
DNA Methylation Profiles Are Stable in H3 K27M-Mutant Diffuse Midline Glioma Neurosphere Cell Lines
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview
2.2. Histology Evaluation and Tumor Specimen Classification
2.3. In Vitro Cell Culture
2.4. DNA Extraction from Brain Tumor Neurospheres
2.5. DNA Extraction of Patient Tumor from FFPE Tissue Blocks
2.6. Genome-Wide Methylation Profiling
2.7. CNV and tSNE
3. Results
3.1. Classifier Scores
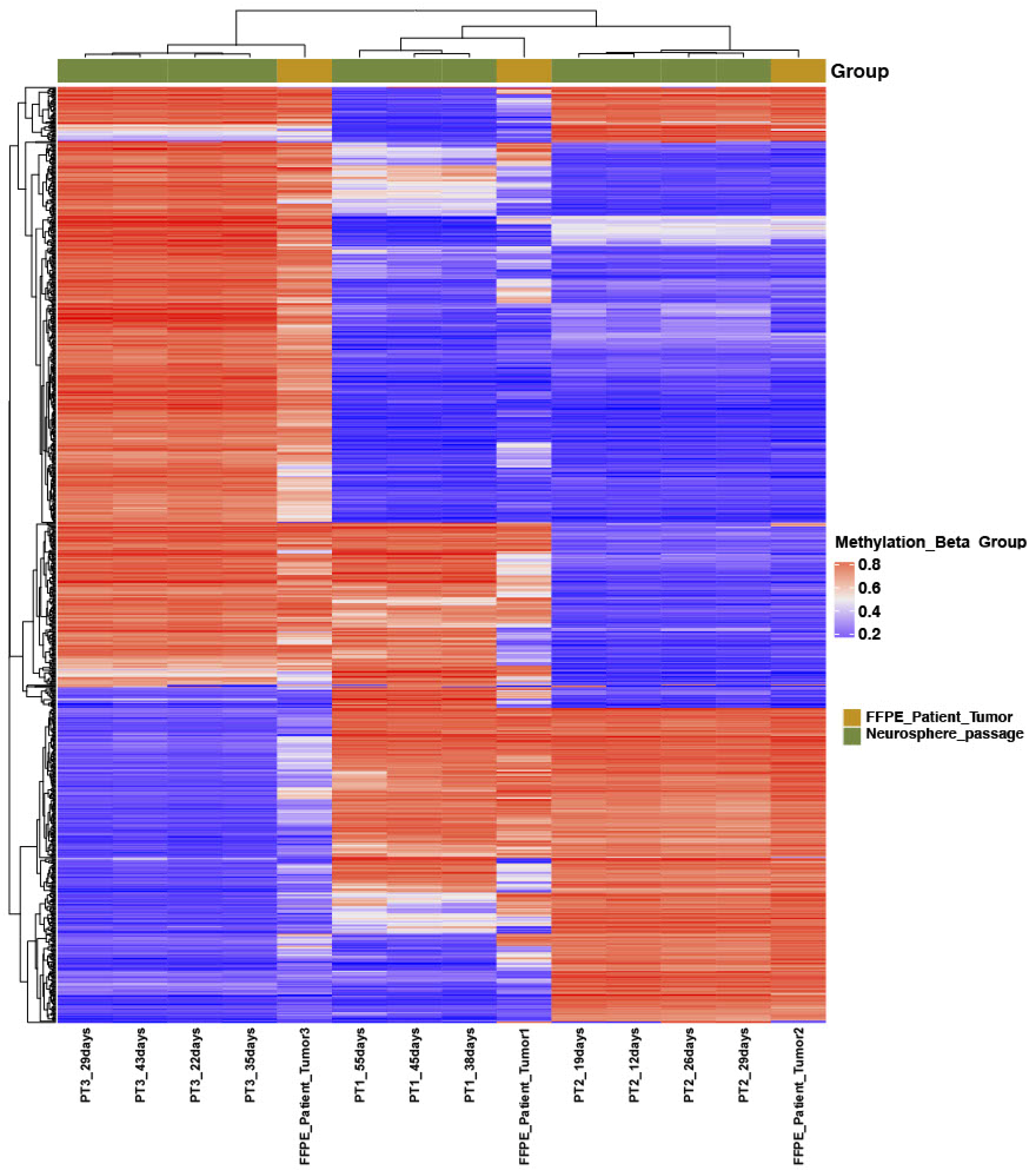
3.2. Clustering Analysis
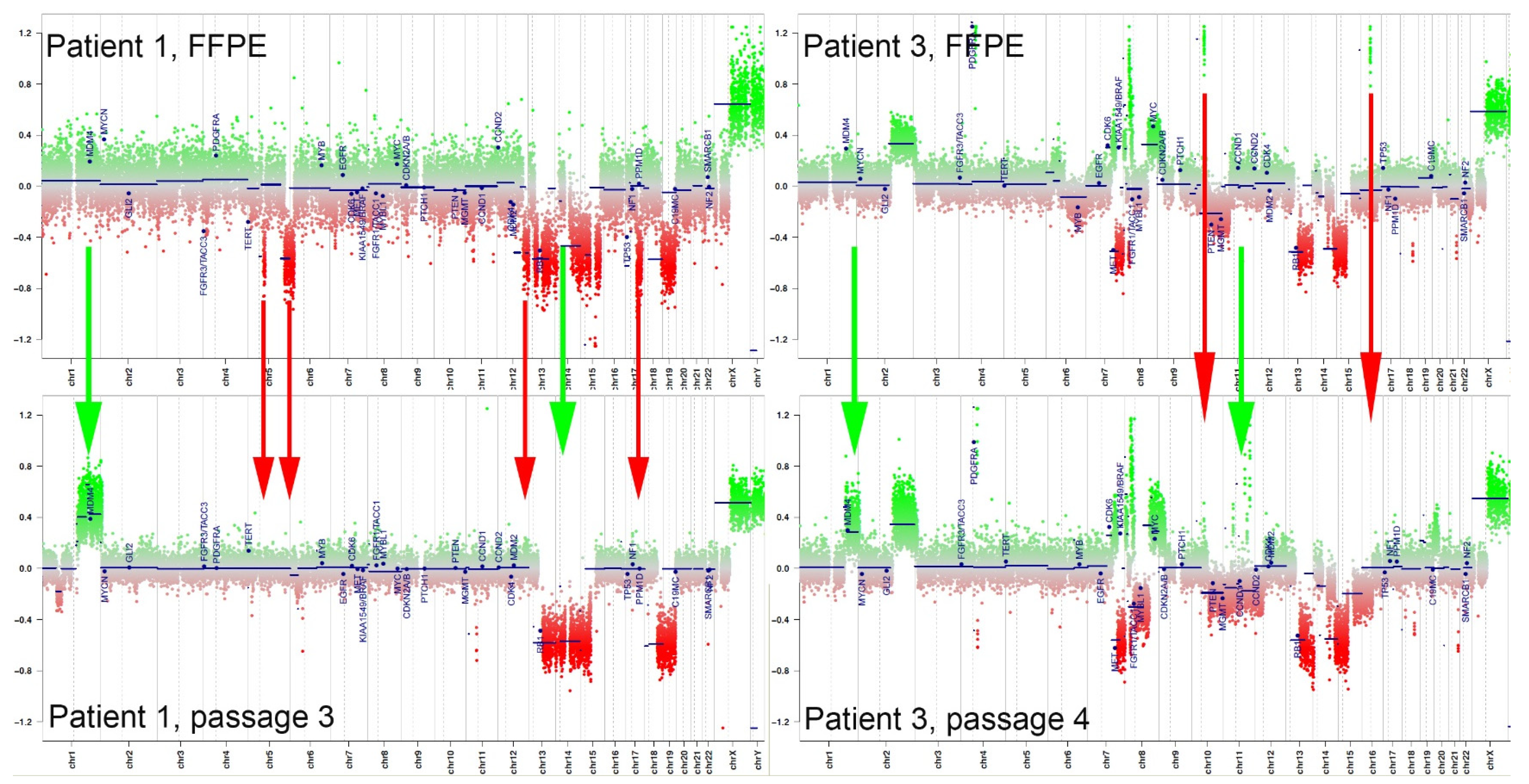
3.3. Gain of Chromosome 1q
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veldhuijzen van Zanten, S.E.M.; Lane, A.; Heymans, M.W.; Baugh, J.; Chaney, B.; Hoffman, L.M.; Doughman, R.; Jansen, M.H.A.; Sanchez, E.; Vandertop, W.P.; et al. External validation of the diffuse intrinsic pontine glioma survival prediction model: A collaborative report from the International DIPG Registry and the SIOPE DIPG Registry. J. Neurooncol. 2017, 134, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.L.; Monje, M. A Protocol for Rapid Post-mortem Cell Culture of Diffuse Intrinsic Pontine Glioma (DIPG). J. Vis. Exp. 2017, 121, e55360. [Google Scholar] [CrossRef]
- Capes-Davis, A.; Theodosopoulos, G.; Atkin, I.; Drexler, H.G.; Kohara, A.; MacLeod, R.A.; Masters, J.R.; Nakamura, Y.; Reid, Y.A.; Reddel, R.R.; et al. Check your cultures! A list of cross-contaminated or misidentified cell lines. Int. J. Cancer 2010, 127, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pandita, A.; Aldape, K.; Zadeh, G.; Guha, A.; James, C.D. Contrasting In Vivo and In Vitro Fates of Glioblastoma Cell Subpopulations with Amplified EGFR. Genes Chromosomes Cancer 2003, 39, 29–36. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.A.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.; Burns, R.T.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Rey, V.; Menéndez, S.T.; Estupiñán, Ó.; Rodríguez, A.; Santos, L.; Tornín, J.; Martinez-Cruzado, L.; Castillo, D.; Ordóñez, G.R.; Costilla, S.; et al. New Chondrosarcoma Cell Lines with Preserved Stem Cell Properties to Study the Genomic Drift During In Vitro/In Vivo Growth. J. Clin. Med. 2019, 8, 455. [Google Scholar] [CrossRef] [PubMed]
- Malagutti-Ferreira, M.J.; Crispim, B.D.A.; Barufatti, A.; Cardoso, S.S.; Guarnier, L.P.; Rodriguez, F.; Soares, M.R.; Da Silva Antunes, R.F.; Ribeiro-Paes, J.T. Genomic Instability in Long-Term Culture of Human Adipose-Derived Mesenchymal Stromal Cells. Braz. J. Med. Biol. Res. 2023, 56, e12713. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Bergström, T.; Jiang, Y.; Johansson, P.; Marinescu, V.D.; Lindberg, N.; Segerman, A.; Wicher, G.; Niklasson, M.; Baskaran, S.; et al. The Human Glioblastoma Cell Culture Resource: Validated Cell Models Representing All Molecular Subtypes. EBioMedicine 2015, 2, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Capper, D.; Jones, D.; Sill, M.; Hovestadt, V.; Schrimpf, D.; Sturm, D.; Koelsche, C.; Sahm, F.; Chávez, L.; Reuß, D.; et al. DNA Methylation-Based Classification of Central Nervous System Tumours. Nature 2018, 555, 469–474. [Google Scholar] [CrossRef]
- Sturm, D.; Orr, B.A.; Toprak, U.; Hovestadt, V.; Jones, D.; Capper, D.; Sill, M.; Buchhalter, I.; Northcott, P.A.; Leis, I.; et al. New Brain Tumor Entities Emerge from Molecular Classification of CNS-PNETs. Cell 2016, 164, 1060–1072. [Google Scholar] [CrossRef]
- Pajtler, K.W.; Witt, H.; Sill, M.; Jones, D.; Hovestadt, V.; Kratochwil, F.; Wani, K.; Tatevossian, R.; Punchihewa, C.; Johann, P.; et al. Molecular Classification of Ependymal Tumors across All CNS Compartments, Histopathological Grades, and Age Groups. Cancer Cell 2015, 27, 728–743. [Google Scholar] [CrossRef]
- Lim, U.; Song, M.-A. DNA Methylation as a Biomarker of Aging in Epidemiologic Studies. Methods Mol. Biol. 2018, 1856, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Malan-Müller, S.; Seedat, S.; Hemmings, S.M.J. Understanding Posttraumatic Stress Disorder: Insights from the Methylome. Genes Brain Behav. 2013, 13, 52–68. [Google Scholar] [CrossRef]
- Weng, J.-H.; Wu, L.S.-H.; Lee, C.; Hsu, P.; Cheng, A.T.A. Integrative Epigenetic Profiling Analysis Identifies DNA Methylation Changes Associated with Chronic Alcohol Consumption. Comput. Biol. Med. 2015, 64, 299–306. [Google Scholar] [CrossRef]
- Castel, D.; Philippe, C.; Kergrohen, T.; Sill, M.; Merlevede, J.; Barret, E.; Puget, S.; Sainte-Rose, C.; Kramm, C.M.; Jones, C.; et al. Transcriptomic and Epigenetic Profiling of ‘Diffuse Midline Gliomas, H3 K27M-Mutant’ Discriminate Two Subgroups Based on the Type of Histone H3 Mutated and Not Supratentorial or Infratentorial Location. Acta Neuropathol. Commun. 2018, 6, 117. [Google Scholar] [CrossRef] [PubMed]
- WHO Classification of Tumours Editorial Board. WHO Classification of Tumours of the Central Nervous System, 4th ed.; Revised; International Agency for Research on Cancer: Lyon, France, 2016; Volume 6. [Google Scholar]
- Varlet, P.; Baker, S.J.; Ellison, D.W.; Jabado, N.; Jones, C.; Jones, D.T.W.; Leske, H.; Orr, B.A.; Solomon, D.A.; Suvà, M.L.; et al. Diffuse Midline Glioma, H3 H27-altered. In WHO Classification of Tumours of the Central Nervous System, 5th ed.; Editorial Board; International Agency for Research on Cancer: Lyon, France, 2021; Volume 6. [Google Scholar]
- Aryee, M.J.; Jaffe, A.E.; Bravo, H.C.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A Flexible and Comprehensive Bioconductor Package for the Analysis of Infinium DNA Methylation Microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Maaten, L.V.D.; Hinton, G.E. Visualizing Data using t-SNE. J. Mach. Learn. Res. 2008, 9, 2579–2605. [Google Scholar]
- Feber, A.; Guilhamon, P.; Lechner, M.; Fenton, T.R.; Wilson, G.A.; Thirlwell, C.; Morris, T.; Flanagan, A.M.; Teschendorff, A.E.; Kelly, J.D.; et al. Using High-Density DNA Methylation Arrays to Profile Copy Number Alterations. Genome Biol. 2014, 15, R30. [Google Scholar] [CrossRef]
- Morán, S.; Vizoso, M.; Martínez-Cardús, A.; Gómez, A.; Matías-Guiu, X.; Chiavenna, S.; Fernandez, A.G.; Esteller, M. Validation of DNA methylation profiling in formalin-fixed paraffin-embedded samples using the Infinium HumanMethylation450 Microarray. Epigenetics 2014, 9, 829–833. [Google Scholar] [CrossRef]
- Espinal, A.C.; Wang, D.; Yan, L.; Liu, S.; Tang, L.; Hu, Q.; Morrison, C.; Ambrosone, C.B.; Higgins, M.J.; Sucheston-Campbell, L.E. A methodological study of genome-wide DNA methylation analyses using matched archival formalin-fixed paraffin embedded and fresh frozen breast tumors. Oncotarget 2017, 8, 14821–14829. [Google Scholar] [CrossRef] [PubMed]
- Warren, K.E.; Killian, K.; Suuriniemi, M.; Wang, Y.; Quezado, M.; Meltzer, P.S. Genomic Aberrations in Pediatric Diffuse Intrinsic Pontine Gliomas. Neuro-Oncology 2011, 14, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Zarghooni, M.; Bartels, U.; Lee, E.; Buczkowicz, P.; Morrison, A.; Huang, A.; Bouffet, É.; Hawkins, C. Whole-Genome Profiling of Pediatric Diffuse Intrinsic Pontine Gliomas Highlights Platelet-Derived Growth Factor Receptor α and Poly (ADP-Ribose) Polymerase As Potential Therapeutic Targets. J. Clin. Oncol. 2010, 28, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Barrow, J.H.; Adamowicz-Brice, M.; Cartmill, M.; Macarthur, D.; Lowe, J.; Robson, K.; Bründler, M.-A.; Walker, D.; Coyle, B.; Grundy, R.G. Homozygous Loss of ADAM3A Revealed by Genome-Wide Analysis of Pediatric High-Grade Glioma and Diffuse Intrinsic Pontine Gliomas. Neuro-Oncology 2010, 13, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Castel, D.; Philippe, C.; Calmon, R.; Dret, L.L.; Truffaux, N.; Boddaert, N.; Pagès, M.; Taylor, K.R.; Saulnier, P.; Lacroix, L.; et al. Histone H3F3A and HIST1H3B K27M Mutations Define Two Subgroups of Diffuse Intrinsic Pontine Gliomas with Different Prognosis and Phenotypes. Acta Neuropathol. 2015, 130, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Segers, H.; Van Den Heuvel-Eibrink, M.M.; Williams, R.D.; Van Tinteren, H.; Vujanić, G.; Pieters, R.; Pritchard-Jones, K.; Bown, N. Gain of 1q Is a Marker of Poor Prognosis in Wilms’ Tumors. Genes Chromosomes Cancer 2013, 52, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.M.; Barwick, B.G.; Joseph, N.; Heffner, L.T.; Hofmeister, C.C.; Bernal, L.; Dhodapkar, M.V.; Gupta, V.A.; Jaye, D.L.; Wu, J.; et al. Gain of Chromosome 1q Is Associated with Early Progression in Multiple Myeloma Patients Treated with Lenalidomide, Bortezomib, and Dexamethasone. Blood Cancer J. 2019, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Kjellman, P.; Lagercrantz, S.; Höög, A.; Wallin, G.; Larsson, C.; Zedenius, J. Gain of 1q and Loss of 9q21.3-q32 Are Associated with a Less Favorable Prognosis in Papillary Thyroid Carcinoma. Genes Chromosomes Cancer 2001, 32, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Dyer, S.; Prebble, E.; Davison, V.; Davies, P.; Ramani, P.; Ellison, D.W.; Grundy, R.G. Genomic Imbalances in Pediatric Intracranial Ependymomas Define Clinically Relevant Groups. Am. J. Pathol. 2002, 161, 2133–2141. [Google Scholar] [CrossRef]
- Gabeau-Lacet, D.; Engler, D.; Gupta, S.; Scangas, G.A.; Betensky, R.A.; Barker, F.G.; Loeffler, J.S.; Louis, D.N.; Mohapatra, G. Genomic Profiling of Atypical Meningiomas Associates Gain of 1Q with Poor Clinical Outcome. J. Neuropathol. Exp. Neurol. 2009, 68, 1155–1165. [Google Scholar] [CrossRef]
- Chiang, J.; Dalton, J.; Upadhyaya, S.A.; Patay, Z.; Qaddoumi, I.; Li, X.; Segura, A.D.; Sharma, S.; Ismail, A.; Shurtleff, S.; et al. Chromosome Arm 1q Gain Is an Adverse Prognostic Factor in Localized and Diffuse Leptomeningeal Glioneuronal Tumors with BRAF Gene Fusion and 1p Deletion. Acta Neuropathol. 2018, 137, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Miwa, T.; Hirose, Y.; Sasaki, H.; Ezaki, T.; Yoshida, K.; Kawase, T. Single-Copy Gain of Chromosome 1Q Is a Negative Prognostic Marker in Pediatric Nonependymal, Nonpilocytic Gliomas. Neurosurgery 2011, 68, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Ohka, F.; Okuno, Y.; Aoki, K.; Motomura, K.; Takeuchi, K.; Kusakari, H.; Yanagisawa, N.; Sato, S.; Yamaguchi, J.; et al. H3F3A Mutant Allele Specific Imbalance in an Aggressive Subtype of Diffuse Midline Glioma, H3 K27M-Mutant. Acta Neuropathol. Commun. 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age */Sex | Sample No. | Sample ID | Classification | Score |
|---|---|---|---|---|
| 5F | 1 | IFF BT105 FFPE patient tumor | DMG H3 K27M | 0.9962 |
| 2 | IFF BT105 F1p1d38 (passage 1) | DMG H3 K27M | 0.9931 | |
| 3 | IFF BT105 F1p2d45 (passage 2) | DMG H3 K27M | 0.9963 | |
| 4 | IFF BT105 F1p3d55 (passage 3) | DMG H3 K27M | 0.9963 | |
| 2M | 5 | IFF BT128 FFPE patient tumor | DMG H3 K27M | 0.9817 |
| 6 | IFF BT128 F1p1d12 (passage 1) | DMG H3 K27M | 0.9973 | |
| 7 | IFF BT128 F1p2d19 (passage 2) | DMG H3 K27M | 0.9967 | |
| 8 | IFF BT128 F1p3d26 (passage 3) | DMG H3 K27M | 0.9959 | |
| 9 | IFF BT128 F1p4d29 (passage 4) | DMG H3 K27M | 0.9958 | |
| 8F | 10 | IFF BT138 FFPE patient tumor | DMG H3 K27M | 0.9999 |
| 11 | IFF BT138 F1p1d22 (passage 1) | DMG H3 K27M | 0.9998 | |
| 12 | IFF BT138 F1p2d29 (passage 2) | DMG H3 K27M | 0.9998 | |
| 13 | IFF BT138 F1p3d35 (passage 3) | DMG H3 K27M | 0.9997 | |
| 14 | IFF BT138 F1p4d43 (passage 4) | DMG H3 K27M | 0.9994 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schniederjan, M.J.; Potnis, C.; Vasudevaraja, V.; Moser, C.D.; Watson, B.; Snuderl, M.; MacDonald, T.; Rogers, B.B. DNA Methylation Profiles Are Stable in H3 K27M-Mutant Diffuse Midline Glioma Neurosphere Cell Lines. Children 2024, 11, 492. https://doi.org/10.3390/children11040492
Schniederjan MJ, Potnis C, Vasudevaraja V, Moser CD, Watson B, Snuderl M, MacDonald T, Rogers BB. DNA Methylation Profiles Are Stable in H3 K27M-Mutant Diffuse Midline Glioma Neurosphere Cell Lines. Children. 2024; 11(4):492. https://doi.org/10.3390/children11040492
Chicago/Turabian StyleSchniederjan, Matthew J., Cahil Potnis, Varshini Vasudevaraja, Catherine D. Moser, Bethany Watson, Matija Snuderl, Tobey MacDonald, and Beverly B. Rogers. 2024. "DNA Methylation Profiles Are Stable in H3 K27M-Mutant Diffuse Midline Glioma Neurosphere Cell Lines" Children 11, no. 4: 492. https://doi.org/10.3390/children11040492