considered as the principal photoprotective mechanism, plants have an effective antioxidant defence system, with both enzymatic and nonenzymatic systems [40,91,92]. Efficient enzymatic antioxidants, such as SOD, APX, MDAR, glutathione reductase (GR), glutathione peroxidase (GPX), guaiacol peroxidase (GOPX), and catalase (CAT), and the non-enzymatic metabolites, such as AsA, glutathione (GSH), a-tocopherol, carotenoids, phenolic compounds, flavonoids, and proline [93,94,95], play critical roles in removing the water deficiency-induced excessive ROS [96]. The ascorbate-glutathione (AsA-GSH) cycle is a crucial component of the enzymatic antioxidant defence system in plants [97,98].
The most reactive of all ROS is the hydroxyl radical (OH•) that reacts with almost all molecules but it is the shortest lived. The e− leakage to O2 at PSI results in O2•− which is rapidly converted by SOD to H2O2 that is longer lived than O2•−, which is shorter lived than 1O2 but longer lived than OH•. Hydrogen peroxide is the most stable and least reactive ROS with the longest lifetime, and is being able to easily diffuse through the membranes [29,30,99,100,101].
ROS production in the process of light absorption and energy use in photosynthesis confers an important biological function (plant growth and development, redox signaling) besides generating oxidative stress [99,100,101,102]. The role of antioxidants (enzymic and non-enzymic) in the photosynthetic apparatus is not to totally remove ROS, but rather to accomplish an appropriate equilibrium among production and scavenging so as to pair the process of photosynthesis, permitting an efficient spreading of signals to the nucleus [103,104,105].
ROS function is not only to monitor electron transport and, consequently, prevent over-reduction or over-oxidation, but also generate redox regulatory networks that allow plants to sense and react to environmental stress conditions [88,104,106,107]. ROS trigger plant’s protection mechanisms in order to manage oxidative stress, and are now considered important signaling molecules for the regulation of plethora physiological functions and the acclimation response [65,85,87,88,98,107,108,109,110,111].
Under DS, an excess accumulation of ROS can damage the chloroplast, with PSII being especially exposed to damage [9,24,65,77]. Although ROS were primarily considered to be toxic by-products it is now recognized that a basal level of ROS is fundamental to sustain life [99,112,113]. A basal level of ROS is desirable for optimal plant growth, while a little amplified level of ROS is beneficial for triggering stress defense responses, but a high level of ROS out of the limits is considered harmful to plants [25,103]. Nowadays, the consequences of global climate change request a better understanding of the relationship between PSII photochemistry and ROS role as a molecule for photoprotection [101]. The illumination of this interaction could assist to enhance agricultural sustainability under a global climate change [101].
6. Plant Phenotyping for Drought Stress Tolerance
Climate change quickly turns-off into a climate crisis with huge worries for agricultural production. Since water deficit is one of the key hazards for the future of agriculture and the total worldwide population, assessing and investigating the ability of crops to grow with restricted water is therefore essential [65,114]. In recent years, much effort has been made to study plant responses to drought in order to address the present and future risks associated with climate change [65,115,116]. The availability of diverse imaging techniques has allowed real-time imaging analysis of physiological changes in plants under DS for high-throughput screening [117,118,119]. The development of modern analytical techniques has created huge inputs to high-throughput plant phenotyping providing various information related to plant status [120]. Selecting the proper imaging sensors is fundamental in designing phenotyping facilities, which depend on the special experimental objects [121]. These techniques allow the pre-symptomatic monitoring of plant stress, a long time before any visible symptoms developed, enabling for high-throughput screening [117]. By the time that visible symptoms of stress are displayed, the plant could have been already adversely affected [117]. Current imaging techniques allow non-destructive monitoring of changes in the physiological state of plants under DS [37,120,122]. Thermal imaging (also called far-infrared thermal imaging) that determines energy loss from stomatal aperture by the leaf temperature, hyperspectral imaging (visible and near-infrared) that can provide spatial information simultaneously, and chlorophyll a fluorescence imaging are the most common verified imaging techniques for detecting stress-related changes from plant leaves [117,121,122,123,124,125,126,127,128]. Crop monitoring using imaging techniques would allow us to relieve stress at an early stage, avoiding permanent damage and thus considerably decreasing yield losses [117,120,129].
Chlorophyll a fluorescence imaging uses blue and red fluorescence to detect the emission that results from absorbed light energy which is not dissipated as heat, or it is not used for the photosynthetic reactions in photochemistry [122]. The chlorophyll fluorescence parameters that are measured can be decoded in terms of photosynthetic activity to acquire knowledge about photosynthetic function and particularly of PSII [130,131]. The data of chlorophyll a fluorescence measurements have been considerably used to probe the function of the photosynthetic apparatus and for screening different crops for plant tolerance to numerous stresses [122,132,133,134,135,136,137,138,139,140,141,142,143,144]. Chlorophyll fluorescence imaging instruments offer the option to evaluate photosynthetic function at the whole leaf surface and identify leaf spatial heterogeneity [122,145,146]. Photosynthetic performance is extremely heterogeneous at the leaf surface, especially under stress conditions [145,147,148,149,150,151,152,153]. Chlorophyll fluorescence analysis is a quick, easy, non-invasive, cheap, and highly sensitive method that can determine photosynthetic function accurately and sense the impact of different stresses on plants [122,151,154].
7. Theoretical Aspects of Chlorophyll Fluorescence Analysis
Chlorophyll a fluorescence analysis, with the use of pulse amplitude modulation (PAM) method can principally calculate the amount of absorbed light energy that is directed to PSII for photochemistry, dissipated as heat through the non-photochemical quenching (NPQ) mechanism, or it is dissipated by the less well characterized non-radiative fluorescence processes, that are marked as ΦPSII, ΦNPQ, and ΦNO, respectively, with the sum of them to be equal to one [122,155,156].
Chlorophyll fluorescence quenching analysis using the PAM method is based on the principle that the leaf has to be in a dark-adapted state before the measurements, so as the first stable acceptor of PSII, QA, is fully oxidized. This can be achieved by dark incubation for several minutes [154]. In the dark-adapted state a “measuring light” is switched on that is high enough to produce the minimal level of chlorophyll fluorescence, termed Fo (Figure 2). A short-lived saturating pulse of light results in the formation of the maximum yield of fluorescence, Fm. By subtracting Fo from Fm the variable fluorescence, Fv results. The ratio Fv/Fm is indicator of the maximum quantum yield of PSII photochemistry. The application of saturating pulses under actinic light illumination closes all the reaction centers and provides the maximum fluorescence in the light-adapted state, that is termed Fm′. The steady-state level of fluorescence in the light is the Fs and is calculated before switching off the actinic light. Fo′ is measured directly after switching off the actinic light. By subtracting Fo′ from Fm′ the variable fluorescence, Fv′ results. The ratio Fv′/Fm′ is indicative of the efficiency of excitation energy capture by open PSII reaction centers.
Since photosynthetic performance is not homogeneous at the leaf surface, especially under DS conditions, it renders conventional chlorophyll fluorescence analysis non-representative of the physiological status of the whole leaf [145,149,150]. The manufactured new chlorophyll fluorescence imaging instruments that are capable of analysing the spatial photosynthetic heterogeneity at the whole leaf surface offer new potentials that cannot be obtained by conventional chlorophyll fluorescence analysis [126,127,138,145]. A list of chlorophyll a fluorescence parameters used in studies of photosystem II photochemistry, and in this review, together with their definitions, are given in Table 1.
8. Chlorophyll Fluorescence Analysis for Drought Stress Tolerance
Chloroplasts throughout the procedure of photosynthesis play an essential role as redox sensors of DS conditions and stimulate acclimatory or stress defense responses [38,157,158,159,160]. The redox state of QA has a critical influence on plant growth, development, and defence [161], and is considered as a sensor of the energy imbalance under any stress conditions [162,163]. If the excess energy can not be dissipated under stress conditions, over-reduction of the photosynthetic electron transport chain (ETC) occurs [156]. Over-reduction of the ETC can severely damage the chloroplast and the cell [164]. Excess excitation energy and consequently an imbalance between energy supply and demand outcomes in increased ROS production [84,105,110], that causes damage to proteins, lipids, and nucleic acids [156,165,166]. Photoinhibition is a product of this damage, and PSII is the primary photoinhibition target [167]. Photoinhibition reduces the number of active PSII centers and is widespread across species, light situations and habitats [168,169,170]. It can be estimated by chlorophyll a fluorescence analysis, based on the ratio Fv/Fm, the maximum efficiency of PSII photochemistry [167,169].
The redox state of QA, as estimated by the parameter 1 − qL, is representing the fraction of PSII reaction centers in closed state (based on a “lake” model for the photosynthetic unit) [155]. Changes in the redox state of QA are considered to act as a signal to the stomatal guard cells [65,163]. A more oxidized QA pool under DS conditions matches to the lowest stomatal opening and it is linearly correlated to stomatal conductance [65,171], that is a measure of stomatal closure and is commonly used as a water stress index [172,173]. It is now broadly accepted that the redox signals are important regulators of plant metabolism and also death [174,175].
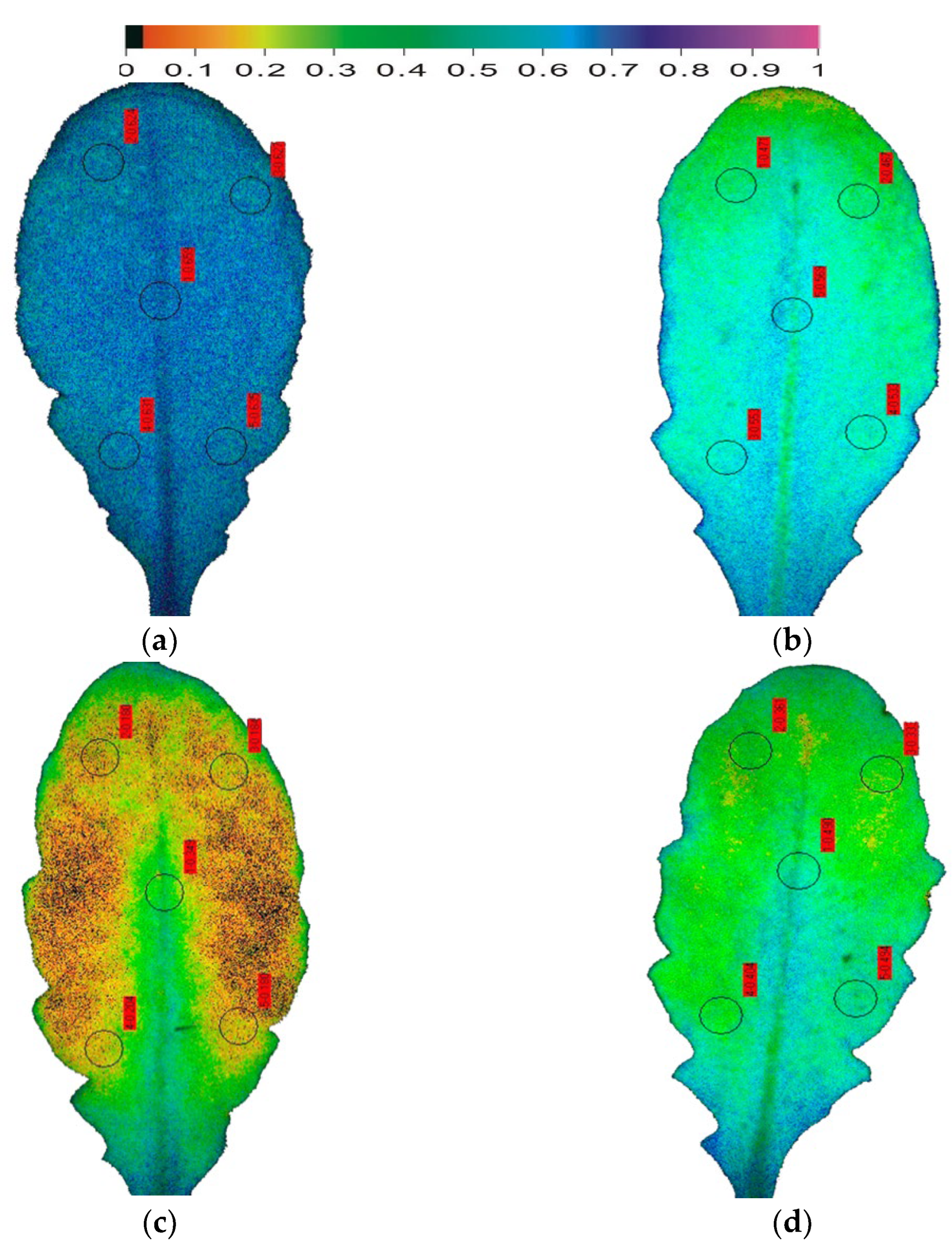
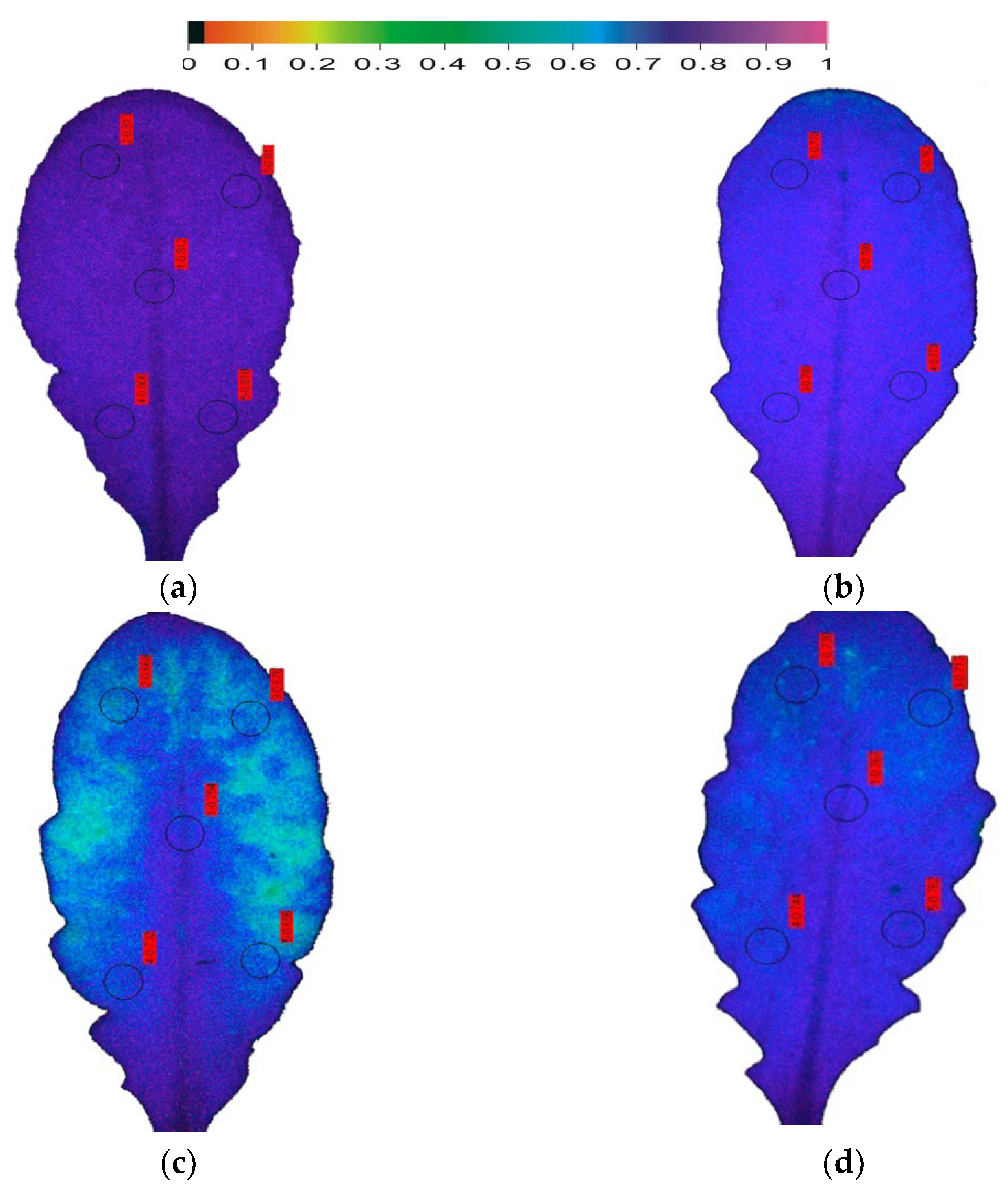
Using the model plant Arabidopsis thaliana, that is considered as the most suitable for the application of the method of chlorophyll fluorescence imaging analysis [37], the early DS responses of photosynthesis were assessed (Figure 3 and Figure 4). Between the chlorophyll fluorescence parameters that have been used for evaluation, monitoring, and selection of drought-tolerant plants, the maximum efficiency of PSII photochemistry (Fv/Fm) was the one that was used most [72]. The photosynthetic efficiency of Arabidopsis thaliana plants, whose watering stopped (i) twenty-four hours before sampling, characterized as being at the onset of drought stress (OnDS); (ii) six days before sampling, characterized as at mild drought stress (MiDS); and (iii) ten days before sampling, characterized as at moderate drought stress (MoDS) [37], was evaluated by chlorophyll fluorescence imaging analysis. The maximum efficiency of PSII photochemistry (Fv/Fm) decreased as soon as 24 h after the onset of drought stress (OnDS) (Figure 3b), while under MiDS decreased further (Figure 3c). However, under further water deficit treatment (ten days, MoDS), the maximum efficiency of PSII photochemistry recovered (Figure 3d). Further DS treatment, characterized as severe drought stress (SDS), resulted in a significant diminished PSII photochemistry [37]. Most authors did not find significant decreases in Fv/Fm under MoDS [3,176,177], indicating that ETR is unaltered under MoDS [178]. The reduction status of the plastoquinone pool (qp) in Arabidopsis thaliana decreased as soon as 24 h after the onset of drought stress (OnDS) (Figure 4b) by 18%, while under MiDS by 66% compared to well-watered Arabidopsis plants (Figure 4c). However, under MoDS qp was 34% lower than well-watered Arabidopsis (Figure 4d). Photosystem II reaction centers are supposed to be open (qp = 1) or closed (qp = 0) depending upon whether they are ready to accept light energy from antennas to excite an electron (open), or unable to accept light energy (closed). Based on the abovementioned data the fraction of open reaction centers of PSII (qp), is a more sensitive parameter to probe DS effects compared to Fv/Fm. Recent studies proposed that the fraction of open reaction centers of photosystem II (PSII) (qp), or in other words the reduction status of the plastoquinone pool, is more sensitive than the Fv/Fm that is traditionally used, and thus qp is a more proper indicator to probe the effects of biotic or abiotic stresses on leaf photosynthesis [9,105,179], and to select drought tolerant cultivars under deficit irrigation [65].
The resulting increase in leaf spatial heterogeneity from well-watered (control plants) to MiDS is reflected in the decrease of the fraction of open reaction centers (qp) integrated over the leaf (Figure 4). The increase of the fraction of open reaction centers (qp) under MoDS retained the leaf spatial heterogeneity. After exposure to DS, qp values decreased differentially in different parts of the leaf. Under water deficit treatments qp values were higher in the proximal (leaf base) compared to the distal (leaf tip) part (Figure 4). Under all DS treatments, Fv/Fm decreased less in the leaf base than in the leaf tip (Figure 3). Decline of the Fv/Fm ratio has been related to a decline in the ability of PSII to reduce the primary electron acceptor, QA [180].
The spatiotemporal heterogeneity observed in A. thaliana leaves under DS implies that pigment concentration and composition, water potential and stomatal function unquestionably fluctuate in different cell regions of the leaf, contributing to spatial differentiations in photochemical activity [181,182]. This spatial photosynthetic heterogeneity under DS may reflect diverse zones of leaf anatomy and mesophyll development [37]. Blade maturation of Arabidopsis leaves appears from the tip to the base of the leaf [37], the latter representing younger cells in leaf anatomy [183], with not fully developed chloroplasts [184].
While A. thaliana does not seem to activate any tolerance mechanism under MiDS, it appears not to suffer under MoDS [37]. This tolerance mechanism under MoDS was suggested to be activated by an antioxidant defense mechanism that activated ROS scavenging [9]. An early drought warning system is much more than a forecast for decision making in response to a changing climate [185,186].
9. Conclusions
The method of chlorophyll fluorescence analysis has been widely used to monitor biotic and abiotic stress effects on plants by using leaf photosynthesis attributes [130,135,137,154,187]. Recently, besides the above-ground parts of crops, a phenotyping method focused on the rooting system and its activity was proposed [188]. The color pictures of PSII photochemistry that can be obtained by chlorophyll fluorescence imaging analysis [37,122] are suitable to characterize and differentiate plant tolerance to DS by evaluating their photosynthetic efficiency and produce also useful information that can be used effectively to phenotype plants under water deficit, with the aim to identify the optimum irrigation conditions. The method of chlorophyll a fluorescence imaging analysis by providing colour pictures of the whole leaf PSII photochemistry, can successfully identify the early DS warning signals allowing the pre-symptomatic monitoring of DS in a non-destructive way. It is an easy, quick, cheap, non-invasive, and highly sensitive method [122,187]. Its implementation allowed visualization of the leaf spatial photosynthetic heterogeneity and discrimination between mild drought stress (MiDS), moderate drought stress (MoDS), and severe drought stress (SDS) [37]. The reduction status of the plastoquinone pool or in other words the fraction of open reaction centers of PSII (qp) has been shown to be the most sensitive and suitable indicator to probe photosynthetic function and determine the impact of biotic and abiotic stresses on plants [37,105,189], and also to select drought tolerant cultivars under deficit irrigation [65].
Author Contributions
Conceptualization, M.M.; software, J.M.; validation, M.M., I.S. and J.M.; formal analysis, M.M., I.S. and J.M.; data curation, M.M., I.S. and J.M.; writing—original draft preparation, M.M.; writing—review and editing, M.M., I.S. and J.M.; visualization, M.M.; supervision, M.M.; project administration, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harbinson, J.; Yin, X. Modelling the impact of improved photosynthetic properties on crop performance in Europe. Food Energy Secur. 2022. ahead of print. [Google Scholar] [CrossRef]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Flexas, J.; Barón, M.; Bota, J.; Ducruet, J.-M.; Gallé, A.; Galmés, J.; Jiménez, M.; Pou, A.; Ribas-Carbó, M.; Sajnani, C. Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri × V. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Cao, J.; Chen, Q.; He, J.; Liu, Z.; Wang, J.; Li, X.; Yang, Y. The kinase CIPK11 functions as a negative regulator in drought stress response in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros Junior, U.O.; Lima, M.D.R.; Alsahli, A.A.; Lobato, A.K.S. Unraveling the roles of brassinosteroids in alleviating drought stress in young Eucalyptus urophylla plants: Implications on redox homeostasis and photosynthetic apparatus. Physiol. Plant. 2021, 172, 748–761. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Change 2012, 3, 52–58. [Google Scholar] [CrossRef]
- Urban, L.; Aarrouf, J.; Bidel, L.P.R. Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front. Plant Sci. 2017, 8, 2068. [Google Scholar] [CrossRef] [Green Version]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; DiDio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of drought stress research: Experimental setup and physiological characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [Green Version]
- Sperdouli, I.; Moustaka, J.; Ouzounidou, G.; Moustakas, M. Leaf age-dependent photosystem II photochemistry and oxidative stress responses to drought stress in Arabidopsis thaliana are modulated by flavonoid accumulation. Molecules 2021, 26, 4157. [Google Scholar] [CrossRef]
- Aubert, L.; Konrádová, D.; Barris, S.; Quinet, M. Different drought resistance mechanisms between two buckwheat species Fagopyrum esculentum and Fagopyrum tataricum. Physiol Plant. 2021, 172, 577–586. [Google Scholar] [CrossRef]
- Walter, J.; Nagy, L.; Hein, R.; Rascher, U.; Beierkuhnlein, C.; Willner, E.; Jentsch, A. Do plants remember drought? Hints towards a drought-memory in grasses. Environ. Exp. Bot. 2011, 71, 34–40. [Google Scholar] [CrossRef]
- Zhao, T.; Dai, A. The magnitude and causes of global drought changes in the twenty-first century under a low–severe emissions scenario. J. Clim. 2015, 28, 4490–4512. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, C.; Ogaya, R.; Fernández-Martínez, M.; Pugh, T.A.M.; Peñuelas, J. Increasing climatic sensitivity of global grassland vegetation biomass and species diversity correlates with water availability. New Phytol. 2021, 230, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Muktadir, M.A.; Adhikari, K.N.; Ahmad, N.; Merchant, A. Chemical composition and reproductive functionality of contrasting faba bean genotypes in response to water deficit. Physiol. Plant. 2021, 172, 540–551. [Google Scholar] [CrossRef]
- Hsiao, T.C. Plant responses to water stress. Ann. Rev. Plant Physiol. 1973, 24, 519–570. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Zhang, W.; Du, T.; Kang, S.; Davies, W.J. Responses of water accumulation and solute metabolism in tomato fruit to water scarcity and implications for main fruit quality variables. J. Exp. Bot. 2020, 71, 1249–1264. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509531. [Google Scholar] [CrossRef]
- Walczyk, A.M.; Hersch-Green, E.I. Do water and soil nutrient scarcities differentially impact the performance of diploid and tetraploid Solidago gigantea (Giant Goldenrod, Asteraceae)? Plant Biol. 2022. ahead of print. [Google Scholar] [CrossRef]
- Long, S.P.; Zhu, X.G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Ann. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, J. Photosynthetic energy conversion: Natural and artificial. Chem. Soc. Rev. 2009, 38, 185–196. [Google Scholar] [CrossRef]
- Moustaka, J.; Ouzounidou, G.; Sperdouli, I.; Moustakas, M. Photosystem II is more sensitive than photosystem I to Al3+ induced phytotoxicity. Materials 2018, 11, 1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustakas, M.; Moustaka, J.; Sperdouli, I. Hormesis in photosystem II: A mechanistic approach. Curr. Opin. Toxicol. 2022, 29, 57–64. [Google Scholar] [CrossRef]
- Moustakas, M.; Bayçu, G.; Sperdouli, I.; Eroğlu, H.; Eleftheriou, E.P. Arbuscular mycorrhizal symbiosis enhances photosynthesis in the medicinal herb Salvia fruticosa by improving photosystem II photochemistry. Plants 2020, 9, 962. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Li, X.P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant. Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Niyogi, K.K.; Wolosiuk, R.A.; Malkin, R. Photosynthesis. In Biochemistry & Molecular Biology of Plants, 2nd ed.; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; John Wiley & Sons: West Sussex, UK, 2015; pp. 508–566. [Google Scholar]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Moustaka, J.; Tanou, G.; Adamakis, I.D.; Eleftheriou, E.P.; Moustakas, M. Leaf age dependent photoprotective and antioxidative mechanisms to paraquat-induced oxidative stress in Arabidopsis thaliana. Int. J. Mol. Sci. 2015, 16, 13989–14006. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postiglione, A.E.; Muday, G.K. Abscisic acid drives stomatal closure through increases in hydrogen peroxide in distinct subcellular compartments including mitochondria. bioRxiv 2022. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Amrutha, S.; Parveen, A.; Muthupandi, M.; Sivakumar, V.; Dasgupta, M.G. Variation in morpho-physiological, biochemical and molecular responses of two Eucalyptus species under short-term water stress. Acta Bot. Croat. 2019, 78, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Sperdouli, I.; Moustakas, M. Differential response of photosystem II photochemistry in young and mature leaves of Arabidopsis thaliana to the onset of drought stress. Acta Physiol. Plant. 2012, 34, 1267–1276. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Spatio-temporal heterogeneity in Arabidopsis thaliana leaves under drought stress. Plant Biol. 2012, 14, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Moustakas, M. A better energy allocation of absorbed light in photosystem II and less photooxidative damage contribute to acclimation of Arabidopsis thaliana young leaves to water deficit. J. Plant Physiol. 2014, 171, 587–593. [Google Scholar] [CrossRef]
- Cotado, A.; Munne-Bosch, S.; Pinto-Marijuan, M. Strategies for severe drought survival and recovery in a Pyrenean relict species. Physiol. Plant. 2020, 169, 276–290. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.Y.; Zhang, N.N.; Yao, B.H.; Zhang, X.; Liu, W.Y.; Zhang, W.Q.; Zhang, J.H.; Wei, G.H.; Chen, J. Interactions between hydrogen sulphide and rhizobia modulate the physiological and metabolism process during water deficiency-induced oxidative defense in soybean. Plant Cell Environ. 2022. ahead of print. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsis thaliana to drought stress. J. Plant Physiol. 2012, 169, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Hanjra, M.A.; Qureshi, M.E. Global water crisis and future food security in an era of climate change. Food Policy 2010, 35, 365–377. [Google Scholar] [CrossRef]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Nilson, S.E.; Assmann, S.M. The control of transpiration. Insights from Arabidopsis. Plant Physiol. 2007, 143, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.B.; Dermody, O.; Klein, S.P.; Locke, A.M.; McGrath, J.M.; Paul, R.E.; Rosenthal, D.M.; Ruiz-Vera, U.M.; Siebers, R.; Ainsworth, E.A. Intensifying drought eliminates the expected benefits of elevated carbon dioxide for soybean. Nat. Plants 2016, 2, 16132. [Google Scholar] [CrossRef]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, Y.; Zhang, G.; Gan, Z.; Gao, M.; Lv, J.; Wu, T.; Zhang, X.; Xu, X.; Yang, S.; et al. Group-C/S1 bZIP heterodimers regulate MdIPT5b to negatively modulate drought tolerance in apple species. Plant J. 2021, 107, 399–417. [Google Scholar] [CrossRef]
- Sicher, R.C.; Timlin, D.; Bailey, B. Responses of growth and primary metabolism of water-stressed barley roots to rehydration. J. Plant Physiol. 2012, 169, 686–695. [Google Scholar] [CrossRef]
- Goufo, P.; Moutinho-Pereira, J.M.; Jorge, T.F.; Correia, C.M.; Oliveira, M.R.; Rosa, E.A.S.; António, C.; Trindade, H. Cowpea (Vigna unguiculata L. Walp.) metabolomics: Osmoprotection as a physiological strategy for drought stress resistance and improved yield. Front. Plant Sci. 2017, 8, 586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Sperdouli, I.; Kouna, T.; Antonopoulou, C.I.; Therios, I. Exogenous proline induces soluble sugar accumulation and alleviates drought stress effects on photosystem II functioning of Arabidopsis thaliana leaves. Plant Growth Regul. 2011, 65, 315–325. [Google Scholar] [CrossRef]
- Bano, H.; Athar, H.R.; Zafar, Z.U.; Ogbaga, C.C.; Ashraf, M. Peroxidase activity and operation of photo-protective component of NPQ play key roles in drought tolerance of mung bean [Vigna radiata (L.) Wilcziek]. Physiol. Plant. 2021, 172, 603–614. [Google Scholar] [CrossRef]
- Xu, Q.; Liesche, J. Sugar export from Arabidopsis leaves: Actors and regulatory strategies. J. Exp. Bot. 2021, 72, 5275–5284. [Google Scholar] [CrossRef]
- Chen, Y.; Dubois, M.; Vermeersch, M.; Inzé, D.; Vanhaeren, H. Distinct cellular strategies determine sensitivity to mild drought of Arabidopsis natural accessions. Plant Physiol. 2021, 186, 1171–1185. [Google Scholar] [CrossRef]
- Lawlor, D.W. Limitation to photosynthesis in water-stressed leaves: Stomata vs. metabolism and the role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.R.; Pereira, A. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef] [Green Version]
- Skirycz, A.; Vandenbroucke, K.; Clauw, P.; Maleux, K.; De Meyer, B.; Dhondt, S.; Pucci, A.; Gonzalez, N.; Hoeberichts, F.; Tognetti, V.B.; et al. Survival and growth of Arabidopsis plants given limited water are not equal. Nat. Biotechnol. 2011, 29, 212–214. [Google Scholar] [CrossRef]
- Ma, X.; Sukiran, N.L.; Ma, H.; Su, Z. Moderate drought causes dramatic floral transcriptomic reprogramming to ensure successful reproductive development in Arabidopsis. BMC Plant Biol. 2014, 14, 164. [Google Scholar] [CrossRef] [PubMed]
- Clauw, P.; Coppens, F.; De Beuf, K.; Dhondt, S.; Van Daele, T.; Maleux, K.; Storme, V.; Clement, L.; Gonzalez, N.; Inzé, D. Leaf responses to mild drought stress in natural variants of Arabidopsis. Plant Physiol. 2015, 167, 800–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperdouli, I.; Moustakas, M. Leaf developmental stage modulates metabolite accumulation and photosynthesis contributing to acclimation of Arabidopsis thaliana to water deficit. J. Plant Res. 2014, 127, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Mellidou, I.; Moustakas, M. Harnessing chlorophyll fluorescence for phenotyping analysis of wild and cultivated tomato for high photochemical efficiency under water deficit for climate change resilience. Climate 2021, 9, 154. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Trenberth, K.E.; Dai, A.; van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Change 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Baczewska-Dąbrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M.; Wójcik-Jagła, M.; Pawluśkiewicz, B.; Bąba, W.; Brestic, M. Exploration of chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in perennial ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Diaz-Espejo, A.; Galmeî, S.J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell Environ. 2007, 30, 1284–1298. [Google Scholar] [CrossRef]
- Silva, E.N.; Ferreira-Silva, S.L.; de Vasconcelos Fontenele, A.; Ribeiro, R.V.; Viégas, R.A.; Silveira, J.A.G. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. J. Plant Physiol. 2010, 167, 1157–1164. [Google Scholar] [CrossRef]
- Yao, J.; Sun, D.; Cen, H.; Xu, H.; Weng, H.; Yuan, F.; He, Y. Phenotyping of Arabidopsis drought stress response using kinetic chlorophyll fluorescence and multicolor fluorescence imaging. Front. Plant Sci. 2018, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Cornic, G.; Le Gouallec, J.L.; Briantais, J.M.; Hodges, M. Effect of dehydration and high light on photosynthesis of two C3 plants (Phaseolus vulgaris L. and Elatostema repens (Lour.) Hall f.). Planta 1989, 177, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Bączek-Kwinta, R.; Kozieł, A.; Seidler-Łożykowska, K. Are the fluorescence parameters of German chamomile leaves the first indicators of the anthodia yield in drought conditions? Photosynthetica 2011, 49, 87–97. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J. Effects of water stress on photosystem II photochemistry and its thermostability in wheat plants. J. Exp. Bot. 1999, 50, 1199–1206. [Google Scholar] [CrossRef]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta 2007, 1767, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavafer, A.; Mancilla, C. Concepts of photochemical damage of Photosystem II and the role of excessive excitation. J. Photochem. Photobiol. C 2021, 47, 100421. [Google Scholar] [CrossRef]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Moustaka, J.; Moustakas, M. Photoprotective mechanism of the non-target organism Arabidopsis thaliana to paraquat exposure. Pest. Biochem. Physiol. 2014, 111, 1–6. [Google Scholar] [CrossRef]
- Ruban, A.V. Nonphotochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protecting plants from photodamage. Plant. Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [Green Version]
- Welc, R.; Luchowski, R.; Kluczyk, D.; Zubik-Duda, M.; Grudzinski, W.; Maksim, M.; Reszczynska, E.; Sowinski, K.; Mazur, R.; Nosalewicz, A.; et al. Mechanisms shaping the synergism of zeaxanthin and PsbS in photoprotective energy dissipation in the photosynthetic apparatus of plants. Plant J. 2021, 107, 418–433. [Google Scholar] [CrossRef]
- Ruban, A.V.; Wilson, S. The mechanism of non-photochemical quenching in plants: Localisation and driving forces. Plant Cell Physiol. 2021, 62, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Müller-Moulé, P.; Gilmore, A.M.; Niyogi, K.K. PsbS-dependent enhancement of feedback de-excitation protects photosystem II from photoinhibition. Proc. Natl. Acad. Sci. USA 2002, 99, 15222–15227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Stamelou, M.L.; Sperdouli, I.; Pyrri, I.; Adamakis, I.D.S.; Moustakas, M. Hormetic responses of photosystem II in tomato to Botrytis cinerea. Plants 2021, 10, 521. [Google Scholar] [CrossRef]
- Ruban, A.V. Evolution under the sun: Optimizing light harvesting in photosynthesis. J. Exp. Bot. 2015, 66, 7–23. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Peñuelas, J. Photo- and antioxidative protection, and a role for salicylic acid during drought and recovery in field-grown Phillyrea angustifolia plants. Planta 2003, 217, 758–766. [Google Scholar] [CrossRef]
- Hajiboland, R.; Cheraghvareh, L.; Poschenrieder, C. Improvement of drought tolerance in tobacco (Nicotiana rustica L.) plants by silicon. J. Plant Nutr. 2017, 40, 1661–1676. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Ashraf, U.; Kanu, A.S.; Mo, Z.; Hussain, S.; Anjum, S.A.; Khan, I.; Abbas, R.N.; Tang, X. Lead toxicity in rice: Effects, mechanisms, and mitigation strategies-a mini review. Environ. Sci. Pollut. Res. Int. 2015, 22, 18318–18332. [Google Scholar] [CrossRef] [PubMed]
- Moustaka, J.; Tanou, G.; Giannakoula, A.; Panteris, E.; Eleftheriou, E.P.; Moustakas, M. Anthocyanin accumulation in poinsettia leaves and its functional role in photo-oxidative stress. Environ. Exp. Bot. 2020, 175, 104065. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Tian, Z.; Pan, X.; Zhao, X.; Wang, F. Oxidative stress and non-enzymatic antioxidants in leaves of three edible canna cultivars under drought stress. Hortic. Environ. Biotechnol. 2013, 54, 1–8. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Puertas, M.C.; Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Molina-Moya, E.; Sandalio, L.M. An update on redox signals in plant responses to biotic and abiotic stress crosstalk: Insights from cadmium and fungal pathogen interactions. J. Exp. Bot. 2021, 72, 5857–5875. [Google Scholar] [CrossRef]
- Moustakas, M. Plant Photochemistry, Reactive Oxygen Species, and Photoprotection. Photochem 2022, 2, 5–8. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox signaling in plants. Antioxid. Redox Signal. 2013, 18, 2087–2090. [Google Scholar] [CrossRef] [PubMed]
- Adamakis, I.D.S.; Sperdouli, I.; Eleftheriou, E.P.; Moustakas, M. Hydrogen peroxide production by the spot-like mode action of bisphenol A. Front. Plant Sci. 2020, 11, 1196. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Andreadis, S.; Moustaka, J.; Panteris, E.; Tsaballa, A.; Moustakas, M. Changes in light energy utilization in photosystem II and reactive oxygen species generation in potato leaves by the pinworm Tuta absoluta. Molecules 2021, 26, 2984. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Adamakis, I.D.S.; Malea, P.; Sperdouli, I.; Panteris, E.; Kokkinidi, D.; Moustakas, M. Evaluation of the spatiotemporal effects of bisphenol A on the leaves of the seagrass Cymodocea nodosa. J. Hazard. Mater. 2021, 404, 124001. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Adamakis, I.-D.S.; Moustaka, J.; İşgören, S.; Şaş, B. Harnessing the role of foliar applied salicylic acid in decreasing chlorophyll content to reassess photosystem II photoprotection in crop plants. Int. J. Mol. Sci. 2022, 23, 7038. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Sperdouli, I.; Hanć, A.; Dobrikova, A.; Apostolova, E.; Moustakas, M. Rapid hormetic responses of photosystem II photochemistry of clary sage to cadmium exposure. Int. J. Mol. Sci. 2021, 22, 41. [Google Scholar] [CrossRef]
- Moustaka, J.; Panteris, E.; Adamakis, I.D.S.; Tanou, G.; Giannakoula, A.; Eleftheriou, E.P.; Moustakas, M. High anthocyanin accumulation in poinsettia leaves is accompanied by thylakoid membrane unstacking, acting as a photoprotective mechanism, to prevent ROS formation. Environ. Exp. Bot. 2018, 154, 44–55. [Google Scholar] [CrossRef]
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The evolution of reactive oxygen species metabolism. J. Exp. Bot. 2016, 67, 5933–5943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, V.; D’Amelia, V.; Esposito, M.; Amitrano, C.; Carillo, P.; Carputo, D.; Maggio, A. Anthocyanins are key regulators of drought stress tolerance in tobacco. Biology 2021, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Gurrieri, L.; Merico, M.; Trost, P.; Forlani, G.; Sparla, F. Impact of drought on soluble sugars and free proline content in selected Arabidopsis mutants. Biology 2020, 9, 367. [Google Scholar] [CrossRef]
- Chaerle, L.; Van Der Straeten, D. Imaging techniques and the early detection of plant stress. Trends Plant Sci. 2000, 5, 495–501. [Google Scholar] [CrossRef]
- Furbank, R.T.; Tester, M. Phenomics technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhou, L.; Xiao, Q.; Bai, X.; Wu, B.; Wu, N.; Zhao, Y.; Wang, J.; Feng, L. End-to-end fusion of hyperspectral and chlorophyll fluorescence imaging to identify rice stresses. Plant Phenomics 2022, 2022, 9851096. [Google Scholar] [CrossRef]
- Yang, W.; Feng, H.; Zhang, X.; Zhang, J.; Doonan, J.H.; Batchelor, W.D.; Xiong, L.; Yan, J. Crop phenomics and high-throughput phenotyping: Past decades, current challenges, and future perspectives. Mol. Plant 2020, 13, 187–214. [Google Scholar] [CrossRef]
- Moustakas, M.; Calatayud, A.; Guidi, L. Chlorophyll fluorescence imaging analysis in biotic and abiotic stress. Front. Plant Sci. 2021, 12, 658500. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Filella, I. Visible and near-infrared reflectance techniques for diagnosing plant physiological status. Trends Plant Sci. 1998, 3, 151–156. [Google Scholar] [CrossRef]
- Chaerle, L.; Van Caeneghem, W.; Messens, E.; Lambers, H.; Van Montagu, M.; Van Der Straeten, D. Presymptomatic visualization of plant-virus interactions by thermography. Nat. Biotechnol. 1999, 17, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G. Use of thermography for quantitative studies of spatial and temporal variation of stomatal conductance over leaf surfaces. Plant Cell Environ. 1999, 22, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Chaerle, L.; Van Der Straeten, D. Seeing is believing: Imaging techniques to monitor plant health. Biochim. Biophys. Acta 2001, 1519, 153–166. [Google Scholar] [CrossRef]
- Chaerle, L.; Lenk, S.; Leinonen, I.; Jones, H.G.; Van Der Straeten, D.; Buschmann, C. Multi-sensor plant imaging: Towards the development of a stress-catalogue. Biotechnol. J. 2009, 4, 1152–1167. [Google Scholar] [CrossRef]
- Lowe, A.; Harrison, N.; French, A.P. Hyperspectral image analysis techniques for the detection and classification of the early onset of plant disease and stress. Plant Methods 2017, 13, 80. [Google Scholar] [CrossRef]
- Song, P.; Wang, J.; Guo, X.; Yang, W.; Zhao, C. High-throughput phenotyping: Breaking through the bottleneck in future crop breeding. Crop J. 2021, 9, 633645. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Kalaji, M.H.; Carpentier, R.; Allakhverdiev, S.I.; Bosa, K. Fluorescence parameters as an early indicator of light stress in barley. J. Photochem. Photobiol. B 2012, 112, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Asfi, M.; Ouzounidou, G.; Panajiotidis, S.; Therios, I.; Moustakas, M. Toxicity effects of olive-mill wastewater on growth, photosynthesis and pollen morphology of spinach plants. Ecotoxicol. Environ. Saf. 2012, 80, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.; Calatayud, A. Non-invasive tools to estimate stress-induced changes in photosynthetic performance in plants inhabiting Mediterranean areas. Environ. Exp. Bot. 2014, 103, 42–52. [Google Scholar] [CrossRef]
- Guo, Y.; Tan, J. Recent advances in the application of chlorophyll a fluorescence from photosystem II. Photochem. Photobiol. 2015, 91, 1–14. [Google Scholar] [CrossRef]
- Moustakas, M.; Malea, P.; Zafeirakoglou, A.; Sperdouli, I. Photochemical changes and oxidative damage in the aquatic macrophyte Cymodocea nodosa exposed to paraquat-induced oxidative stress. Pest. Biochem. Physiol. 2016, 126, 28–34. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Sperdouli, I.; Moustaka, J.; Antonoglou, O.; Adamakis, I.-D.S.; Dendrinou-Samara, C.; Moustakas, M. Leaf age-dependent effects of foliar-sprayed CuZn nanoparticles on photosynthetic efficiency and ROS generation in Arabidopsis thaliana. Materials 2019, 12, 2498. [Google Scholar] [CrossRef] [Green Version]
- Moustaka, J.; Meyling, N.V.; Hauser, T.P. Induction of a compensatory photosynthetic response mechanism in tomato leaves upon short time feeding by the chewing insect Spodoptera exigua. Insects 2021, 12, 562. [Google Scholar] [CrossRef]
- Dobrikova, A.; Apostolova, E.; Hanć, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.D.S.; Moustakas, M. Tolerance mechanisms of the aromatic and medicinal plant Salvia sclarea to excess zinc. Plants 2021, 10, 194. [Google Scholar] [CrossRef]
- Sperdouli, I.; Andreadis, S.S.; Adamakis, I.-D.S.; Moustaka, J.; Koutsogeorgiou, E.I.; Moustakas, M. Reactive oxygen species initiate defence responses of potato photosystem II to sap-sucking insect feeding. Insects 2022, 13, 409. [Google Scholar] [CrossRef]
- Moustaka, J.; Meyling, V.N.; Hauser, T.P. Root-associated entomopathogenic fungi modulate host plant’ s photosystem II photochemistry and its response to herbivorous insects. Molecules 2022, 27, 207. [Google Scholar] [CrossRef] [PubMed]
- Stefanov, M.A.; Rashkov, G.D.; Apostolova, E.L. Assessment of the photosynthetic apparatus functions by chlorophyll fluorescence and P 700 absorbance in C3 and C4 plants under physiological conditions and under salt stress. Int. J. Mol. Sci. 2022, 23, 3768. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Adamakis, I.D.S.; Dobrikova, A.; Apostolova, E.; Hanć, A.; Moustakas, M. Excess zinc supply reduces cadmium uptake and mitigates cadmium toxicity effects on chloroplast structure, oxidative stress, and photosystem II photochemical efficiency in Salvia sclarea plants. Toxics 2022, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, R.P.; Oxborough, K.; Pallett, K.E.; Baker, N.R. Rapid noninvasive screening for perturbations of metabolism and plant growth using chlorophyll fluorescence imaging. Plant Physiol. 2003, 132, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorbe, E.; Calatayud, A. Applications of chlorophyll fluorescence imaging technique in horticultural research: A review. Sci. Hortic. 2012, 138, 24–35. [Google Scholar] [CrossRef]
- Iqbal, M.J.; Goodwin, P.H.; Leonardos, E.D.; Grodzinski, B. Spatial and temporal changes in chlorophyll fluorescence images of Nicotiana benthamiana leaves following inoculation with Pseudomonas syringae pv. tabaci. Plant Pathol. 2012, 61, 1052–1062. [Google Scholar] [CrossRef]
- Bayçu, G.; Moustaka, J.; Gevrek-Kürüm, N.; Moustakas, M. Chlorophyll fluorescence imaging analysis for elucidating the mechanism of photosystem II acclimation to cadmium exposure in the hyperaccumulating plant Noccaea caerulescens. Materials 2018, 11, 2580. [Google Scholar] [CrossRef] [Green Version]
- Moustakas, M.; Hanć, A.; Dobrikova, A.; Sperdouli, I.; Adamakis, I.D.S.; Apostolova, E. Spatial heterogeneity of cadmium effects on Salvia sclarea leaves revealed by chlorophyll fluorescence imaging analysis and laser ablation inductively coupled plasma mass spectrometry. Materials 2019, 12, 2953. [Google Scholar] [CrossRef] [Green Version]
- Moustakas, M.; Bayçu, G.; Gevrek-Kürüm, N.; Moustaka, J.; Csatári, I.; Rognes, S.E. Spatiotemporal heterogeneity of photosystem II function during acclimation to zinc exposure and mineral nutrition changes in the hyperaccumulator Noccaea caerulescens. Environ. Sci. Pollut. Res. 2019, 26, 6613–6624. [Google Scholar] [CrossRef]
- Moustakas, M. The role of metal ions in biology, biochemistry and medicine. Materials 2021, 14, 549. [Google Scholar] [CrossRef]
- Suárez, J.C.; Vanegas, J.I.; Contreras, A.T.; Anzola, J.A.; Urban, M.O.; Beebe, S.E.; Rao, I.M. Chlorophyll fluorescence imaging as a tool for evaluating disease resistance of common bean lines in the western Amazon region of Colombia. Plants 2022, 11, 1371. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Dobrikova, A.; Sperdouli, I.; Hanć, A.; Adamakis, I.-D.S.; Moustaka, J.; Apostolova, E. A hormetic spatiotemporal photosystem II response mechanism of salvia to excess zinc exposure. Int. J. Mol. Sci. 2022, 23, 11232. [Google Scholar] [CrossRef] [PubMed]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Broddrick, J.T.; Ware, M.A.; Jallet, D.; Palsson, B.O.; Peers, G. Integration of physiologically relevant photosynthetic energy flows into whole genome models of light-driven metabolism. Plant J. 2022. ahead of print. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef]
- Havaux, M. Plastoquinone in and beyond photosynthesis. Trends Plant Sci. 2020, 25, 1252–1265. [Google Scholar] [CrossRef]
- Gawroński, P.; Burdiak, P.; Scharff, L.B.; Mielecki, J.; Górecka, M.; Zaborowska, M.; Leister, D.; Waszczak, C.; Karpiński, S. CIA2 and CIA2-LIKE are required for optimal photosynthesis and stress responses in Arabidopsis thaliana. Plant J. 2021, 105, 619–638. [Google Scholar] [CrossRef]
- Wilson, K.E.; Ivanov, A.G.; Öquist, G.; Grodzinski, B.; Sarhan, F.; Huner, N.P.A. Energy balance, organellar redox status, and acclimation to environmental stress. Can. J. Bot. 2006, 84, 1355–1370. [Google Scholar] [CrossRef]
- Lodeyro, A.F.; Krapp, A.R.; Carrillo, N. Photosynthesis and chloroplast redox signaling in the age of global warming: Stress tolerance, acclimation, and developmental plasticity. J. Exp. Bot. 2021, 72, 5919–5937. [Google Scholar]
- Pfannschmidt, T.; Yang, C.H. The hidden function of photosynthesis: A sensing system for environmental conditions that regulates plant acclimation responses. Protoplasma 2012, 249, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Busch, F.A. Opinion: The red-light response of stomatal movement is sensed by the redox state of the photosynthetic electron transport chain. Photosynth. Res. 2014, 119, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Tsabari, O.; Nevo, R.; Meir, S.; Carrillo, L.R.; Kramer, D.M.; Reich, Z. Differential effects of ambient or diminished CO2 and O2 levels on thylakoid membrane structure in light-stressed plants. Plant J. 2015, 81, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.-J.; Turkan, I.; Krieger-Liszkay, A. Redox- and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef] [Green Version]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef]
- Serôdio, J.; Campbell, D.A. Photoinhibition in optically thick samples: Effects of light attenuation on chlorophyll fluorescence-based parameters. J. Theor. Biol. 2021, 513, 110580. [Google Scholar] [CrossRef]
- Raven, J.A. The cost of photoinhibition. Physiol. Plant. 2011, 142, 87–104. [Google Scholar] [CrossRef]
- Campbell, D.A.; Tyystjärvi, E. Parameterization of photosystem II photoinactivation and repair. Biochim. Biophys. Acta 2012, 1817, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Tyystjärvi, E. Photoinhibition of Photosystem II. Int. Rev. Cell Mol. Biol. 2013, 300, 243–303. [Google Scholar]
- Głowacka, K.; Kromdijk, J.; Kucera, K.; Xie, J.; Cavanagh, A.P.; Leonelli, L.; Leakey, A.D.B.; Ort, D.R.; Niyogi, K.K.; Long, S.P. Photosystem II Subunit S overexpression increases the efficiency of water use in a field-grown crop. Nat. Commun. 2018, 9, 868. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvides, A.M.; Velez-Ramirez, A.I.; Fotopoulos, V. Challenging the water stress index concept: Thermographic assessment of Arabidopsis transpiration. Physiol. Plant. 2022, 174, e13762. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Allen, J.F. Lessons from redox signaling in plants. Antioxid. Redox Signal. 2003, 5, 3–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornas, A.; Kuźniak, E.; Slesak, I.; Miszalski, Z. The key role of the redox status in regulation of metabolism in photosynthesizing organisms. Acta Biochim. Pol. 2010, 57, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Grieu, P.; Robin, C.; Guckert, A. Effect of drought on photosynthesis in Trifolium repens: Maintenance of photosystem II efficiency and of measured photosynthesis. Plant Physiol. Biochem. 1995, 33, 19–24. [Google Scholar]
- Tezara, W.; Mitchell, V.J.; Driscoll, S.D.; Lawlor, D.W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 1999, 401, 914–917. [Google Scholar] [CrossRef]
- Ghannoum, O.; Conroy, J.P.; Driscoll, S.P.; Paul, M.J.; Foyer, C.H.; Lawlor, D.W. Nonstomatal limitations are responsible for drought induced photosynthetic inhibition in four C4 grasses. New Phytol. 2003, 159, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Malea, P.; Charitonidou, K.; Sperdouli, I.; Mylona, Z.; Moustakas, M. Zinc uptake, photosynthetic efficiency and oxidative stress in the seagrass Cymodocea nodosa exposed to ZnO nanoparticles. Materials 2019, 12, 2101. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, A.; Barreno, E. Chlorophyll a fluorescence, antioxidant enzymes and lipid peroxidation in tomato in response to ozone and benomyl. Environ. Pollut. 2001, 115, 283–289. [Google Scholar] [CrossRef]
- Terashima, I. Anatomy of non-uniform leaf photosynthesis. Photosyn. Res. 1992, 31, 195212. [Google Scholar] [CrossRef]
- Calatayud, A.; Roca, D.; Martínez, P.F. Spatial-temporal variations in rose leaves under water stress conditions studied by chlorophyll fluorescence imaging. Plant Physiol. Biochem. 2006, 44, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, E.; Horiguchi, G.; Tsukaya, H. Mechanisms of leaf tooth formation in Arabidopsis. Plant J. 2010, 62, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Siebke, K.; Lippert, P.; Baur, B.; Mukherjee, U.; Weis, E. Sink–source transition in tobacco leaves visualized using chlorophyll fluorescence imaging. New Phytol. 2001, 151, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Pulwarty, R.S.; Sivakumar, M.V.K. Information systems in a changing climate: Early warnings and drought risk management. Weather Clim. Extrem. 2014, 3, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Van Ginkel, M.; Biradar, C. Drought early warning in agri-food systems. Climate 2021, 9, 134. [Google Scholar] [CrossRef]
- Moustakas, M.; Guidi, L.; Calatayud, A. Editorial: Chlorophyll fluorescence analysis in biotic and abiotic stress, volume II. Front. Plant Sci. 2022, 13, 1066865. [Google Scholar] [CrossRef]
- Zhao, D.; Eyre, J.X.; Wilkus, E.; de Voil, P.; Broad, I.; Rodriguez, D. 3D characterization of crop water use and the rooting system in field agronomic research. Comput. Electron. Agric. 2022, 202, 107409. [Google Scholar] [CrossRef]
- Antonoglou, O.; Moustaka, J.; Adamakis, I.D.; Sperdouli, I.; Pantazaki, A.; Moustakas, M.; Dendrinou-Samara, C. Nanobrass CuZn nanoparticles as foliar spray non phytotoxic fungicides. ACS Appl. Mater. Interfaces 2018, 10, 4450–4461. [Google Scholar] [CrossRef]
Figure 1.
Light energy capture and utilization in the thylakoids of chloroplasts. The electron transport chain from photosystem II (PSII) to photosystem I (PSI) and finally to ferredoxin (Fdx) to form NADPH, is depicted. Detail explanation in the text (Adopted from [26]).
Figure 1.
Light energy capture and utilization in the thylakoids of chloroplasts. The electron transport chain from photosystem II (PSII) to photosystem I (PSI) and finally to ferredoxin (Fdx) to form NADPH, is depicted. Detail explanation in the text (Adopted from [26]).

Figure 2.
A typical modulated fluorescence trace using dark-adapted leaf material showing how Fo, Fm, Fo′, Fm′ and Fs, are formed to measure chlorophyll fluorescence parameters. In the dark-adapted state a “measuring light” of low light intensity is switched on to induce electron transport through PSII and to elicit the minimal level of chlorophyll fluorescence, termed Fo. A brief saturating pulse of light results in the formation of the maximum yield of fluorescence, Fm. The difference between Fm and Fo is the variable fluorescence, Fm. The application of saturating pulses under actinic light illumination closes all the reaction centers and provides the maximum fluorescence in the light-adapted state, termed Fm′. The steady-state level of fluorescence in the light is termed, Fs and is measured immediately before switching off the actinic light. Fo′ is measured immediately after switching off the actinic light. The difference between Fm′ and Fo′ is the variable fluorescence, Fv′ (Adopted from [26]).
Figure 2.
A typical modulated fluorescence trace using dark-adapted leaf material showing how Fo, Fm, Fo′, Fm′ and Fs, are formed to measure chlorophyll fluorescence parameters. In the dark-adapted state a “measuring light” of low light intensity is switched on to induce electron transport through PSII and to elicit the minimal level of chlorophyll fluorescence, termed Fo. A brief saturating pulse of light results in the formation of the maximum yield of fluorescence, Fm. The difference between Fm and Fo is the variable fluorescence, Fm. The application of saturating pulses under actinic light illumination closes all the reaction centers and provides the maximum fluorescence in the light-adapted state, termed Fm′. The steady-state level of fluorescence in the light is termed, Fs and is measured immediately before switching off the actinic light. Fo′ is measured immediately after switching off the actinic light. The difference between Fm′ and Fo′ is the variable fluorescence, Fv′ (Adopted from [26]).
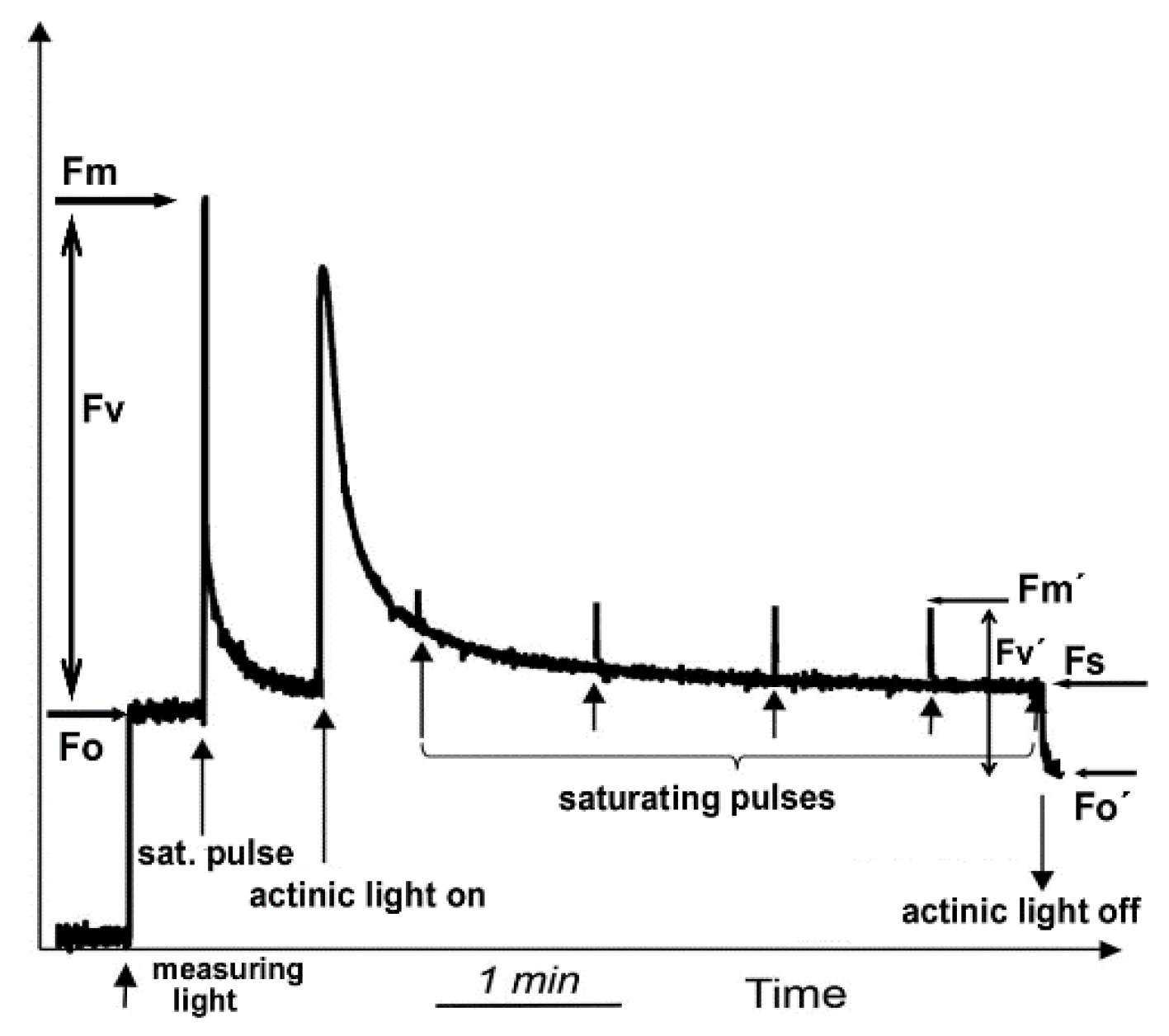
Figure 3.
Color-coded leaf images of the maximum efficiency of Arabidopsis thaliana PSII photochemistry (Fv/Fm). Fv/Fm decreased from the control value of 0.811 (a), with the onset of drought stress (OnDS) to 0.775 (b), and decreased further to 0.714 under mild drought stress (MiDS) (c); while under further water deficit treatment, Fv/Fm recovered to 0.743 at moderate drought stress (MoDS) (d).
Figure 3.
Color-coded leaf images of the maximum efficiency of Arabidopsis thaliana PSII photochemistry (Fv/Fm). Fv/Fm decreased from the control value of 0.811 (a), with the onset of drought stress (OnDS) to 0.775 (b), and decreased further to 0.714 under mild drought stress (MiDS) (c); while under further water deficit treatment, Fv/Fm recovered to 0.743 at moderate drought stress (MoDS) (d).

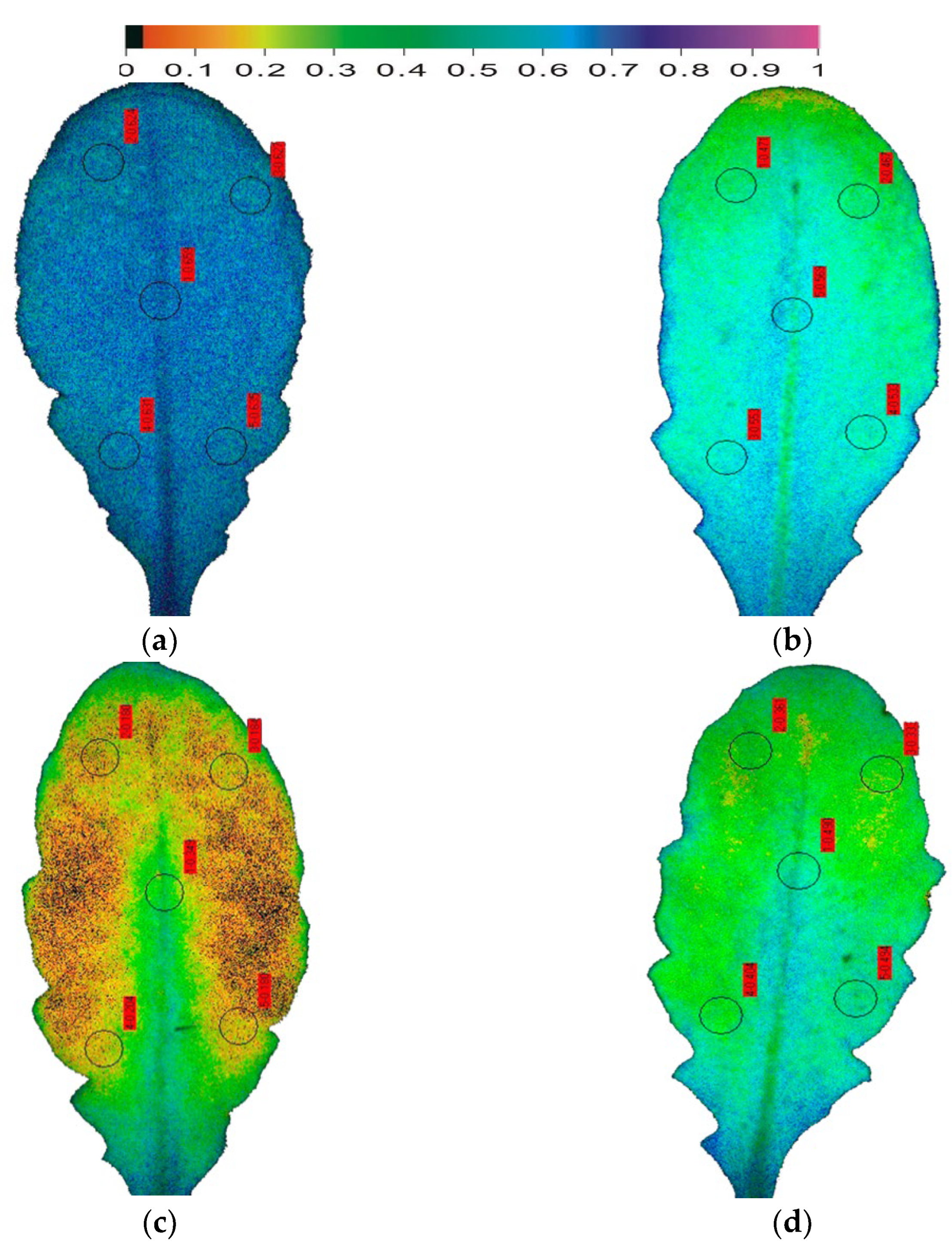
Figure 4.
Color-coded images of the fraction of open PSII reaction centres (qp = [Fm′ − Fs]/[Fm′ − Fo′]), representing the redox state of QA, in Arabidopsis thaliana leaves. The fraction of open PSII reaction centres (qp) decreased from the control value of 0.635 (a), with the onset of drought stress (OnDS, 24 h DS) to 0.519 (b), under mild drought stress (MiDS) decreased further to 0.218 (c); while under moderate drought stress (MoDS), qp recovered to 0.416 (d).
Figure 4.
Color-coded images of the fraction of open PSII reaction centres (qp = [Fm′ − Fs]/[Fm′ − Fo′]), representing the redox state of QA, in Arabidopsis thaliana leaves. The fraction of open PSII reaction centres (qp) decreased from the control value of 0.635 (a), with the onset of drought stress (OnDS, 24 h DS) to 0.519 (b), under mild drought stress (MiDS) decreased further to 0.218 (c); while under moderate drought stress (MoDS), qp recovered to 0.416 (d).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Chlorophyll fluorescence parameters used in studies of photosystem II photochemistry with their definitions.
Table 1.
Chlorophyll fluorescence parameters used in studies of photosystem II photochemistry with their definitions.
| Parameter | Definition | Calculation |
|---|---|---|
| Fv/Fm | Maximum efficiency of PSII photochemistry | (Fm − Fo)/Fm |
| ΦPSII | Effective quantum yield of PSII photochemistry | (Fm′ − Fs)/Fm′ |
| ΦNPQ | Quantum yield of regulated non-photochemical energy loss in PSII | Fs/Fm′ − Fs/Fm |
| ΦNO | Quantum yield of nonregulated energy loss in PSII | Fs/Fm |
| Fv′/Fm′ | Efficiency of open PSII centers | (Fm’ − Fo′)/Fm′ |
| Fv/Fo | Efficiency of the oxygen evolving complex (OEC) on the donor side of PSII | (Fm − Fo)/Fo |
| ETR | Electron transport rate | ΦPSII × PAR × c × abs, where PAR is the photosynthetically active radiation, c is 0.5, and abs is the total light absorption of the leaf taken as 0.84 |
| qp | Photochemical quenching, representing the fraction of PSII reaction centers in open state (puddle model) | (Fm′ − Fs)/(Fm′ − Fo′) |
| NPQ | Non-photochemical quenching reflecting the dissipation of excitation energy as heat | (Fm − Fm′)/Fm′ |
| EXC | Excess excitation energy | (Fv/Fm − ΦPSII)/Fv/Fm |
| 1−qL | The fraction of PSII reaction centers in closed state (based on a “lake” model for the photosynthetic unit) | qp × Fo′/Fs |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moustakas, M.; Sperdouli, I.; Moustaka, J. Early Drought Stress Warning in Plants: Color Pictures of Photosystem II Photochemistry. Climate 2022, 10, 179. https://doi.org/10.3390/cli10110179
AMA Style
Moustakas M, Sperdouli I, Moustaka J. Early Drought Stress Warning in Plants: Color Pictures of Photosystem II Photochemistry. Climate. 2022; 10(11):179. https://doi.org/10.3390/cli10110179
Chicago/Turabian StyleMoustakas, Michael, Ilektra Sperdouli, and Julietta Moustaka. 2022. "Early Drought Stress Warning in Plants: Color Pictures of Photosystem II Photochemistry" Climate 10, no. 11: 179. https://doi.org/10.3390/cli10110179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.